Epipactis tremolsii Seed Diversity in Two Close but Extremely Different Populations: Just a Case of Intraspecific Variability?

 , and
, and 
Abstract
:1. Introduction
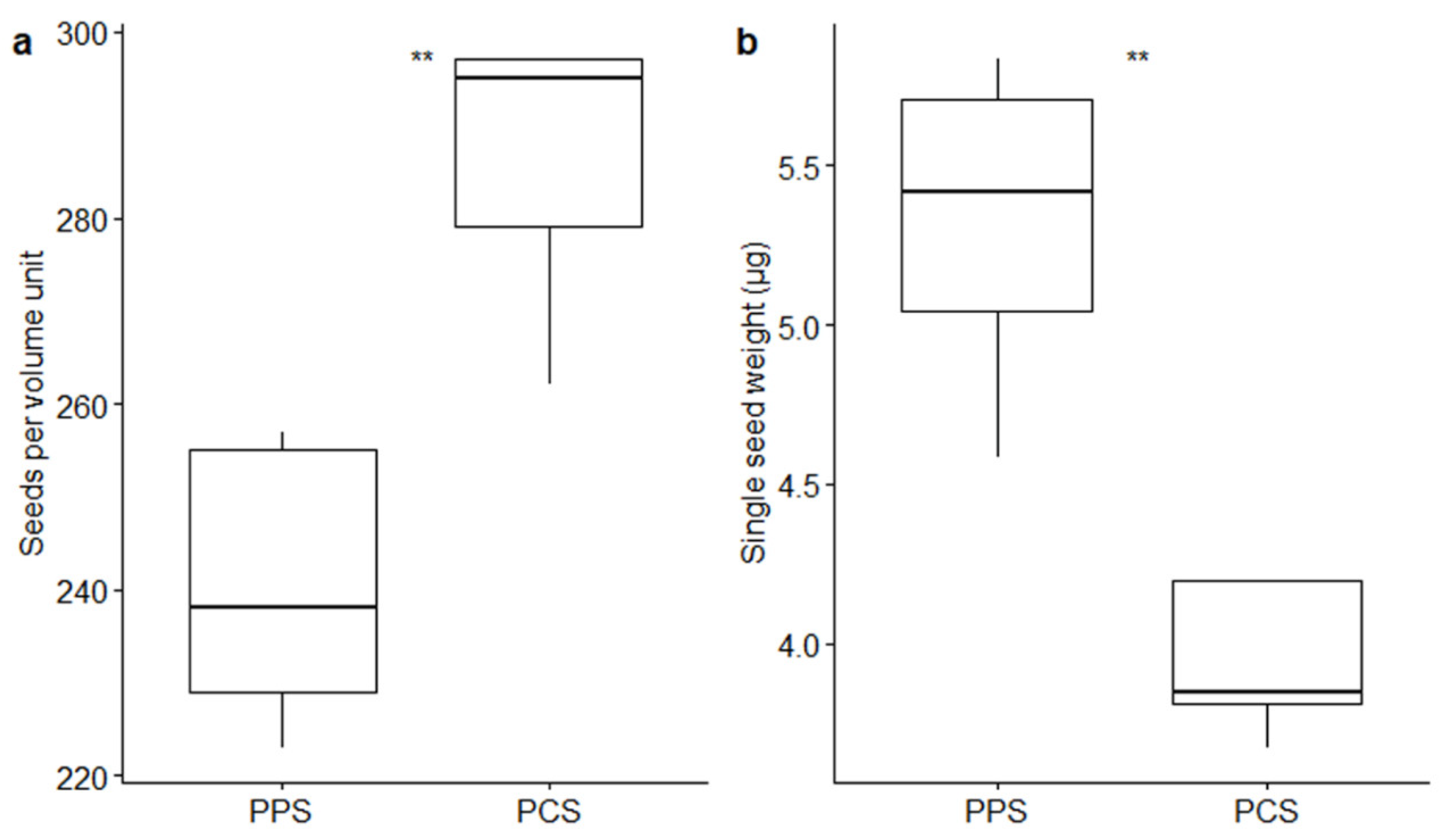
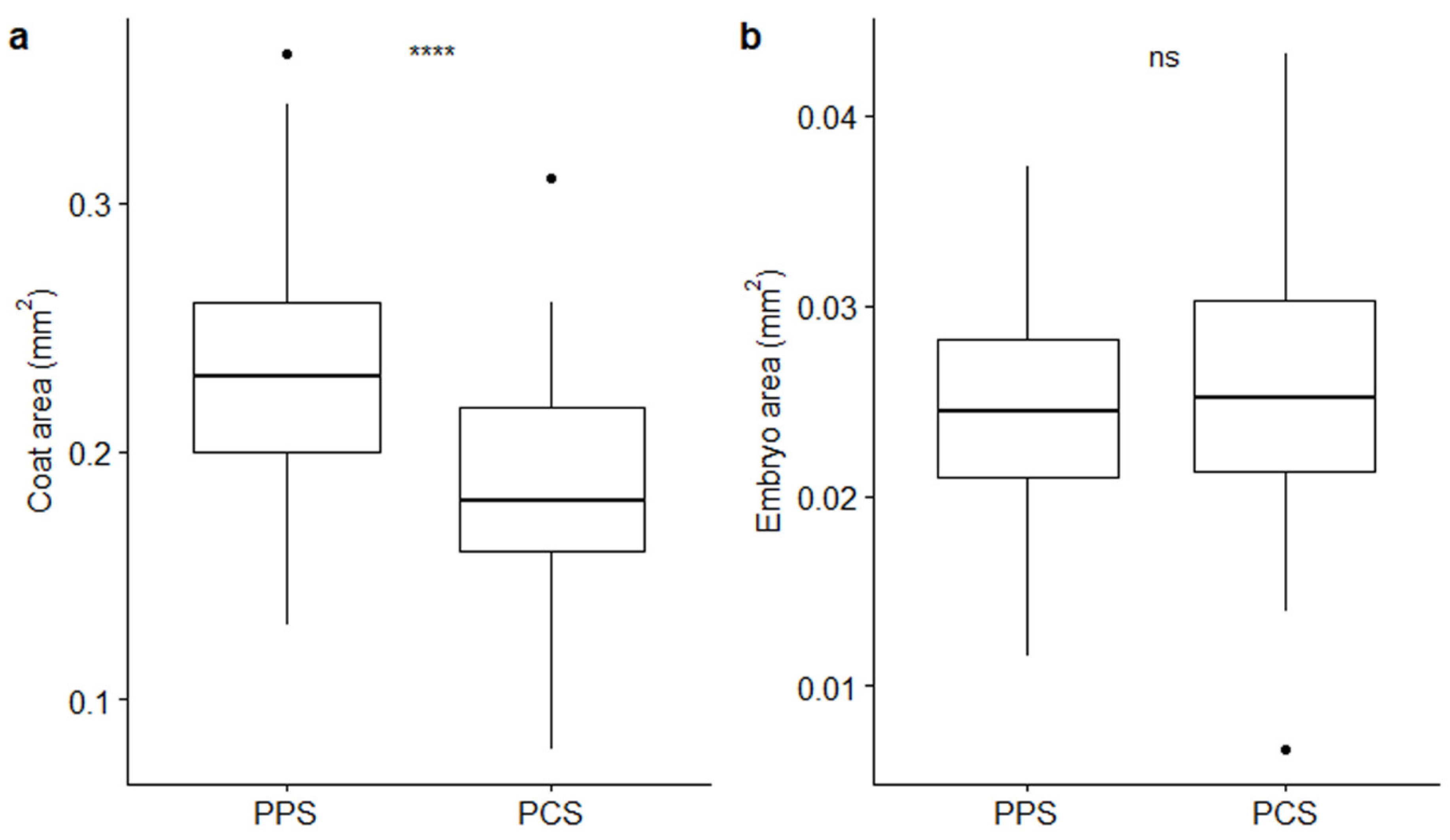
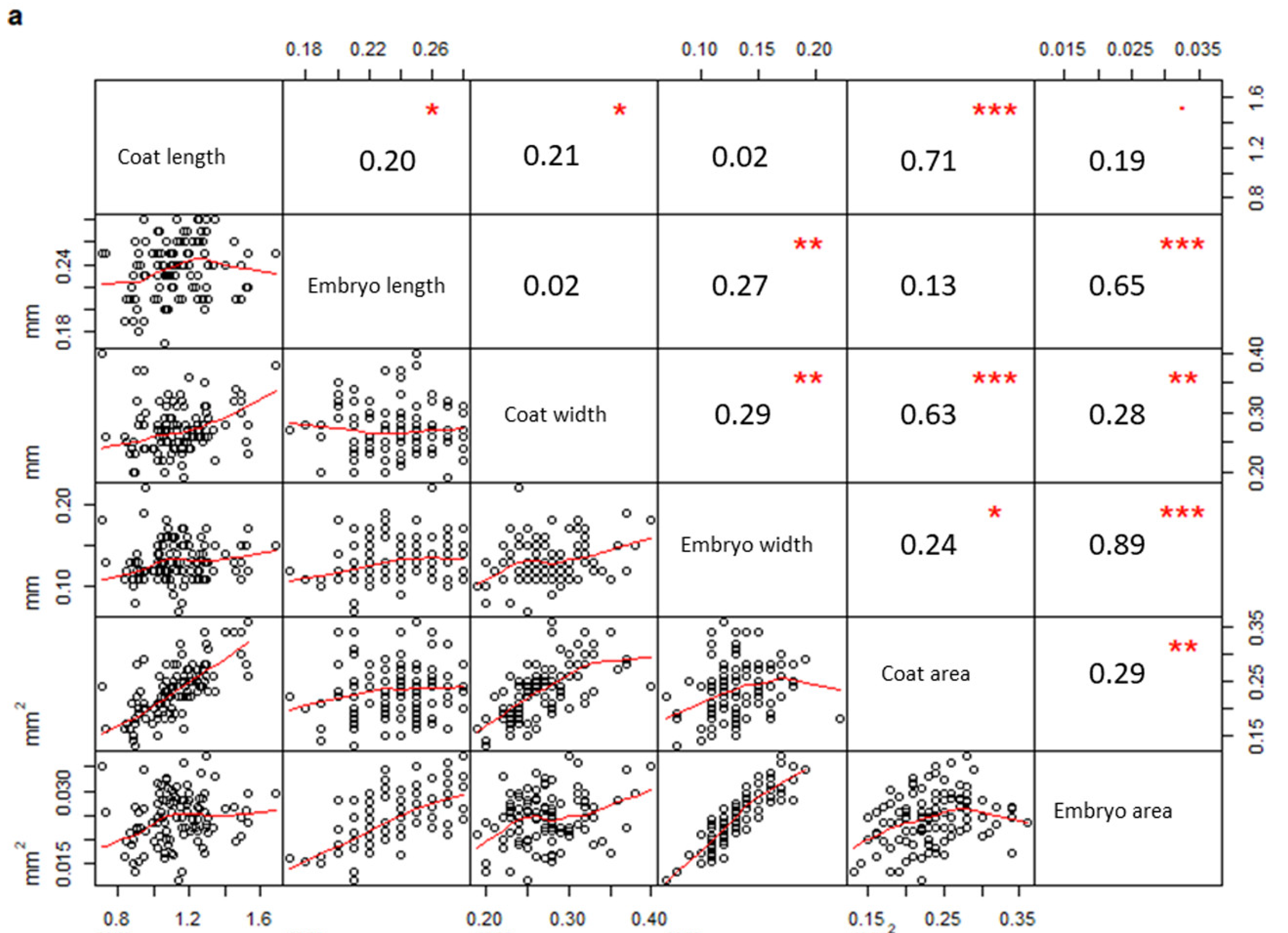
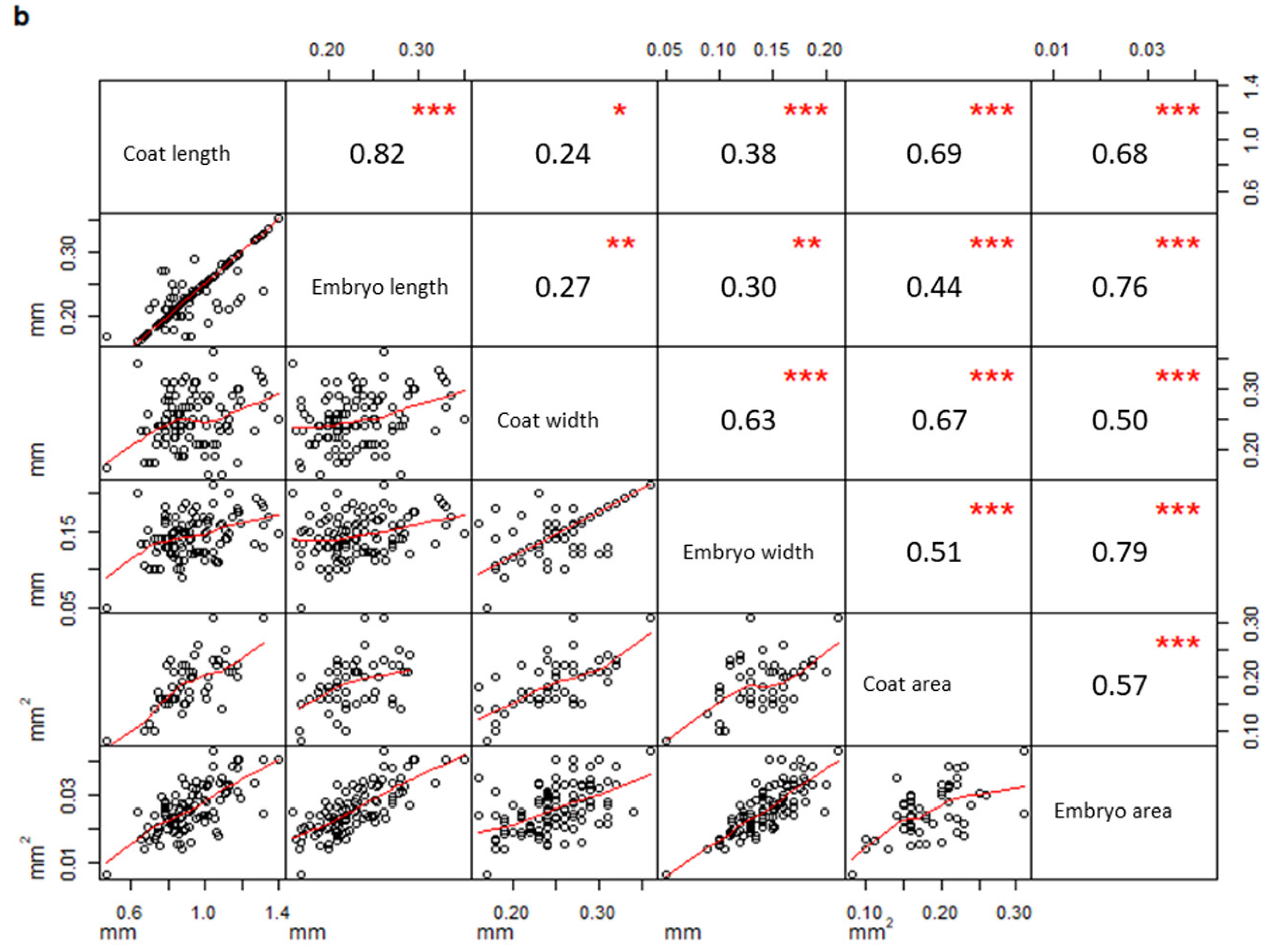
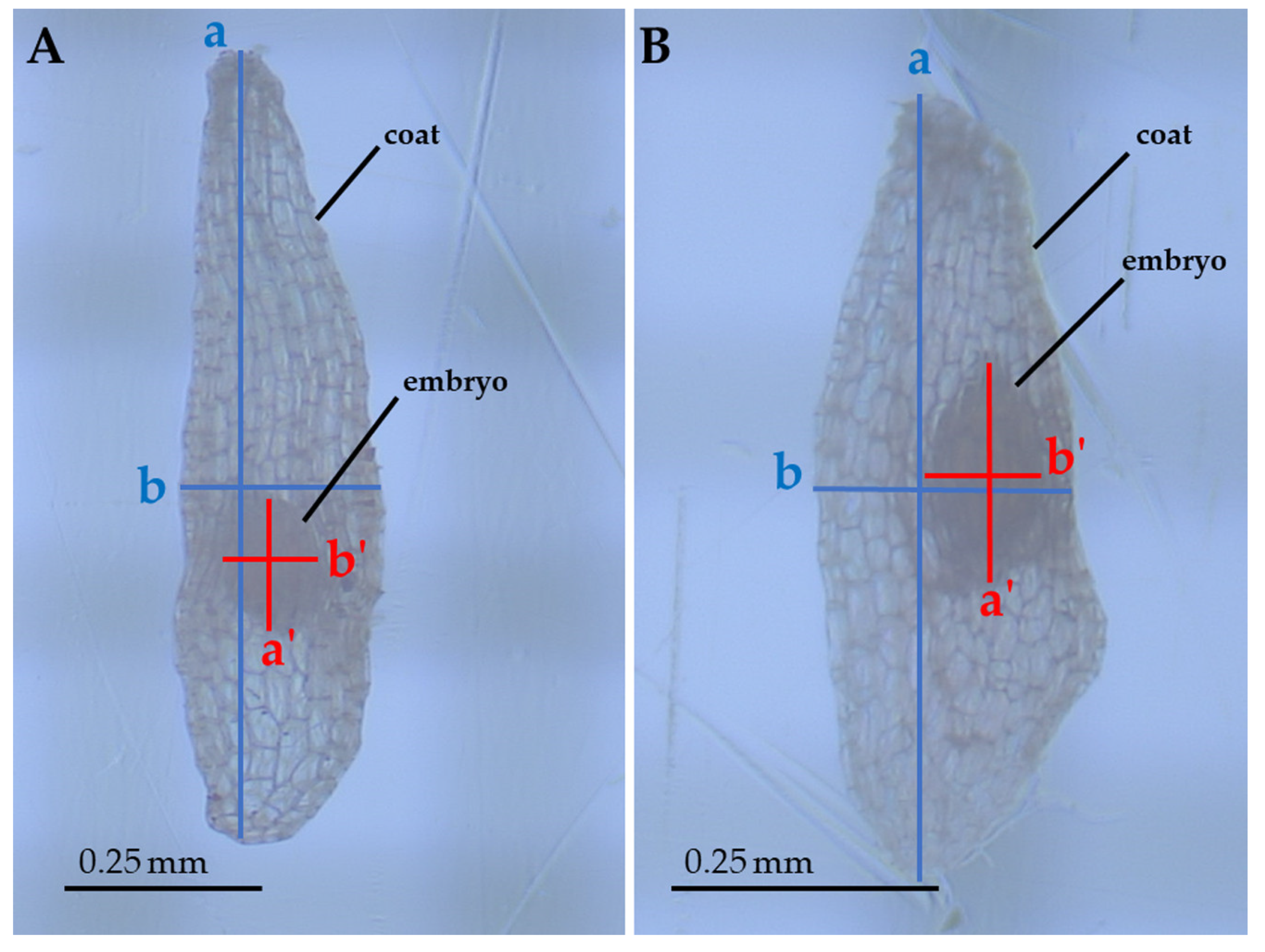
2. Results
3. Discussion
4. Materials and Methods
4.1. Species Description
4.2. Data Collection
4.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lussu, M.; De Agostini, A.; Cogoni, A.; Marignani, M.; Cortis, P. Does size really matter? A comparative study on floral traits in orchids with two different pollination strategies. Plant Biol. 2019, 21, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Gögler, J.; Stökl, J.; Cortis, P.; Beyrle, H.; Lumaga, M.R.B.; Cozzolino, S.; Ayasse, M. Increased divergence in floral morphology strongly reduces gene flow in sympatric sexually deceptive orchids with the same pollinator. Evol. Ecol. 2015, 29, 703–717. [Google Scholar] [CrossRef]
- Arditti, J.; Elliott, J.; Kitching, I.J.; Wasserthal, L.T. ‘Good Heavens what insect can suck it’- Charles Darwin, Angraecum sesquipedale and Xanthopan morganii praedicta. Bot. J. Linn. Soc. 2012, 169, 403–432. [Google Scholar] [CrossRef] [Green Version]
- Schiestl, F.P. On the success of a swindle: Pollination by deception in orchids. Naturwissenschaften 2005, 92, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Pellegrino, G.; Bellusci, F.; Musacchio, A. The effects of inflorescence size and flower position on female reproductive success in three deceptive orchids. Bot. Stud. 2010, 51, 351–356. [Google Scholar]
- Arditti, J.; Ghani, A.K.A. Tansley Review No. 110. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [Green Version]
- Barsberg, S.; Lee, Y.-I.; Rasmussen, H.N. Development of C-lignin with G/S-lignin and lipids in orchid seed coats—An unexpected diversity exposed by ATR-FT-IR spectroscopy. Seed Sci. Res. 2018, 28, 41–51. [Google Scholar] [CrossRef]
- Şeker Şenay, S.; Şenel, G. Comparative seed micromorphology and morphometry of some orchid species (Orchidaceae) belong to the related Anacamptis, Orchis and Neotinea genera. Biologia 2017, 72, 14–23. [Google Scholar] [CrossRef]
- Diantina, S.; McGill, C.R.; Millner, J.; Nadarajan, J.; Pritchard, H.W.; McCormick, A.C. Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits. Plants 2020, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Miura, C.; Saisho, M.; Yagame, T.; Yamato, M.; Kaminaka, H. Bletilla striata (Orchidaceae) Seed Coat Restricts the Invasion of Fungal Hyphae at the Initial Stage of Fungal Colonization. Plants 2019, 8, 280. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.-L.; Chomicki, G.; Hao, K.; Liu, Q.; Xiong, Y.-Z.; Renner, S.S.; Gao, J.-Y.; Huang, S.-Q. Transitions between the Terrestrial and Epiphytic Habit Drove the Evolution of Seed-Aerodynamic Traits in Orchids. Am. Nat. 2019, 195, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.D.; Clark, J.Y.; Culham, A. The Cinderella discipline: Morphometrics and their use in botanical classification. Bot. J. Linn. Soc. 2020, 194, 385–396. [Google Scholar] [CrossRef]
- Arditti, J.; Michaud, J.D.; Healey, P.L. Morphometry of Orchid Seeds. II. Native California and Related Species of Calypso, Cephalanthera, Corallorhiza and Epipactis. Am. J. Bot. 1980, 67, 347–360. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Broeck, A.V.; Brys, R.; De Kort, H. Immigrant and extrinsic hybrid seed inviability contribute to reproductive isolation between forest and dune ecotypes of Epipactis helleborine (Orchidaceae). Oikos 2017, 127, 73–84. [Google Scholar] [CrossRef]
- Jacquemyn, H.; De Kort, H.; Broeck, A.V.; Brys, R. Low genetic divergence and variation in coastal dune populations of the widespread terrestrial orchid Epipactis helleborine. Bot. J. Linn. Soc. 2020, 193, 419–430. [Google Scholar] [CrossRef]
- Rewicz, A.; Kołodziejek, J.; Jakubska-Busse, A. The role of anthropogenic habitats as substitutes for natural habitats: A case study on Epipactis helleborine (L.) Crantz (Orchidaceae, Neottieae).Variations in size and nutrient composition of seeds. Turk. J. Bot. 2016, 40, 258–268. [Google Scholar] [CrossRef]
- Jakubska-Busse, A.; Żołubak, E.; Górniak, M.; Łobas, Z.; Tsiftsis, S.; Steiu, C. A Revision of the Taxonomy and Identification of Epipactis greuteri (Orchidaceae, Neottieae). Plants 2020, 9, 783. [Google Scholar] [CrossRef]
- Delforge, P. Epipactis. In Orchidées d’Europe, d’Afrique du Nord et du Påroche Orient, 4th ed.; Delachaux et Niestlé: Paris, France, 2016; pp. 45–46. [Google Scholar]
- De Agostini, A.; Caltagirone, C.; Caredda, A.; Cicatelli, A.; Cogoni, A.; Farci, D.; Guarino, F.; Garau, A.; Labra, M.; Lussu, M.; et al. Heavy metal tolerance of orchid populations growing on abandoned mine tailings: A case study in Sardinia Island (Italy). Ecotoxicol. Environ. Saf. 2020, 189, 110018. [Google Scholar] [CrossRef]
- Jurkiewicz, A.; Turnau, K.; Mesjasz-Przybyłowicz, J.; Przybyłowicz, W.; Godzik, B. Heavy metal localisation in mycorrhizas ofEpipactis atrorubens (Hoffm.) Besser (Orchidaceae) from zinc mine tailings. Protoplasma 2001, 218, 117–124. [Google Scholar] [CrossRef]
- Richards, A.J.; Swan, G.A. Epipactis leptochila (Godfery) Godfery and E. phyllanthes G. E. Srn. occurring in South Northumberland on lead and zinc soils. Watsonia 1976, 11, 1–5. [Google Scholar]
- Shefferson, R.P.; Kull, T.; Tali, K. Mycorrhizal interactions of orchids colonizing Estonian mine tailings hills. Am. J. Bot. 2008, 95, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Szarek-Łukaszewska, G. Vegetation of reclaimed and spontaneously vegetated Zn-Pb mine wastes in Southern Poland. Pol. J. Environ. 2009, 18, 717–733. [Google Scholar]
- Howard-Williams, C. Morphological Variation between Isolated populations of Becium homblei (De Wild) Duvign & Plancke growing on heavy metal soils. Vegetatio 1971, 23, 141–151. [Google Scholar]
- Luzuriaga, A.L.; Escudero, A.; Perez-Garcia, F. Environmental maternal effects on seed morphology and germination in Sinapis arvensis (Cruciferae). Weed Res. 2006, 46, 163–174. [Google Scholar] [CrossRef]
- Farci, D.; Haniewicz, P.; Cocco, E.; De Agostini, A.; Cortis, P.; Kusaka, M.; Loi, M.C.; Piano, D. The Impact of Fruit Etiolation on Quality of Seeds in Tobacco. Front. Plant Sci. 2020, 11, 563971. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Lan, H.; Zhang, F. Variation of seed heteromorphism in Chenopodium album and the effect of salinity stress on the descendants. Ann. Bot. 2010, 105, 1015–1025. [Google Scholar] [CrossRef]
- Bradley, R.; Burt, A.J.; Read, D.J. The biology of mycorrhiza in the Ericaceae. VIII. The role of mycorrhizal infection in heavy metal resistance. New Phytol. 1982, 91, 197–209. [Google Scholar] [CrossRef]
- Gonzalez-Chavez, C.; Harris, P.J.; Dodd, J.; Meharg, A.A. Arbuscular mycorrhizal fungi confer enhanced arsenate resistance on Holcus lanatus. New Phytol. 2002, 155, 163–171. [Google Scholar] [CrossRef]
- González-Guerrero, M.; Melville, L.H.; Ferrol, N.; Lott, J.N.; Azcón-Aguilar, C.; Peterson, R.L. Ultrastructural localization of heavy metals in the extraradical mycelium and spores of the arbuscular mycorrhizal fungus Glomus intraradices. Can. J. Microbiol. 2008, 54, 103–110. [Google Scholar] [CrossRef]
- Joner, E.J.; Briones, R.; Leyval, C. Metal-binding capacity of arbuscular mycorrhizal mycelium. Plant. Soil 2000, 226, 227–234. [Google Scholar] [CrossRef]
- Martino, E.; Turnau, K.; Girlanda, M.; Bonfante, P.; Perotto, S. Ericoid mycorrhizal fungi from heavy metal polluted soils: Their identification and growth in the presence of zinc ions. Mycol. Res. 2000, 104, 338–344. [Google Scholar] [CrossRef]
- Perotto, S.; Girlanda, M.; Martino, E. Ericoid mycorrhizal fungi: Some new perspectives on old acquaintances. Plant. Soil 2002, 244, 41–53. [Google Scholar] [CrossRef]
- Turnau, K.; Dexheimer, J. Acid phosphatase activity in Pisolithus arrhizus mycelium treated with cadmium dust. Mycorrhiza 1995, 5, 205–211. [Google Scholar] [CrossRef]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Gadd, G.M.; De Rome, L. Biosorption of copper by fungal melanin. Appl. Microbiol. Biotechnol. 1988, 29, 610–617. [Google Scholar] [CrossRef]
- Kumar, M.S.J.; Singh, M.; Singh, S.; Singh, V.P.; Prasad, S.M.; Singh, M. Adaptation Strategies of Plants against Heavy Metal Toxicity: A Short Review. Biochem. Pharmacol. Open Access 2015, 4, 161. [Google Scholar] [CrossRef] [Green Version]
- Chaoui, A.; El Ferjani, E. β-Estradiol Protects Embryo Growth from Heavy-Metal Toxicity in Germinating Lentil Seeds. J. Plant Growth Regul. 2013, 32, 636–645. [Google Scholar] [CrossRef]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Li, W.; Khan, M.A.; Yamaguchi, S.; Kamiya, Y. Effects of heavy metals on seed germination and early seedling growth of Arabidopsis thaliana. Plant Growth Regul. 2005, 46, 45–50. [Google Scholar] [CrossRef]
- Monika, K.; Romana, I. Ovules, Female Gametophytes and Embryos are More Sensitive to Heavy Metal Pollution than Anthers and Pollen of Cardaminopsis Arenosa (L.) Hayek (Brassicaceae), A Member of Calamine Flora. Acta Biol. Cracoviensia Ser. Bot. 2014, 56, 128–137. [Google Scholar] [CrossRef]
- Yang, J.; Cai, L.; Liu, D.; Chen, G.; Gratzfeld, J.; Sun, W. China’s conservation program on Plant Species with Extremely Small Populations (PSESP): Progress and perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- Sulis, E.; Bacchetta, G.; Cogoni, D.; Gargano, D.; Fenu, G. Assessing the global conservation status of the rock rose Helianthemum caput-felis. Oryx 2019, 54, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Fenu, G.; Bacchetta, G.; Christodoulou, C.S.; Cogoni, D.; Fournaraki, C.; Pietro, G.D.G.G.; Gotsiou, P.; Kyratzis, A.; Piazza, C.; Vicens, M.; et al. A Common Approach to the Conservation of Threatened Island Vascular Plants: First Results in the Mediterranean Basin. Diversity 2020, 12, 157. [Google Scholar] [CrossRef] [Green Version]
- Lai, R. Aggiornamento Corologico, Tassonomico, Nomenclaturale Della Flora Orchidologica Della Sardegna. Ph.D. Thesis, University of Cagliari, Cagliari, Italy, 2008. [Google Scholar]
- Lussu, M.; Marignani, M.; Lai, R.; Loi, M.C.; Cogoni, A.; Cortis, P. A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids? Plants 2020, 9, 853. [Google Scholar] [CrossRef]
- De Waele, J.; Pisano, M. Interazione fra attività mineraria ed un acquifero carsico: L’esempio di Barraxiutta (Sardegna sud-occidentale). In Proceedings of the Convegno Nazionale Sull’Inquinamento Delle Grotte e Degli Acquiferi Carsici e Possibili Ricadute Sulla Collettività, Ponte di Brenta, Italy, 26–27 September 1998; pp. 195–209. [Google Scholar]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 14 September 2020).
- Kassambara, A. Ggpubr: ‘Ggplot2’ Based Publication Ready Plots. R Package Version 0.2.4. 2019. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 20 November 2020).
- Peterson, B.G.; Carl, P. PerformanceAnalytics: Econometric Tools for Performance and Risk Analysis, R Package Version 2.0.4. 2020. Available online: https://CRAN.R-project.org/package=PerformanceAnalytics (accessed on 20 November 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | PPS | PCS | Test | Test Results |
|---|---|---|---|---|
| Coat length (mm) | 1.13 ± 0.02 | 0.94 ± 0.02 | Mann–Whitney U test | W = 8846; p = 1.06 × 10−5 **** |
| Embryo length (mm) | 0.24 ± 0.002 | 0.23 ± 0.004 | Mann–Whitney U test | W = 6281.5; p = 0.13, ns |
| Coat width (mm) | 0.27 ± 0.004 | 0.25 ± 0.004 | Mann–Whitney U test | W = 7204.5; p = 3.61 × 10−4 ** |
| Embryo width (mm) | 0.13 ± 0.002 | 0.14 ± 0.003 | Mann–Whitney U test | W = 3904.5; p = 1.21 × 10−4 *** |
| Coat area (mm2) | 0.24 ± 0.005 | 0.19 ± 0.006 | t-test | t = 6.07; p = 5.79 × 10−4 **** |
| Embryo area (mm2) | 0.02 ± 0.001 | 0.03 ± 0.001 | t-test | t = -1.83; p = 0.07, ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Agostini, A.; Cortis, P.; Cogoni, A.; Gargiulo, R.; Fenu, G. Epipactis tremolsii Seed Diversity in Two Close but Extremely Different Populations: Just a Case of Intraspecific Variability? Plants 2020, 9, 1625. https://doi.org/10.3390/plants9111625
De Agostini A, Cortis P, Cogoni A, Gargiulo R, Fenu G. Epipactis tremolsii Seed Diversity in Two Close but Extremely Different Populations: Just a Case of Intraspecific Variability? Plants. 2020; 9(11):1625. https://doi.org/10.3390/plants9111625
Chicago/Turabian StyleDe Agostini, Antonio, Pierluigi Cortis, Annalena Cogoni, Roberta Gargiulo, and Giuseppe Fenu. 2020. "Epipactis tremolsii Seed Diversity in Two Close but Extremely Different Populations: Just a Case of Intraspecific Variability?" Plants 9, no. 11: 1625. https://doi.org/10.3390/plants9111625