Compositional Analyses Reveal Relationships among Components of Blue Maize Grains


Abstract
:1. Introduction
2. Results
2.1. Descriptive Statistics and Trait Variation
2.2. Performance Statistics of Calibration and Validation Sets
2.3. Multivariate Analysis
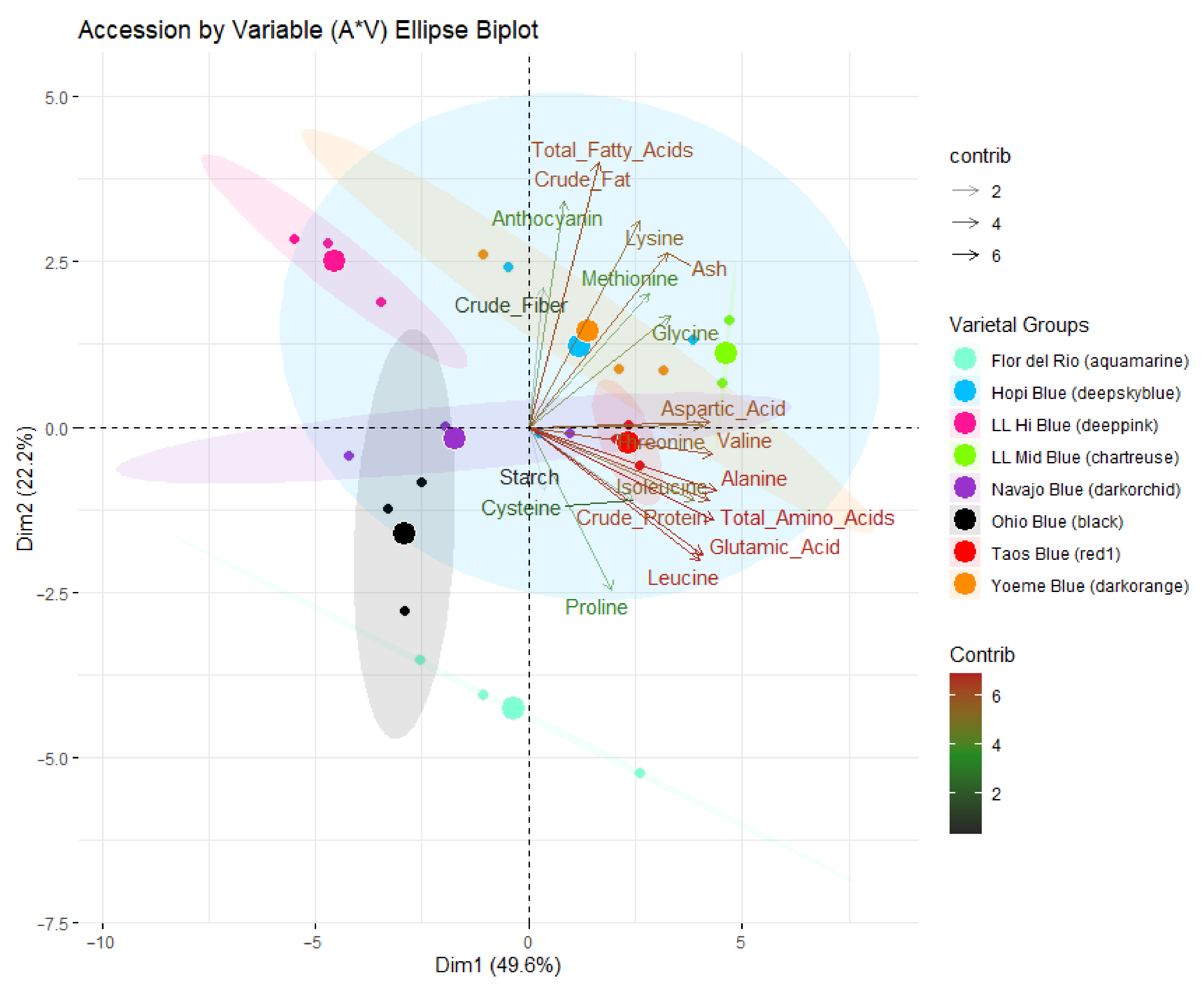
2.3.1. Principal Component Analysis
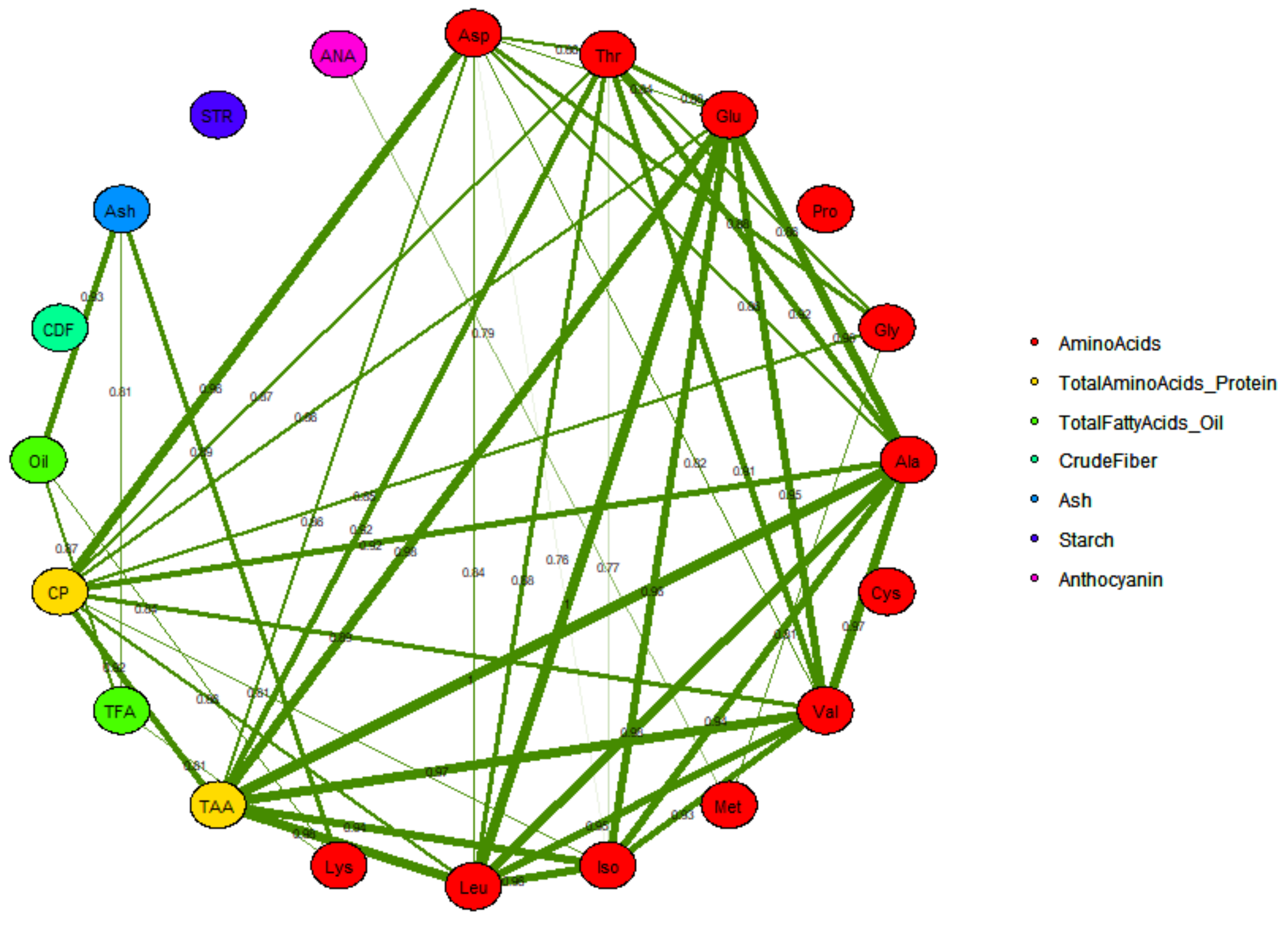
2.3.2. Correlation between Different Grain Compositional Traits and Correlation Network
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Plant Material
4.1.1. Experimental Design
4.1.2. Plant Material
4.1.3. Sample Preparation
4.2. NIR Spectral Data Collection and Pretreatments
4.3. Wet Chemistry for Grain Constituent Analysis
4.4. Data Preprocessing and Construction of Calibration Equation
4.4.1. Data Preprocessing
4.4.2. Construction of Calibration Equations
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Norris, K.H. Instrumentation of infrared radiation. Trans. ASAE 1962, 5, 17–20. [Google Scholar] [CrossRef]
- Hart, J.R.; Norris, K.H.; Golumbic, C. Determination of the moisture content of seeds by near-infrared. Spectrophotometry of their methanol extracts. Cereal. Chem. 1962, 39, 94–99. [Google Scholar]
- Shenk, J.S.; Workman, J.J.; Westerhaus, M.O. Application of NIR spectroscopy to agricultural products. Pract. Spectrosc. Ser. 2001, 27, 419–474. [Google Scholar]
- Cozzolino, D.; Vadell, A.; Ballesteros, F.; Gallietta, G.; Barlocco, N. Combining visible and near-infrared. Spectroscopy with chemometrics to trace muscles from an autochthonous breed of pig produced in.Uruaguay: A feasibility study. Anal. Bioanal. Chem. 2006, 385, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Roggo, Y.; Chalus, P.; Maurer, L.; Lema-Martinez, C.; Edmond, A.; Jent, N. A review of near infrared spectroscopy and chemometrics in pharmaceutical technologies. J. Pharm. Biomed. Analy. 2007, 44, 683–700. [Google Scholar] [CrossRef]
- Orman, B.A.; Schumann, R.A., Jr. Comparison of near-infrared spectroscopy calibration methods for the prediction of protein, oil, and starch in maize grain. J. Agric. Food Chem. 1991, 39, 883–886. [Google Scholar] [CrossRef]
- Osborne, B.G. Applications of near infrared spectroscopy in quality screening of early-generation material in cereal breeding programmes. J. Near Infrared Spec. 2006, 14, 93–101. [Google Scholar] [CrossRef]
- Egesel, C.Ö.; Kahriman, F. Determination of quality parameters in maize grain by NIR reflectance spectroscopy. J. Agric. Sci. 2012, 18, 31–42. [Google Scholar]
- Delwiche, S.R.; Pordesimo, L.O.; Scaboo, A.M.; Pantalone, V.R. Measurement of inorganic phosphorous in soybeans with near-infrared spectroscopy. J. Agric. Food Chem. 2006, 54, 6951–6956. [Google Scholar] [CrossRef]
- Cozzollino, D.; Cynkar, W.U.; Dambergs, R.G.; Janik, L.; Gishen, M. Effect of both homogenization and storage on the spectra of red grapes and on the measurement of total anthocyanins, total soluble solids and pH by visual near infrared spectroscopy. J. Near Infrared Spec. 2005, 13, 213–223. [Google Scholar] [CrossRef]
- Montes, J.M.; Melchinger, A.E.; Reif, J.C. Novel throughput phenotyping platforms in plant genetic studies. Trends Plant Sci. 2007, 12, 433–436. [Google Scholar] [CrossRef]
- Williams, P.J. Near Infrared (NIR) Hyperspectral Imaging for Evaluation of Whole Maize Kernels: Chemometrics for Exploration and Classification. Ph.D. Thesis, Stellenbosch University, Western Cape, South Africa, 2009. [Google Scholar]
- Meng, Q.; Murray, S.C.; Mahan, A.; Collison, A.; Yang, L.; Awika, J. Rapid estimation of phenolic content in colored maize by near-infrared reflectance spectroscopy and its use in breeding. Crop Sci. 2015, 55, 2234–2243. [Google Scholar] [CrossRef]
- Berardo, N.; Mazzinelli, G.; Valoti, P.; Lagana, P.; Redaelli, R. Characterization of maize germplasm for the chemical composition of the grain. J. Agric. Food Chem. 2009, 57, 2378–2384. [Google Scholar] [CrossRef]
- Williams, P.J. Near-infrared technology—Getting the best out of light. In A Short Course in the Practical Implementation of Near-Infrared Spectroscopy for the User; PDK Projects, Inc.: Nanaimo, CA, USA, 2007. [Google Scholar]
- Maslovaric, M.; de la Roza-Delgado, B.; Jankovic, S.; Levic, J.; Jovanovic, R. Development of NIR calibration models to predict chemical composition and in vitro organic matter digestibility of maize silage. In Proceedings of the 10th International Symposium Modern Trends in Livestock Production, Belgrade, Serbia, 2–4 October 2013. [Google Scholar]
- Hurburgh, C.R., Jr. Moisture and composition analysis in the maize and soybean market. Cereal Foods World 1988, 33, 503–505. [Google Scholar]
- Siska, J.; Hurburgh, C.R., Jr. Maize density measurement by near-infrared transmittance. Trans. ASAE 1995, 38, 1821–1824. [Google Scholar] [CrossRef]
- Velasco, L.; Goffman, F.D.; Becker, H.C. Development of calibration equations to predict oil content and fatty acid composition in Brassicaceae germplasm by near-infrared reflectance spectroscopy. J. Am. Oil Chem. Soc. 1999, 76, 25–30. [Google Scholar] [CrossRef]
- Baye, T.M.; Pearson, T.C.; Settles, M.A. Development of a calibration to predict maize seed composition. using single kernel near infrared spectroscopy. J. Cereal Sci. 2006, 43, 236–243. [Google Scholar] [CrossRef]
- Spielbauer, G.; Armstrong, P.; Baier, J.W.; Allen, W.B.; Richardson, K.; Shen, B.; Settles, A.M. High-throughput near-infrared reflectance spectroscopy for predicting quantitative and qualitative composition phenotypes of individual maize kernels. Cereal Chem. 2009, 86, 556–564. [Google Scholar] [CrossRef]
- Fox, G.P.; Osborne, B.; Bowman, J.; Kelly, A.; Cakir, M.; Poulsen, D.; Inkerman, A.; Henry, R. Measurement of genetic and environmental variation in barley (Hordeum vulgare) grain hardness. J. Cereal Sci. 2007, 46, 82–92. [Google Scholar] [CrossRef]
- Flint-Garcia, S.A.; Bodnar, A.L.; Scott, M.P. Wide variability in kernel composition, seed characteristics, and zein profiles among diverse maize inbreds, landraces, and teosinte. Theor. Appl. Genet. 2009, 119, 1129–1142. [Google Scholar] [CrossRef] [Green Version]
- Yadav, O.P.; Hossain, F.; Karjagi, C.G.; Kumar, B.; Zaidi, P.H.; Jat, S.L.; Chawla, J.S.; Kaul, J.; Hooda, K.S.; Kumar, P.; et al. Genetic improvement of maize in India: Retrospect and prospects. Agric. Res. 2015, 4, 325–338. [Google Scholar] [CrossRef]
- Meng, Y.; Li, J.; Liu, J.; Hu, H.; Li, W.; Liu, W.; Chen, S. Ploidy effect and genetic architecture exploration of stalk traits using DH and its corresponding haploid populations in maize. BMC Plant Biol. 2016, 16, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.R.; Sykes, J.; Glover, D.V. Classification of single- and double-mutant maize endosperm genotypes by near-infrared transmittance spectroscopy. Cereal. Chem. 2000, 77, 774–778. [Google Scholar] [CrossRef]
- Berardo, N.; Brenna, O.V.; Amato, A.; Valotia, P.; Pisacanea, V.; Mottoa, M. Carotenoids concentration among maize genotypes measured by near infrared reflectance spectroscopy (NIRS). Innov. Food Sci. Emerg. Technol. 2004, 5, 393–398. [Google Scholar] [CrossRef]
- Nankar, A.; Holguin, F.O.; Scott, M.P.; Pratt, R.C. Grain and nutritional quality traits of Southwestern U.S. blue maize landraces. Cereal Chem. 2017, 94, 950–955. [Google Scholar] [CrossRef]
- Welle, R.; Greten, W.; Müler, T.; Weber, G.; Wehrmann, H. Application of near infrared spectroscopy on-combine in maize grain breeding. J. Near Infrared Spec. 2005, 13, 69–75. [Google Scholar] [CrossRef]
- Robutti, J.L.; Borras, F.; Ferrer, M.; Percibaldi, M.; Knuston, C.A. Evaluation of quality factors in Argentine maize races. Cereal Chem. 2000, 77, 24–26. [Google Scholar] [CrossRef]
- Pearson, T.C.; Wicklow, D.T.; Maghirang, F.; Xie, F.; Dowell, F.E. Detecting aflatoxin in single maize kernels by transmit and reflectance spectroscopy. Trans. ASAE 2001, 44, 1247–1254. [Google Scholar] [CrossRef] [Green Version]
- Dowell, F.E.; Pearson, T.C.; Maghirang, E.B.; Xie, F.; Wicklow, D.T. Reflectance and transmittance spectroscopy applied to detecting fumonisin in single maize kernels infected with Fusarium verticillioides. Cereal Chem. 2002, 79, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Doehlert, D.C.; Lambert, R.J. Metabolic characteristics associated with starch, protein, and oil deposition in developing maize kernels. Crop Sci. 1991, 31, 151–157. [Google Scholar] [CrossRef]
- Scott, M.P.; Edwards, J.W.; Bell, C.P.; Schussler, J.R.; Smith, J.S. Grain composition and amino acid content in maize cultivars representing 80 years of commercial maize varieties. Maydica 2006, 51, 417–423. [Google Scholar]
- Gutierrez-Rojas, A.; Scott, M.P.; Leyva, O.R.; Menz, M.; Betran, J. Phenotypic characterization of quality protein maize endosperm modification and amino acid contents in a segregating recombinant inbred population. Crop Sci. 2008, 48, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Nankar, A.N.; Grant, L.; Scott, M.P.; Pratt, R.C. Agronomic and kernel compositional traits of blue maize landraces from the southwestern United States. Crop Sci. 2016, 56, 2663–2674. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.P.; Bhatnagar, S.; Betran, J. Tryptophan and methionine levels in quality protein maize breeding germplasm. Maydica 2004, 49, 303–311. [Google Scholar]
- Reynolds, T.L.; Nemeth, M.A.; Glenn, K.C.; Ridley, W.P.; Astwood, J.D. Natural variability of metabolites in maize grain: Differences due to genetic background. J. Agric. Food Chem. 2005, 53, 10061–10067. [Google Scholar] [CrossRef]
- Jiang, H.Y.; Zhu, Y.J.; Wei, L.M.; Dai, J.R.; Song, T.M.; Yan, Y.L.; Chen, S.J. Analysis of protein, starch, and oil of single intact kernels by near infrared reflectance spectroscopy (NIRS) in maize (Zea mays L.). Plant Breed. 2007, 126, 492–497. [Google Scholar] [CrossRef]
- Wehling, R.L.; Jackson, D.S.; Hooper, D.G.; Ghaedian, A.R. Prediction of wet-milling starch yield from maize by Near-Infrared Spectroscopy. Cereal Chem. 1996, 70, 720–723. [Google Scholar]
- Lu, W.Z.; Yuan, H.F.; Xu, G.T. Modern Analysis Technique of NIR; Sinca Petro-chemistry Press: Beijing, China, 2000; pp. 165–192. [Google Scholar]
- Ngonyamo-Majee, D.; Shaver, R.D.; Coors, J.G.; Sapienza, D.; Correa, C.E.S.; Lauer, J.G.; Berzaghi, P. Relationship between kernel vitreousness and dry matter degradability for diverse maize germplasm I. Development of near-infrared reflectance spectroscopy calibrations. Anim. Feed Sci. 2008, 142, 247–258. [Google Scholar] [CrossRef]
- Cogdill, R.P.; Hurburgh, C.R.; Rippke, G.R. Single-kernel maize analysis by near-infrared hyperspectral imaging. Trans. ASAE 2004, 47, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Lošák, T.; Hlušek, J.; Filipčík, R.; Pospíšilová, L.; Maňásek, J.; Prokeš, K.; Buňka, F.; Kráčmar, S.; Martensson, A.; Orosz, F. Effect of nitrogen fertilization on metabolisms of essential and non-essential amino acids in field-grown grain maize (Zea mays L.). Plant Soil Environ. 2010, 56, 574–579. [Google Scholar]
- Cone, K.C. Anthocyanin synthesis in maize aleurone tissue. In Endosperm; Olsen, O.A., Ed.; Springer: Berlin, Germany, 2007; Volume 8. [Google Scholar] [CrossRef]
- Jetter, R.; Kunst, L. Plant surface lipid biosynthetic pathways and their utility for metabolic engineering of waxes and hydrocarbon biofuels. Plant J. 2008, 54, 670–683. [Google Scholar] [CrossRef] [PubMed]
- Lane, H.M.; Murray, S.C.; Montesinos-López, O.A.; Montesinos-López, A.; Crossa, J.; Rooney, D.K.; Barrero-Farfan, I.D.; De La Fuente, G.N.; Morgan, C.L.S. Phenomic selection and prediction of maize grain yield from near-infrared reflectance spectroscopy of kernels. TPPJ. 2020, 3, e20002. [Google Scholar] [CrossRef] [Green Version]
- Marita, J.M.; Vermerris, W.; Ralph, J.; Hatfield, R.D. Variations in the cell wall composition of maize brown midrib mutants. J. Agric. Food Chem. 2003, 51, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Marczewski, S.E. Methionine, homocysteine, one carbon metabolism and fetal growth. Rev. Endocr. Metab. Disord. 2012, 13, 109–119. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Protein (Crude) in Animal Feed: Combustion (Method 990.03) Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemistry International: Washington, DC, USA, 1990. [Google Scholar]
- Association of Official Analytical Chemists. Fat (Crude) or Ether Extract in Animal Feed (Method 920.29) Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemistry International: Washington, DC, USA, 1990. [Google Scholar]
- American Association of Cereal Chemists. Approved Methods of the American Association of Cereal Chemists (Method 76–13), 9th ed.; Approved Methods Committee: St. Paul, MN, USA, 1995. [Google Scholar]
- American Oil Chemists’ Society. AOCS Official Methods and Recommended Practices (Method Ca 5b71), 15th ed.; AOCS: Champaign, IL, USA, 2006. [Google Scholar]
- AOAC International. AOAC Official Method (Method 978.10) Official Methods of Analysis, 18th ed.; AOCS: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Li, J.; Walker, C.E.; Faubion, J.M. Acidulant and oven type affect total anthocyanin content of blue maize cookies. J. Sci. Food Agric. 2011, 91, 38–43. [Google Scholar] [CrossRef]
- AOAC International. AOAC Official Method (Method 994.12) Official Methods of Analysis, 15th ed.; AOCS: Gaithersburg, MD, USA, 1990. [Google Scholar]
- Morozova, M.; Elizarova, T.; Pleteneva, T. Discriminant analysis and Mahalanobis distance (NIR diffuse.reflectance spectra) in the assessment of drug’s batch-to-batch dispersion and quality threshold establishment. Eur. Sci. J. 2013, 9, 8–24. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Use R!); Springer: New York, NY, USA, 2010. [Google Scholar]
- Josse, J.; Husson, F. missMDA: A package for handling missing values in multivariate data analysis. J. Stat. Softw. 2016, 70, 1–31. [Google Scholar] [CrossRef]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.5. 2017. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 24 November 2020).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Epskamp, S.; Cramer, A.O.J.; Waldrop, L.J.; Schmittman, V.D.; Borsboom, D. qgraph: Network vizualizations of relationships in psychometric data. J. Stat. Soft. 2012, 48, 1–18. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Trait | Code | N | Min | Mean | Max | SD | CV (%) |
|---|---|---|---|---|---|---|---|
| Aspartic acid | Asp | 143 | 0.69 | 0.71 | 0.73 | 0.09 | 8.07 |
| Threonine | Thr | 143 | 0.33 | 0.34 | 0.35 | 0.04 | 8.21 |
| Glutamic acid | Glu | 143 | 1.81 | 1.93 | 1.99 | 0.27 | 9.97 |
| Proline | Pro | 143 | 0.88 | 0.93 | 0.97 | 0.13 | 13.13 |
| Glycine | Gly | 143 | 0.38 | 0.39 | 0.40 | 0.04 | 5.90 |
| Alanine | Ala | 143 | 0.79 | 0.83 | 0.86 | 0.12 | 9.56 |
| Cysteine | Cys | 143 | 0.20 | 0.21 | 0.22 | 0.02 | 6.47 |
| Valine | Val | 143 | 0.49 | 0.50 | 0.52 | 0.06 | 8.76 |
| Methionine | Met | 143 | 0.25 | 0.26 | 0.28 | 0.04 | 11.53 |
| Isoleucine | Iso | 143 | 0.34 | 0.36 | 0.37 | 0.06 | 9.07 |
| Leucine | Leu | 143 | 1.29 | 1.39 | 1.44 | 0.22 | 11.17 |
| Lysine | Lys | 143 | 0.31 | 0.32 | 0.33 | 0.03 | 5.79 |
| Total amino acids | TAA | 143 | 7.81 | 8.18 | 8.44 | 1.02 | 8.90 |
| Total fatty acids | TFA | 143 | 3.76 | 5.48 | 6.26 | 1.49 | 20.69 |
| Protein | CP | 143 | 10.35 | 10.86 | 11.29 | 1.21 | 8.15 |
| Oil | Oil | 143 | 3.89 | 5.68 | 6.49 | 1.54 | 20.97 |
| Crude fiber | CDF | 143 | 1.61 | 1.68 | 1.81 | 0.18 | 8.62 |
| Ash | Ash | 143 | 1.41 | 1.52 | 1.64 | 0.14 | 7.34 |
| Starch | STR | 143 | 55.60 | 56.47 | 57.70 | 2.83 | 4.70 |
| Anthocyanin | ANA | 143 | 16.67 | 46.23 | 58.24 | 20.57 | 23.83 |
| Trait | Accession | Location | Year | Interaction | |||
|---|---|---|---|---|---|---|---|
| A × L | A × Y | L × Y | A × L × Y | ||||
| Aspartic acid | 1.19 | 20.15 *** | 65.48 *** | 1.08 | 2.31 * | 0.03 | 0.93 |
| Threonine | 1.43 | 8.41 *** | 75.71 *** | 1.32 | 2.42 * | 9.27 ** | 0.51 |
| Glutamic acid | 1.66 | 9.88 *** | 65.14 *** | 1.28 | 1.96 | 0.27 | 0.90 |
| Proline | 0.92 | 3.50 * | 11.30 ** | 0.92 | 0.47 | 0.01 | 0.42 |
| Glycine | 1.67 | 16.37 *** | 117.43 *** | 0.81 | 1.09 | 0.78 | 0.37 |
| Alanine | 1.62 | 12.95 *** | 72.20 *** | 1.33 | 1.96 | 0.01 | 0.96 |
| Cysteine | 1.94 | 28.65 *** | 90.36 *** | 1.60 | 0.87 | 3.73 | 0.64 |
| Valine | 1.05 | 4.54 ** | 55.78 *** | 0.95 | 1.86 | 6.59 | 1.09 |
| Methionine | 3.00 ** | 11.96 *** | 81.99 *** | 0.95 | 1.09 | 13.52 *** | 1.01 |
| Isoleucine | 1.21 | 8.25 *** | 107.05 *** | 1.24 | 2.06 | 8.32 *** | 1.29 |
| Leucine | 1.59 | 12.64 *** | 60.73 *** | 1.33 | 1.83 | 0.18 | 1.15 |
| Lysine | 4.00 *** | 17.85 *** | 14.60 *** | 0.48 | 1.75 | 0.11 | 0.47 |
| Total amino acids | 1.20 | 9.37 *** | 66.70 *** | 1.18 | 1.76 | 0.24 | 0.90 |
| Total fatty acids | 9.29 *** | 2.01 | 3.19 | 1.17 | 1.25 | 3.37 | 2.13 * |
| Protein | 1.62 | 15.33 *** | 44.39 *** | 0.94 | 1.74 | 1.00 | 0.83 |
| Oil | 9.29 *** | 2.00 | 3.15 | 1.16 | 1.25 | 3.34 | 2.14 * |
| Crude fiber | 4.03 *** | 4.35 ** | 16.33 *** | 1.08 | 0.36 | 17.26 *** | 0.70 |
| Ash | 7.48 *** | 6.07 *** | 2.03 | 0.82 | 0.86 | 0.00 | 1.31 |
| Starch | 1.69 | 5.19 ** | 4.43 * | 0.73 | 1.30 | 0.09 | 0.91 |
| Anthocyanin | 18.04 *** | 18.88 *** | 14.57 *** | 2.70 *** | 2.97 ** | 16.17 *** | 3.70 ** |
| Constituent | NIR | LAB | RMSEP | |
|---|---|---|---|---|
| PCR | PLS | |||
| Protein (%) | 0.123 | 0.102 | 1.11 | 1.05 |
| Oil (%) | 0.107 | 0.129 | 1.44 | 1.43 |
| Starch (%) | 0.170 | 0.238 | 2.81 | 2.80 |
| Lysine (%) | 0.009 | 0.002 | 0.02 | 0.02 |
| Methionine (%) | 0.007 | 0.004 | 0.33 | 0.03 |
| Cysteine (%) | 0.011 | 0.002 | 0.28 | 0.02 |
| PCR | PLS | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Calibration | Validation | Calibration | Validation | |||||||||
| Trait | R2cv | RMSEC | R2cal | R2val | RMSEP | RPD | R2cv | RMSEC | R2cal | R2val | RMSEP | RPD |
| Asp | 0.37 | 0.08 | 0.19 | 0.14 | 0.08 | 0.93 | 0.49 | 0.07 | 0.30 | 0.25 | 0.07 | 0.81 |
| Thr | 0.31 | 0.04 | 0.17 | 0.10 | 0.04 | 0.91 | 0.44 | 0.04 | 0.26 | 0.20 | 0.04 | 0.91 |
| Glu | 0.34 | 0.25 | 0.25 | 0.25 | 0.12 | 0.44 | 0.48 | 0.23 | 0.28 | 0.23 | 0.24 | 0.89 |
| Pro | 0.04 | 0.13 | 0.0001 | NA | 0.13 | 1.03 | 0.22 | 0.13 | 0.002 | NA | 0.13 | 1.03 |
| Gly | 0.51 | 0.03 | 0.31 | 0.27 | 0.03 | 0.83 | 0.59 | 0.03 | 0.38 | 0.35 | 0.03 | 0.83 |
| Ala | 0.38 | 0.19 | 0.10 | 0.15 | 0.11 | 0.95 | 0.49 | 0.10 | 0.30 | 0.25 | 0.10 | 0.86 |
| Cys | 0.47 | 0.02 | 0.28 | 0.23 | 0.02 | 0.83 | 0.49 | 0.02 | 0.29 | 0.24 | 0.02 | 0.83 |
| Val | 0.22 | 0.10 | 0.06 | 0.04 | 0.06 | 0.98 | 0.43 | 0.05 | 0.24 | 0.18 | 0.05 | 0.82 |
| Met | 0.51 | 0.04 | 0.33 | 0.27 | 0.04 | 0.93 | 0.60 | 0.03 | 0.40 | 0.36 | 0.03 | 0.70 |
| Iso | 0.39 | 0.05 | 0.19 | 0.15 | 0.05 | 0.91 | 0.54 | 0.05 | 0.34 | 0.29 | 0.05 | 0.91 |
| Leu | 0.33 | 0.20 | 0.15 | 0.12 | 0.21 | 0.95 | 0.45 | 0.19 | 0.27 | 0.21 | 0.20 | 0.90 |
| Lys | 0.21 | 0.02 | 0.08 | 0.05 | 0.02 | 0.80 | 0.30 | 0.02 | 0.16 | 0.09 | 0.02 | 0.80 |
| TAA | 0.33 | 0.93 | 0.18 | 0.12 | 0.97 | 0.95 | 0.49 | 0.86 | 0.29 | 0.25 | 0.89 | 0.87 |
| TFA | 0.31 | 1.39 | 0.12 | 0.11 | 1.41 | 0.95 | 0.32 | 1.38 | 0.13 | 0.12 | 1.40 | 0.94 |
| CP | 0.31 | 1.11 | 0.15 | 0.10 | 1.15 | 0.95 | 0.45 | 1.05 | 0.24 | 0.20 | 1.09 | 0.17 |
| Oil | 0.30 | 1.44 | 0.12 | 0.10 | 1.46 | 0.95 | 0.33 | 1.43 | 0.13 | 0.11 | 1.45 | 0.94 |
| CDF | 0.20 | 0.17 | 0.09 | 0.04 | 0.18 | 0.99 | 0.30 | 0.14 | 0.17 | 0.10 | 0.17 | 0.94 |
| Ash | 0.18 | 0.13 | 0.05 | 0.04 | 0.13 | 0.96 | 0.23 | 0.13 | 0.08 | 0.06 | 0.13 | 0.96 |
| STR | 0.45 | 2.81 | 0.0001 | NA | 2.86 | 1.01 | 0.16 | 2.8 | 0.01 | NA | 2.86 | 1.01 |
| ANA | 0.25 | 19.2 | 0.13 | 0.07 | 19.9 | 0.97 | 0.37 | 18.9 | 0.15 | 0.10 | 19.6 | 0.95 |
| PCR | PLS | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Calibration | Validation | Calibration | Validation | |||||||||
| Trait | R2cv | RMSEC | R2cal | R2val | RMSEP | RPD | R2cv | RMSEC | R2cal | R2val | RMSEP | RPD |
| Asp | 0.37 | 0.08 | 0.19 | 0.14 | 0.08 | 0.93 | 0.52 | 0.07 | 0.30 | 0.27 | 0.07 | 0.81 |
| Thr | 0.31 | 0.04 | 0.14 | 0.10 | 0.04 | 0.91 | 0.47 | 0.04 | 0.27 | 0.23 | 0.04 | 0.91 |
| Glu | 0.34 | 0.25 | 0.17 | 0.25 | 0.12 | 0.44 | 0.51 | 0.23 | 0.29 | 0.26 | 0.23 | 0.85 |
| Pro | 0.04 | 0.13 | 0.001 | NA | 0.13 | 1.03 | 0.10 | 0.12 | 0.3 | NA | 0.13 | 1.03 |
| Gly | 0.51 | 0.03 | 0.30 | 0.27 | 0.03 | 0.83 | 0.59 | 0.03 | 0.39 | 0.36 | 0.03 | 0.83 |
| Ala | 0.38 | 0.10 | 0.19 | 0.15 | 0.11 | 0.95 | 0.54 | 0.10 | 0.31 | 0.30 | 0.10 | 0.86 |
| Cys | 0.47 | 0.02 | 0.28 | 0.23 | 0.02 | 0.83 | 0.52 | 0.02 | 0.33 | 0.27 | 0.02 | 0.83 |
| Val | 0.22 | 0.10 | 0.06 | 0.04 | 0.06 | 0.98 | 0.45 | 0.05 | 0.25 | 0.24 | 0.05 | 0.82 |
| Met | 0.51 | 0.03 | 0.33 | 0.27 | 0.04 | 0.93 | 0.36 | 0.03 | 0.40 | 0.36 | 0.03 | 0.70 |
| Iso | 0.38 | 0.05 | 0.19 | 0.15 | 0.05 | 0.91 | 0.56 | 0.04 | 0.37 | 0.32 | 0.05 | 0.91 |
| Leu | 0.33 | 0.20 | 0.15 | 0.12 | 0.21 | 0.95 | 0.47 | 0.19 | 0.27 | 0.20 | 0.22 | 1.00 |
| Lys | 0.21 | 0.02 | 0.10 | 0.05 | 0.02 | 0.80 | 0.31 | 0.02 | 0.15 | 0.10 | 0.02 | 0.80 |
| TAA | 0.33 | 0.92 | 0.18 | 0.12 | 0.97 | 0.95 | 0.51 | 0.85 | 0.30 | 0.27 | 0.88 | 0.86 |
| TFA | 0.31 | 1.39 | 0.12 | 0.11 | 1.41 | 0.95 | 0.30 | 1.39 | 0.12 | 0.10 | 1.41 | 0.95 |
| CP | 0.31 | 1.11 | 0.15 | 0.10 | 1.15 | 0.95 | 0.45 | 1.05 | 0.25 | 0.22 | 1.08 | 0.89 |
| Oil | 0.30 | 1.44 | 0.12 | 0.10 | 1.46 | 0.95 | 0.30 | 1.44 | 0.12 | 0.11 | 1.46 | 0.95 |
| CDF | 0.20 | 0.17 | 0.09 | 0.04 | 0.18 | 0.99 | 0.31 | 0.17 | 0.14 | 0.10 | 0.17 | 0.94 |
| Ash | 0.18 | 0.13 | 0.05 | 0.04 | 0.13 | 0.96 | 0.19 | 0.13 | 0.06 | 0.04 | 0.13 | 0.96 |
| STR | 0.45 | 2.81 | 0.001 | NA | 2.86 | 1.01 | 0.23 | 2.77 | 0.03 | NA | 2.90 | 1.02 |
| ANA | 0.25 | 19.2 | 0.13 | 0.07 | 19.9 | 0.97 | 0.27 | 18.9 | 0.15 | 0.08 | 19.9 | 0.97 |
| Principal Components | Eigenvalue | Variance (%) | Cumulative Variance (%) |
|---|---|---|---|
| 1 | 9.92 | 49.62 | 49.62 |
| 2 | 4.43 | 22.20 | 71.77 |
| 3 | 1.40 | 6.98 | 78.75 |
| 4 | 1.30 | 6.51 | 85.26 |
| 5 | 0.90 | 4.48 | 89.74 |
| 6 | 0.67 | 3.32 | 93.06 |
| 7 | 0.38 | 1.92 | 94.98 |
| 8 | 0.30 | 1.52 | 96.50 |
| 9 | 0.20 | 1.09 | 97.59 |
| 10 | 0.15 | 0.73 | 98.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nankar, A.N.; Scott, M.P.; Pratt, R.C. Compositional Analyses Reveal Relationships among Components of Blue Maize Grains. Plants 2020, 9, 1775. https://doi.org/10.3390/plants9121775
Nankar AN, Scott MP, Pratt RC. Compositional Analyses Reveal Relationships among Components of Blue Maize Grains. Plants. 2020; 9(12):1775. https://doi.org/10.3390/plants9121775
Chicago/Turabian StyleNankar, Amol N., M Paul Scott, and Richard C. Pratt. 2020. "Compositional Analyses Reveal Relationships among Components of Blue Maize Grains" Plants 9, no. 12: 1775. https://doi.org/10.3390/plants9121775