Significant Accumulation of Iodine and Selenium in Chicory (Cichorium intybus L. var. foliosum Hegi) Leaves after Foliar Spraying

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Yield of Red Chicory
2.2. Selenium Content
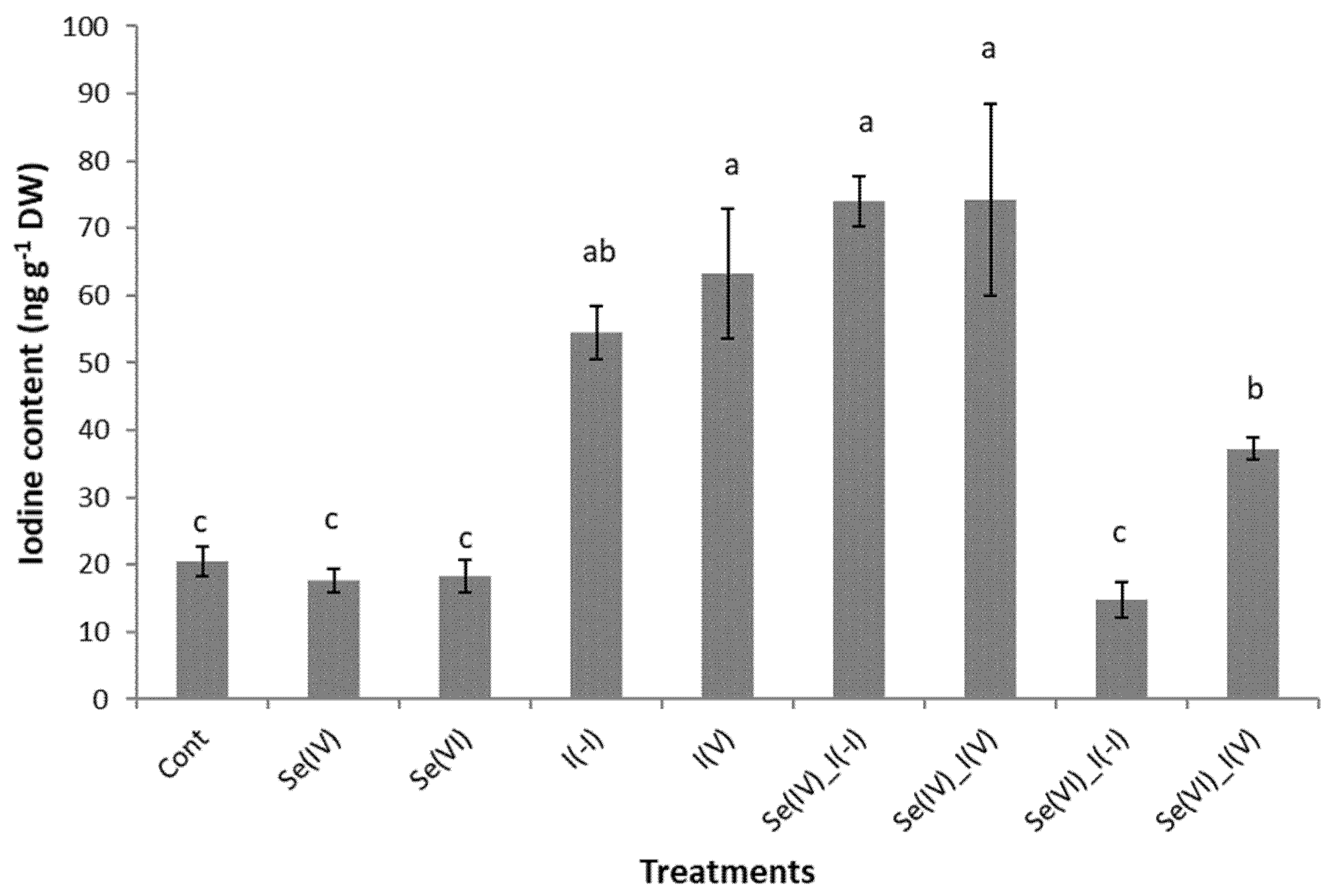
2.3. Iodine Content
2.4. Biochemical and Physiological Plant Parameters
3. Materials and Methods
3.1. Plant Samples
3.2. Determination of Selenium and Iodine Contents
3.3. Determination of Biochemical and Physiological Parameters
3.4. Statistical Analysis
4. Conclusions
- Selenium content increased in the Se and Se plus I foliar-treated plants.
- Iodine content increased in chicory plants treated with I and I plus Se, except for the of selenate and iodide combination.
- Treatment with Se and I is promising way to increase their concentrations in crops, and consequently in humans, without reducing yields. Both forms of I lowered Se content when Se was added together with I in the form of Se (VI), and the same form of Se lowered the amount of I when both were added simultaneously.
- The potential photochemical efficiency of PSII showed the good vitality of the plants.
- Different I and Se treatments did not have any significant effects on the yield and mass of the chicory heads.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schomburg, L.; Köhrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Abdalla, N.; Alshaal, T.; El-Henawy, A.; Faizy, S.E.-D.A.; Shams, M.S.; Domokos-Szabolcsy, É. Selenium and Its Role in Higher Plants; Springer International Publishing: Cham, Switzerland, 2015; pp. 235–296. [Google Scholar] [CrossRef]
- Jerše, A.; Kacjan Maršić, N.; Kroflič, A.; Germ, M.; Šircelj, H.; Stibilj, V. Is foliar enrichment of pea plants with iodine and selenium appropriate for production of functional food. Food Chem. 2018, 267, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Krzepiłko, A.; Prażak, R.; Skwaryło-Bednarz, B.; Molas, J. Agronomic biofortifcation as a means of enriching plant foodstufs with iodine. Acta Agrobot. 2019, 72, 1–9. [Google Scholar] [CrossRef]
- Allen, L.; de Benoist, D.; Dary, O.; Hurrell, R. Guidelines on Food Fortification with Micronutrients Geneva: World Health Organization. 2006. Available online: http://www.who.int/iris/handle/10665/43412 (accessed on 13 November 2020).
- Zimmermann, M.B.; Jooste, P.L.; Pandav, C.S. Iodine-Deficiency disorders. Lancet 2008, 372, 1251–1262. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine biofortification of crops: Agronomic biofortification, metabolic engineering and iodine bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, O.S.; Young, S.D.; Bailey, E.H.; Crout, N.M.J.; Ander, E.L.; Hamilton, E.M.; Watts, M.J. Iodine uptake, storage and translocation mechanisms in spinach (Spinacia oleracea L.). Environ. Geochem. Health 2019, 41, 2145–2156. [Google Scholar] [CrossRef]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Hawrylak-Nowak, B.; Hasanuzzaman, M.; Matraszek-Gawron, R. Mechanisms of selenium-induced enhancement of abiotic stress tolerance in plants. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Ed.; Springer Nature: Singapore, 2018; Volume 269. [Google Scholar] [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.M.; Liang, Y.L.; Gao, D.K.; An, X.J.; Kong, F.C. Spraying foliar selenium fertilizer on quality of table grape (Vitisvinifera L.) from different source varieties. Sci. Hortic. Amst. 2017, 218, 87–94. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Ertani, A.; Pilon-Smits, E.A.H.; Fabrega-Prats, M.; Schiavon, M. Selenium biofortification differentially affects sulfur metabolism and accumulation of phytochemicals in two rocket species (Eruca sativa Mill. and Diplotaxis tenuifolia) grown in hydroponics. Plants 2019, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Cartes, P.; Gianfreda, L.; Paredes, C.; Mora, M.L. Selenium uptake and its antioxidant role in ryegrass cultivars as affected by selenite seed pelletization. J. Soil Sci. Plant Nutr. 2011, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.G.; Huang, Y.; Hu, Y.; Liu, Y.; Christie, P. Interactions between selenium and iodine uptake by spinach (Spinacia oleracea L.) in solution culture. Plant Soil 2004, 261, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Bieżanowska-Kopeć, R.; Pysz, M.; Kapusta-Duch, J.; Aneta, K.; Sylwester, S.; Koronowicz, A.; Piątkowska, E.; Rakoczy, R.; Skoczylas, L.; Leszczyńska, T. The effects of peeling and cooking on the mineral content and antioxidant properties in carrots enriched with potassium iodate and/or selenite (SeIV) and selenite (SeVI). Int. J. Food Sci. Nutr. 2016, 67, 919–928. [Google Scholar] [CrossRef]
- Smoleń, S.; Skoczylas, Ł.; Ledwożyw-Smoleń, I.; Rakoczy, R.; Kopeć, A.; Piątkowska, E.; Kapusta-Duch, J. Biofortification of carrot (Daucus carota L.) with iodine and selenium in a field experiment. Front. Plant Sci. 2016, 7, 730. [Google Scholar] [CrossRef] [Green Version]
- Smoleń, S.; Kowalska, I.; Sady, W. Assessment of biofortification with iodine and selenium of lettuce cultivated in the NFT hydroponic system. Sci. Hortic. Amst. 2014, 166, 9–16. [Google Scholar] [CrossRef]
- Jerše, A.; Kacjan Maršić, N.; Šircelj, H.; Germ, M.; Kroflič, A.; Stibilj, V. Seed soaking in I and Se solutions increases concentrations of both elements and changes morphological and some physiological parameters of pea sprouts. Plant. Physiol. Biochem. 2017, 118, 285–294. [Google Scholar] [CrossRef]
- Golubkina, N.; Kekina, H.; Caruso, G. Yield, quality and antioxidant properties of indian mustard (Brassica juncea L.) in response to foliar biofortification with selenium and iodine. Plants 2018, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Kacjan Maršić, N. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Kacjan Maršić, N.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes) and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef]
- Nwafor, I.C.; Shale, K.; Achilonu, M.C. Chemical composition and nutritive benefits of chicory (Cichorium intybus) as an ideal complementary and/or alternative livestock feed supplement. Sci. World J. 2017. [Google Scholar] [CrossRef] [Green Version]
- Rossetto, M.; Lante, A.; Vanzani, P.; Spettoli, P.; Scarpa, M.; Rigo, A. Red chicories as potent scavengers of highly reactive radicals: A study on their phenolic composition and peroxyl radical trapping capacity and efficiency. J. Agric. Food Chem. 2005, 53, 8169–8175. [Google Scholar] [CrossRef]
- Statistical Office of the Republic of Slovenia. Statistični letopis Republike Slovenije. In Statistical Yearbook of the Republic of Slovenia; Statistical Office of the Republic of Slovenia: Ljubljana, Slovenia, 2013; p. 304. [Google Scholar]
- Pelko, N. Bilanca Proizvodnje in Potrošnje Zelenjave na leto in Kratko Razmišljanje Načrtovanju Pridelave Zelenjave in Krompirja v letu 2020. 2020. Available online: https://www.kgzs.si/uploads/slike/bilanca_proizvodnje_in_potrosnje_zelenjave_2_clanek_splet.pdf (accessed on 4 November 2020).
- Germ, M.; Stibilj, V.; Osvald, J.; Kreft, I. Effect of selenium foliar application on chicory (Cichorium intybus L). J. Agric. Food Chem. 2007, 55, 795–798. [Google Scholar] [CrossRef]
- Blasco, B.; Rios, J.J.; Leyva, R.; Melgarejo, R.; Constan-Aguilar, C.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Romero, L.; Ruiz, J.M. Photosynthesis and metabolism of sugars from lettuce plants (Lactuca sativa L. var. longifolia) subjected to biofortification with iodine. Plant Growth Regul. 2011, 65, 137–143. [Google Scholar] [CrossRef]
- Golob, A.; Kroflič, A.; Jerše, A.; Kacjan Maršić, N.; Šircelj, H.; Stibilj, V.; Germ, M. Response of pumpkin to different concentrations and forms of selenium and iodine, and their combinations. Plants 2020, 9, 899. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Barickman, T.; Deyton, D.E.; Kopsell, D.E. Selenization of basil and cilantro through foliar applications of selenate-selenium and selenite-selenium. Hortic. Sci. Hortic. 2009, 44, 438–442. [Google Scholar] [CrossRef]
- Kacjan Maršič, N.; Golob, A.; Šircelj, H.; Mihorič, M.; Kroflič, A.; Stibilj, V.; Germ, M. Effects of exogenous selenium in different concentrations and forms on selenium accumulation and growth of Spinach (Spinacia oleracea L.). J. Agric. Sci. Technol. Iran 2020, 21, 1905–1917. [Google Scholar]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in higher plants. Annu. Rev. Plant Physiol. Plant. Mol. Biol. 2000, 51, 401. [Google Scholar] [CrossRef] [Green Version]
- Smoleń, S.; Skoczylas, Ł.; Ledwożyw-Smoleń, I.; Rakoczy, R.; Kopeć, A.; Piątkowska, E.; Bieżanowska-Kopeć, R.; Pysz, M.; Koronowicz, A.; Kapusta-Duch, J.; et al. Iodine and selenium biofortification of lettuce (Lactuca sativa L.) by soil fertilization with various compounds of these elements. Acta Sci. Pol. Hortorum Cultus 2016, 15, 69–91. [Google Scholar]
- Packard, T.T. Measurement of electron transport activity of microplankton. In Advances in Aquatic Microbiology; Jannasch, H.W., Williams, P.J., Eds.; Academic Press Inc. Ltd.: London, UK, 1985; pp. 207e–262e. [Google Scholar]
- Gaberščik, A.; Germ, M.; Škof, A.; Drmaž, D.; Trošt Sedej, T. UV-B radiation screen and respiratory potential in two aquatic primary producers: Scenedesmus quadricauda and Ceratophyllum demersum. In Verhandlungen; Internationale Vereinigung für Theoretische und Angewandte Limnologie: Stuttgart, Germany, 2002; pp. 422e–425e. [Google Scholar]
- Bartoli, C.G.; Gomez, F.; Gergoff, G.; Guiam, J.J.; Puntarulo, S. Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions. J. Exp. Bot. 2005, 56, 1269–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, S.; Wachi, T.; Yoshihira, K.; Nakagawa, T.; Ishikawa, A.; Takagi, D.; Tezuka, A.; Yoshida, H.; Yoshida, S.; Sekimoto, H.; et al. Rice (Oryza sativa L.) roots have iodate reduction activity in response to iodine. Front. Plant Sci. 2013, 4, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, U.; Kohl, M.; Klimant, I.; Reising, H. Measurement of chlorophyll fluorescence within leaves using a modified PAM Fluorometer with a fiber-optic microprobe. Photosynth. Res. 1996, 47, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Ćustić, M.; Poljak, M.; Čoga, L.; Ćosić, T.; Toth, N.; Pecina, M. The influence of organic and mineral fertilization on nutrient status, nitrate accumulation, and yield of head chicory. Plant Soil Environ. 2003, 49, 218–222. [Google Scholar] [CrossRef]
- Bortolini, L.; Tolomio, M. Influence of irrigation frequency on radicchio (Cichorium Intybus L.) yield. Water 2019, 11, 2473. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.gov.si/teme/integrirana-pridelava/19.11.2020 (accessed on 19 November 2020).
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. In Current Protocols in Food Analytical Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of phtosynthetic tissues: Chlorophylls and carotenoids. In Current Protocols in Food Analytical Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Kenner, R.A.; Ahmed, S.I. Measurements of electron transport activities in marine phytoplankton. Mar. Biol. 1975, 33, 119–127. [Google Scholar] [CrossRef]
- Packard, T. The measurement of electron transport activity in marine phytoplankton. J. Mar. Res. 1971, 29, 235–244. [Google Scholar]



{kind=link}
{kind=link}
| Treatment | Yield of Chicory (t/ha) | Mass of Above-Ground Plant Parts (g) | Number of Removed Leaves | Marketable Mass of Chicory Heads (g) |
|---|---|---|---|---|
| Control | 48.4 ± 4.0 a | 1148 ± 102 a | 17 ± 2 a | 726 ± 59 a |
| I (−I) | 40.1 ± 2.0 a | 1009 ± 49 a | 20 ± 2 a | 602 ± 30 a |
| I (V) | 43.5 ± 4.1 a | 1043 ± 86 a | 19 ± 1 a | 653 ± 62 a |
| Se (IV) | 51.5 ± 1.4 a | 1199 ± 37 a | 19 ± 1 a | 766 ± 22 a |
| Se (VI) | 46.9 ± 4.3 a | 1079 ± 68 a | 17 ± 1 a | 704 ± 64 a |
| Se (IV) + I (−I) | 50.1 ± 4.0 a | 1176 ± 87 a | 18 ± 2 a | 752 ± 60 a |
| Se (IV) + I (V) | 50.5 ± 3.8 a | 1174 ± 87 a | 17 ± 1 a | 758 ± 57 a |
| Se (VI) + I (−I) | 43.9 ± 1.9 a | 1097 ± 69 a | 19 ± 1 a | 659 ± 29 a |
| Se (VI) + I (V) | 39.8 ± 5.7 a | 970 ± 125 a | 16 ± 2 a | 597 ± 85 a |
| Treatment | Chlorophyll a (mg g−1 DW) | Chlorophyll b (mg g−1 DW) | Carotenoids (mg g−1 DW) | ETS Activity (μL O2 mg−1 DW h) | Fv/Fm |
|---|---|---|---|---|---|
| Control | 16.6 ± 1.3 ab | 13.1 ± 1.1 ab | 4.4 ± 0.3 ab | 16.4 ± 0.9 de | 0.74 ± 0.02 a |
| I (−I) | 15.1 ± 2.2 abc | 9.32 ± 1.4 cde | 4.1 ± 0.6 abc | 16.5 ± 0.4 de | 0.74 ± 0.01 a |
| I (V) | 12.7 ± 1.2 c | 6.90 ± 0.8 e | 3.3 ± 0.2 c | 18.7 ± 0.7 abc | 0.75 ± 0.01 a |
| Se (IV) | 14.1 ± 1.5 bc | 11.1 ± 1.3 bcd | 3.5 ± 0.3 bc | 16.9 ± 0.8 cde | 0.76 ± 0.01 a |
| Se (VI) | 18.9 ± 0.8 a | 14.4 ± 0.9 a | 5.0 ± 0.2 a | 20.2 ± 0.7 ab | 0.77 ± 0.02 a |
| Se (IV) + I (−I) | 15.0 ± 2.3 abc | 9.04 ± 1.4 cde | 4.1 ± 0.5 abc | 19.4 ± 1.5 ab | 0.77 ± 0.01 a |
| Se (IV) + I (V) | 16.4 ± 0.5 abc | 10.1 ± 1.2 bcd | 4.2 ± 0.2 abc | 20.7 ± 0.6 a | 0.75 ± 0.01 a |
| Se (VI) + I (−I) | 13.3 ± 0.8 bc | 8.14 ± 0.5 de | 3.3 ± 0.2 c | 16.1 ± 0.8 e | 0.76 ± 0.02 a |
| Se (VI) + I (V) | 16.4 ± 0.5 abc | 11.2 ± 0.3 bc | 4.3 ± 0.1 ab | 18.3 ± 0.3 bcd | 0.74 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germ, M.; Kacjan-Maršić, N.; Kroflič, A.; Jerše, A.; Stibilj, V.; Golob, A. Significant Accumulation of Iodine and Selenium in Chicory (Cichorium intybus L. var. foliosum Hegi) Leaves after Foliar Spraying. Plants 2020, 9, 1766. https://doi.org/10.3390/plants9121766
Germ M, Kacjan-Maršić N, Kroflič A, Jerše A, Stibilj V, Golob A. Significant Accumulation of Iodine and Selenium in Chicory (Cichorium intybus L. var. foliosum Hegi) Leaves after Foliar Spraying. Plants. 2020; 9(12):1766. https://doi.org/10.3390/plants9121766
Chicago/Turabian StyleGerm, Mateja, Nina Kacjan-Maršić, Ana Kroflič, Ana Jerše, Vekoslava Stibilj, and Aleksandra Golob. 2020. "Significant Accumulation of Iodine and Selenium in Chicory (Cichorium intybus L. var. foliosum Hegi) Leaves after Foliar Spraying" Plants 9, no. 12: 1766. https://doi.org/10.3390/plants9121766