In-silico Exploration of Channel Type and Efflux Silicon Transporters and Silicification Proteins in 80 Sequenced Viridiplantae Genomes

 ,
,  , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Data Retrieval
2.2. Sequence Analyses
2.3. Phylogenetic Analyses
2.4. Prediction of Silica Precipitation Potential of Selected Peptide Sequences
2.5. Digital Expression and Co-Expression Network Analysis
3. Results
3.1. Identification of SITs and Slps
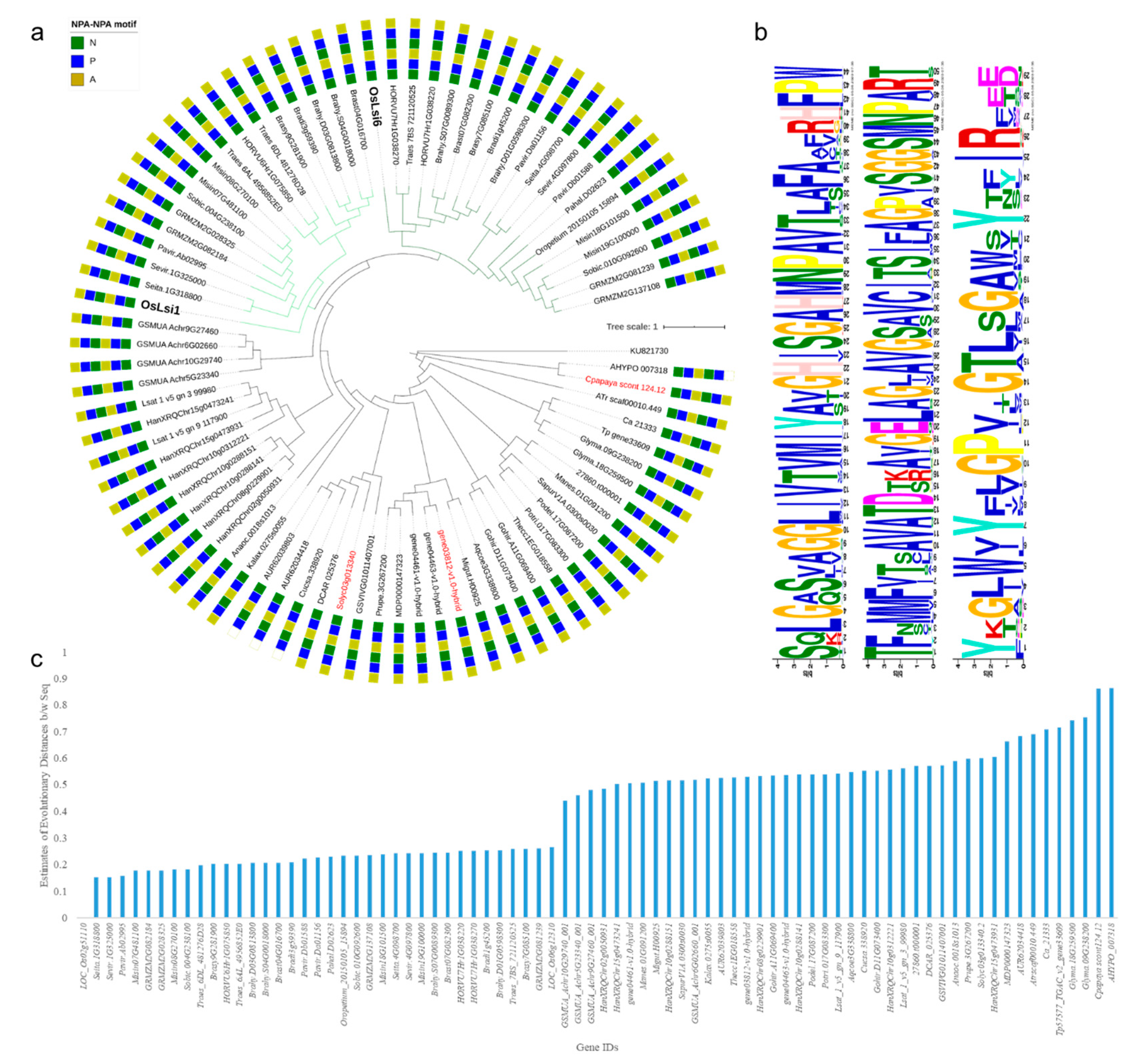
3.1.1. Channel Type SITs in Viridiplantae
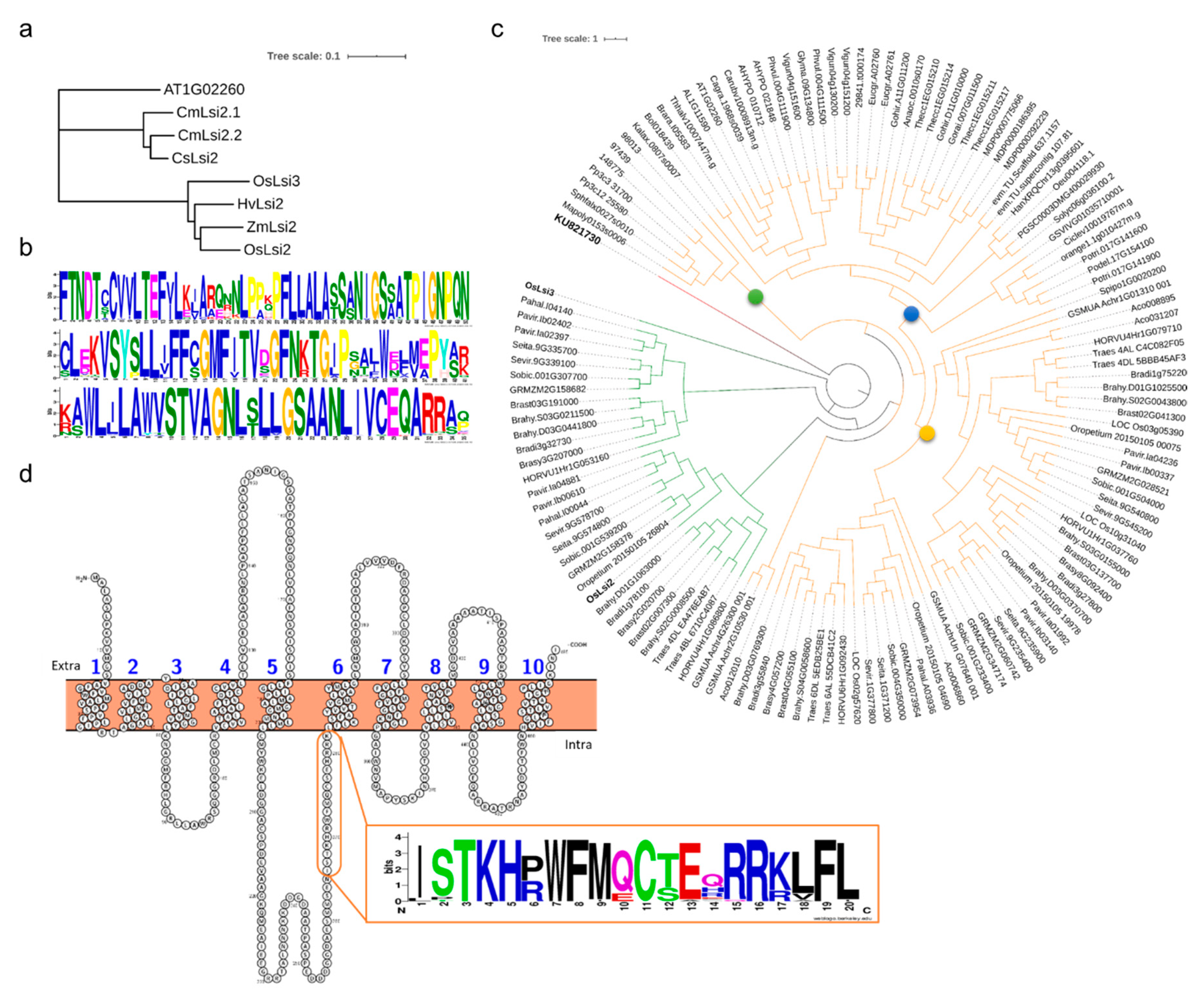
3.1.2. Lsi2s in Viridiplantae
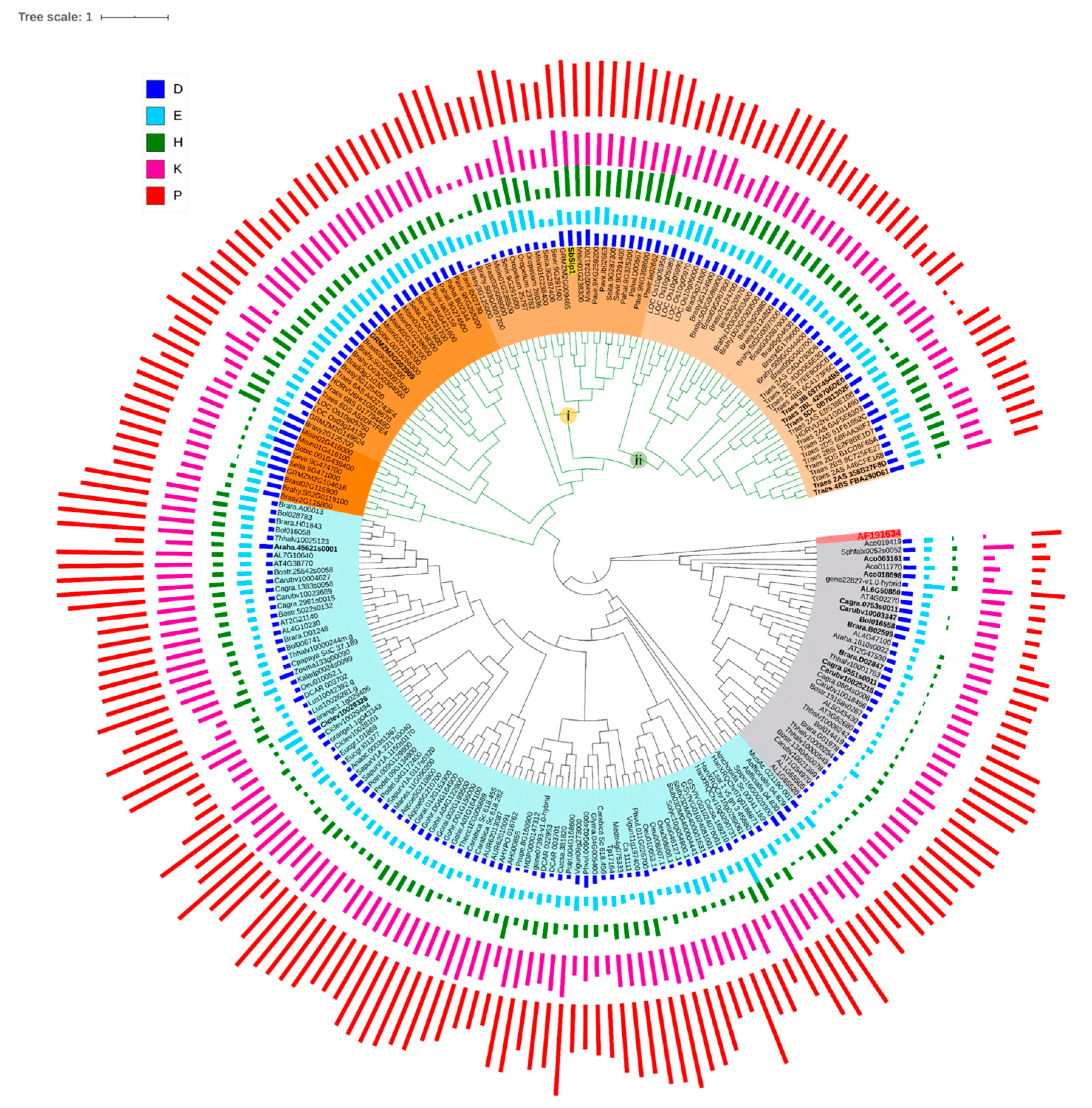
3.1.3. Siliplant1 Homologs in Viridiplantae
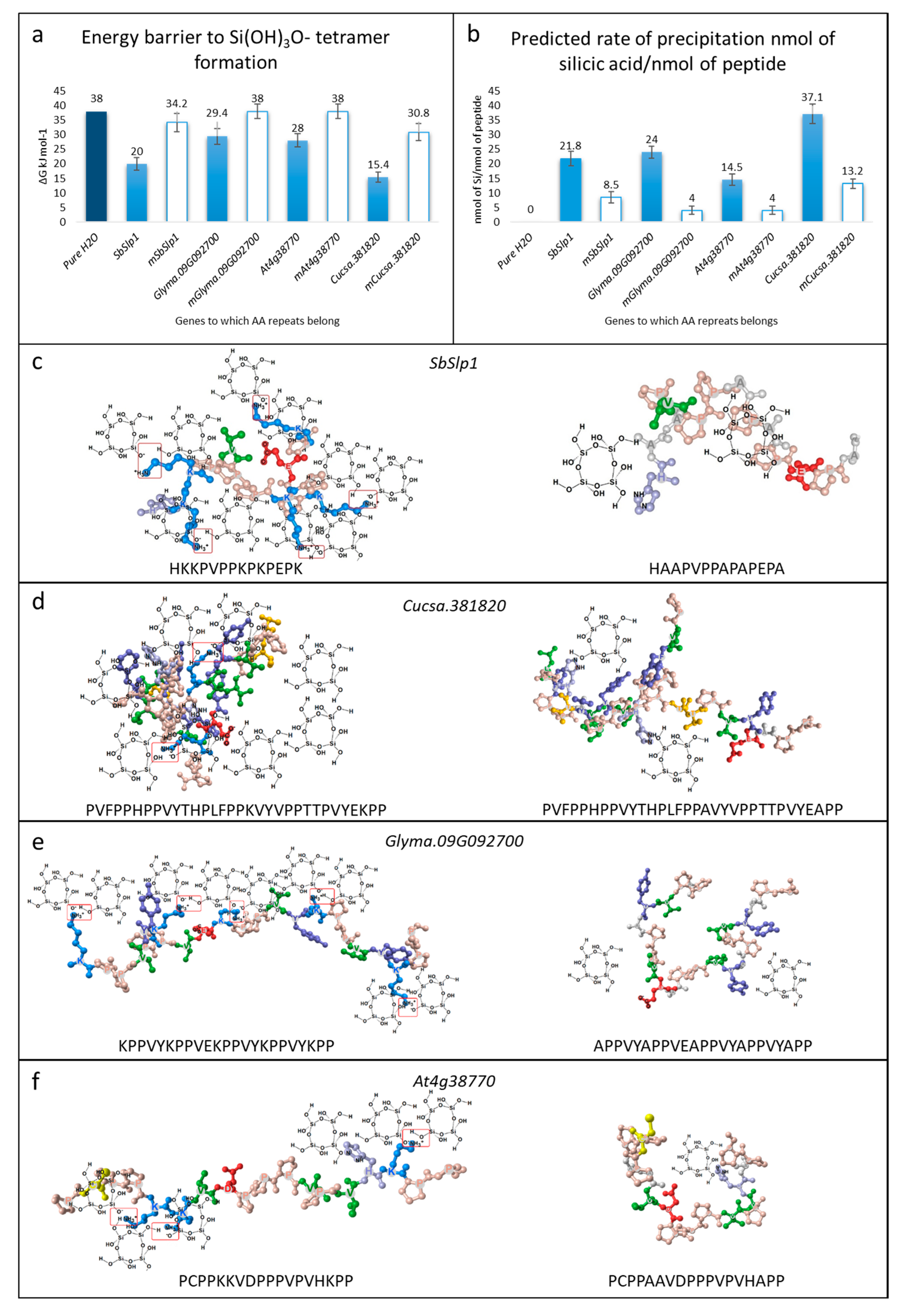
3.2. Silica Precipitation Potential of Repeats
3.3. Digital Expression of SITs and Slps
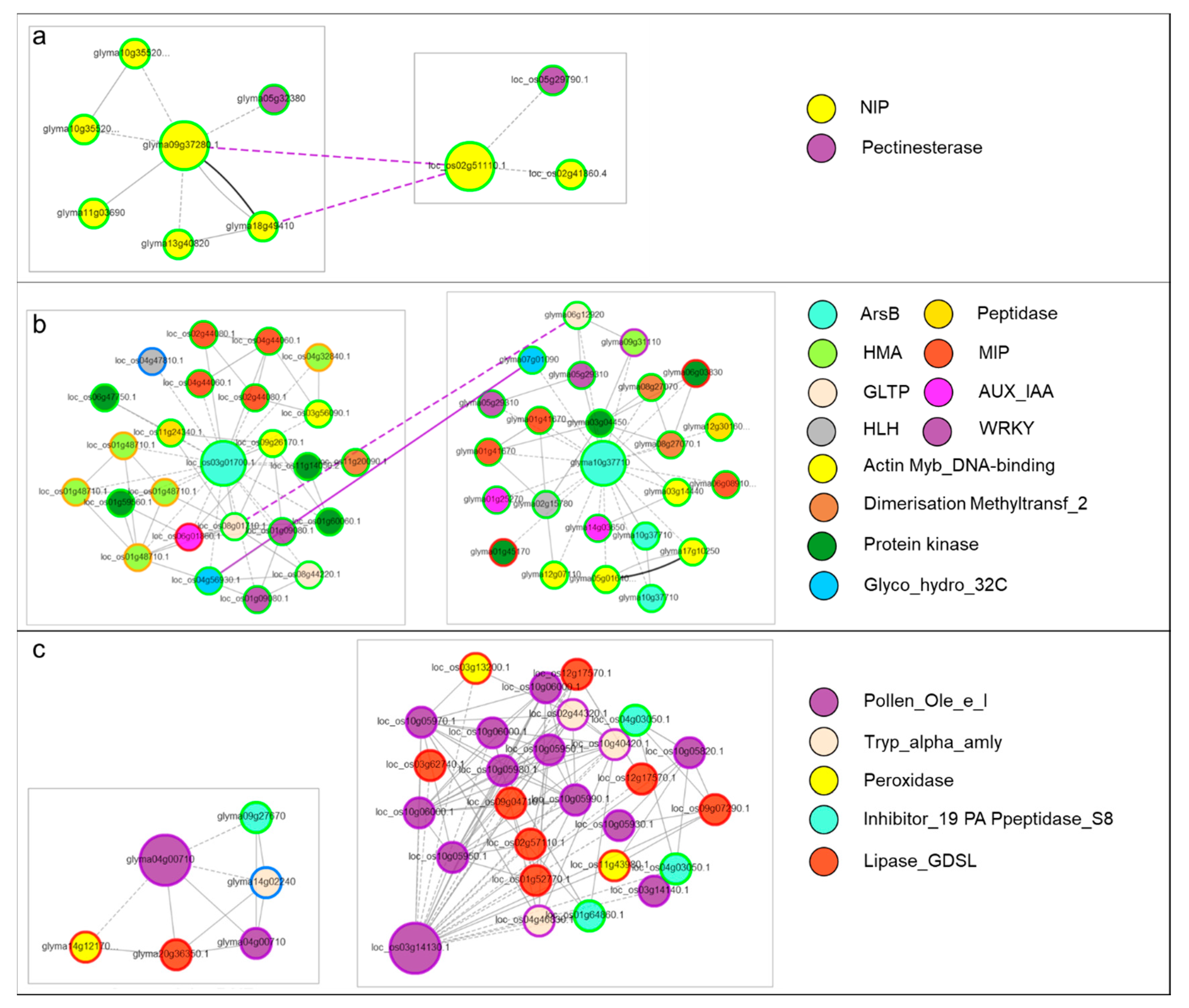
3.4. Co-Expression Networks of Si Transporters and Slps
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Deshmukh, R.; Sonah, H.; Belanger, R. New evidence defining the evolutionary path of aquaporins regulating silicon uptake in land plants. J. Exp. Bot. 2020, eraa342. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Zakharenko, A.M.; Zemchenko, I.V.; Haider, M.S.; Ali, M.A.; Imtiaz, M.; Chung, G.; Tsatsakis, A.; Sun, S.; Golokhvast, K.S. Phytolith Formation in Plants: From Soil to Cell. Plants 2019, 8, 249. [Google Scholar]
- Haynes, R.J. A contemporary overview of silicon availability in agricultural soils. J. Plant Nutr. Soil Sci. 2014, 177, 831–844. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [PubMed]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688. [Google Scholar] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Mitani-Ueno, N.; Yamaji, N.; Zhao, F.J.; Ma, J.F. The aromatic/arginine selectivity filter of NIP aquaporins plays a critical role in substrate selectivity for silicon, boron, and arsenic. J. Exp. Bot. 2011, 62, 4391–4398. [Google Scholar] [CrossRef] [Green Version]
- Gregoire, C.; Remus-Borel, W.; Vivancos, J.; Labbe, C.; Belzile, F.; Belanger, R.R. Discovery of a multigene family of aquaporin silicon transporters in the primitive plant Equisetum arvense. Plant J. 2012, 72, 320–330. [Google Scholar] [CrossRef]
- Sun, H.; Duan, Y.; Mitani-Ueno, N.; Che, J.; Jia, J.; Liu, J.; Guo, J.; Ma, J.F.; Gong, H. Tomato roots have a functional silicon influx transporter but not a functional silicon efflux transporter. Plant Cell Environ. 2020, 43, 732–744. [Google Scholar]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guerin, V.; Carpentier, G.; Sonah, H.; Labbe, C.; Isenring, P.; Belzile, F.J.; Belanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E.; Gozukara, N. Genome-wide exploration of silicon (Si) transporter genes, Lsi1 and Lsi2 in plants; insights into Si-accumulation status/capacity of plants. BioMetals 2017, 30, 185–200. [Google Scholar] [PubMed]
- Kumar, S.; Soukup, M.; Elbaum, R. Silicification in grasses: Variation between different cell types. Front. Plant Sci. 2017, 8, 438. [Google Scholar] [PubMed] [Green Version]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Altamira Press: Lanham, MD, USA, 2006. [Google Scholar]
- Exley, C. A possible mechanism of biological silicification in plants. Front. Plant Sci. 2015, 6, 853. [Google Scholar] [PubMed] [Green Version]
- Guerriero, G.; Hausman, J.F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Science 2016, 7, 463. [Google Scholar]
- Marron, A.O.; Ratcliffe, S.; Wheeler, G.L.; Goldstein, R.E.; King, N.; Not, F.; de Vargas, C.; Richter, D.J. The evolution of silicon transport in eukaryotes. Mol. Biol. Evol. 2016, 33, 3226–3248. [Google Scholar] [CrossRef] [Green Version]
- Pommerrenig, B.; Diehn, T.A.; Bernhardt, N.; Bienert, M.D.; Mitani-Ueno, N.; Fuge, J.; Bieber, A.; Spitzer, C.; Bräutigam, A.; Ma, J.F. Functional evolution of nodulin 26-like intrinsic proteins: From bacterial arsenic detoxification to plant nutrient transport. New Phytol. 2020, 225, 1383–1396. [Google Scholar]
- Trembath-Reichert, E.; Wilson, J.P.; McGlynn, S.E.; Fischer, W.W. Four hundred million years of silica biomineralization in land plants. Proc. Natl. Acad. Sci. USA 2015, 112, 5449–5454. [Google Scholar]
- Kumar, S.; Adiram-Filiba, N.; Blum, S.; Sanchez-Lopez, J.A.; Tzfadia, O.; Omid, A.; Volpin, H.; Heifetz, Y.; Goobes, G.; Elbaum, R. Siliplant1 (Slp1) protein precipitates silica in sorghum silica cells. J. Exp. Bot. 2019, eraa258. [Google Scholar] [CrossRef]
- Raven, J.; Waite, A. The evolution of silicification in diatoms: Inescapable sinking and sinking as escape? New Phytol. 2004, 162, 45–61. [Google Scholar]
- Otzen, D. The role of proteins in biosilicification. Scientifica 2012, 2012. [Google Scholar] [CrossRef]
- Pamirsky, I.E.; Golokhvast, K.S. Origin and status of homologous proteins of biomineralization (Biosilicification) in the taxonomy of phylogenetic domains. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Kröger, N.; Deutzmann, R.; Sumper, M. Polycationic peptides from diatom biosilica that direct silica nanosphere formation. Science 1999, 286, 1129–1132. [Google Scholar] [PubMed] [Green Version]
- Kotzsch, A.; Pawolski, D.; Milentyev, A.; Shevchenko, A.; Scheffel, A.; Poulsen, N.; Shevchenko, A.; Kröger, N. Biochemical composition and assembly of biosilica-associated insoluble organic matrices from the diatom Thalassiosira pseudonana. J. Biol. Chem. 2016, 291, 4982–4997. [Google Scholar] [PubMed] [Green Version]
- Kauss, H.; Seehaus, K.; Franke, R.; Gilbert, S.; Dietrich, R.A.; Kröger, N. Silica deposition by a strongly cationic proline-rich protein from systemically resistant cucumber plants. Plant J. 2003, 33, 87–95. [Google Scholar]
- Kersey, P.J. Plant genome sequences: Past, present, future. Curr. Opin. Plant Biol. 2019, 48, 1–8. [Google Scholar]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953. [Google Scholar]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.; Potter, S.C.; Finn, R.D. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nature Biotechnol. 2019, 37, 420–423. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587. [Google Scholar]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2006, 23, 127–128. [Google Scholar]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H., Eds.; Academic Press: New York, NY, USA, 1965. [Google Scholar]
- Leaver-Fay, A.; Tyka, M.; Lewis, S.M.; Lange, O.F.; Thompson, J.; Jacak, R.; Kaufman, K.W.; Renfrew, P.D.; Smith, C.A.; Sheffler, W. ROSETTA3: An object-oriented software suite for the simulation and design of macromolecules. Methods Enzymol. 2011, 487, 545–574. [Google Scholar]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar]
- Waese, J.; Provart, N.J. The bio-analytic resource: Data visualization and analytic tools for multiple levels of plant biology. Curr. Plant Biol. 2016, 7, 2–5. [Google Scholar]
- Waese, J.; Fan, J.; Pasha, A.; Yu, H.; Fucile, G.; Shi, R.; Cumming, M.; Kelley, L.A.; Sternberg, M.J.; Krishnakumar, V. ePlant: Visualizing and exploring multiple levels of data for hypothesis generation in plant biology. Plant Cell 2017, 29, 1806–1821. [Google Scholar]
- Mutwil, M.; Klie, S.; Tohge, T.; Giorgi, F.M.; Wilkins, O.; Campbell, M.M.; Fernie, A.R.; Usadel, B.; Nikoloski, Z.; Persson, S. PlaNet: Combined sequence and expression comparisons across plant networks derived from seven species. Plant Cell 2011, 23, 895–910. [Google Scholar] [PubMed] [Green Version]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Silicon efflux transporters isolated from two pumpkin cultivars contrasting in Si uptake. Plant Signal. Behav. 2011, 6, 991–994. [Google Scholar] [PubMed] [Green Version]
- Vivancos, J.; Deshmukh, R.; Gregoire, C.; Remus-Borel, W.; Belzile, F.; Belanger, R.R. Identification and characterization of silicon efflux transporters in horsetail (Equisetum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [PubMed] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar]
- Chen, M.; Xu, J.; Devis, D.; Shi, J.; Ren, K.; Searle, I.; Zhang, D. Origin and functional prediction of pollen allergens in plants. Plant Physiol. 2016, 172, 341–357. [Google Scholar]
- Santino, C.G.; Stanford, G.L.; Conner, T.W. Developmental and transgenic analysis of two tomato fruit enhanced genes. Plant Mol. Biol. 1997, 33, 405–416. [Google Scholar]
- Hodson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V. Silicification in the microalgae. In The Physiology of Microalgae; Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer: Cham, Switzerland, 2016; pp. 289–300. [Google Scholar]
- Epstein, E. Silicon. Annu. Rev. Plant Biol. 1999, 50, 641–664. [Google Scholar]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. (Eds.) Silicon in Agriculture; Springer: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Becker, B. Snow ball earth and the split of Streptophyta and Chlorophyta. Trends Plant Sci. 2013, 18, 180–183. [Google Scholar]
- Ma, J.F. Silicon transporters in higher plants. In MIPs and Their Role in the Exchange of Metalloids; Jahn, T.P., Bienert, G.P., Eds.; Springer: New York, NY, USA, 2010; pp. 99–109. [Google Scholar]
- Zellner, W.; Lutz, L.; Khandekar, S.; Leisner, S. Identification of NtNIP2; 1: An Lsi1 silicon transporter in Nicotiana tabacum. J. Plant Nutr. 2019, 42, 1028–1035. [Google Scholar]
- Deshmukh, R.K.; Vivancos, J.; Guerin, V.; Sonah, H.; Labbe, C.; Belzile, F.; Belanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Boldt, J.K.; Locke, J.C.; Altland, J.E. Silicon accumulation and distribution in petunia and sunflower grown in a rice hull-amended substrate. HortScience 2018, 53, 698–703. [Google Scholar]
- Bienert, G.P.; Schüssler, M.D.; Jahn, T.P. Metalloids: Essential, beneficial or toxic? Major intrinsic proteins sort it out. Trends Biochem. Sci. 2008, 33, 20–26. [Google Scholar]
- Benvenuto, M.L.; Osterrieth, M.L. Silicophytoliths from soybean plants in different growth stages of the Argentine Pampas. Braz. J. Bot. 2016, 39, 337–347. [Google Scholar]
- Zellner, W.; Lawrence, D. Silicon as a biostimulant in agriculture. Burleigh Dodds Ser. Agric. Sci. 2020, 1, 1–38. [Google Scholar]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of maize silicon influx transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Raiola, A.; Lionetti, V.; Elmaghraby, I.; Immerzeel, P.; Mellerowicz, E.J.; Salvi, G.; Cervone, F.; Bellincampi, D. Pectin methylesterase is induced in Arabidopsis upon infection and is necessary for a successful colonization by necrotrophic pathogens. Mol. Plant Microbe Interact. 2011, 24, 432–440. [Google Scholar]
- He, C.; Wang, L.; Liu, J.; Liu, X.; Li, X.; Ma, J.; Lin, Y.; Xu, F. Evidence for ‘silicon’within the cell walls of suspension-cultured rice cells. New Phytol. 2013, 200, 700–709. [Google Scholar]
- Głazowska, S.; Baldwin, L.; Mravec, J.; Bukh, C.; Hansen, T.H.; Jensen, M.M.; Fangel, J.U.; Willats, W.G.; Glasius, M.; Felby, C. The impact of silicon on cell wall composition and enzymatic saccharification of Brachypodium distachyon. Biotechnol. Biofuels 2018, 11, 171. [Google Scholar]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the wall—Sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants 2018, 7, 89. [Google Scholar]
- Rui, Y.; Dinneny, J.R. A wall with integrity: Surveillance and maintenance of the plant cell wall under stress. New Phytol. 2020, 225, 1428–1439. [Google Scholar] [PubMed] [Green Version]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant–pathogen interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [PubMed] [Green Version]
- Ali, M.A.; Azeem, F.; Nawaz, M.A.; Acet, T.; Abbas, A.; Imran, Q.M.; Shah, K.H.; Rehman, H.M.; Chung, G.; Yang, S.H. Transcription factors WRKY11 and WRKY17 are involved in abiotic stress responses in Arabidopsis. J. Plant Physiol. 2018, 226, 12–21. [Google Scholar]
- Feng, J.; Shi, Q.; Wang, X.; Wei, M.; Yang, F.; Xu, H. Silicon supplementation ameliorated the inhibition of photosynthesis and nitrate metabolism by cadmium (Cd) toxicity in Cucumis sativus L. Sci. Hortic. 2010, 123, 521–530. [Google Scholar]
- Jia-Wen, W.; Yu, S.; Yong-Xing, Z.; Yi-Chao, W.; Hai-Jun, G. Mechanisms of enhanced heavy metal tolerance in plants by silicon: A review. Pedosphere 2013, 23, 815–825. [Google Scholar]
- Kim, Y.H.; Khan, A.L.; Kim, D.-H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biology 2014, 14, 1–13. [Google Scholar]
- Songnuan, W. Wind-pollination and the roles of pollen allergenic proteins. Asian Pac. J. Allergy Immunol. 2013, 31, 261. [Google Scholar]
- Brugiére, T.; Exley, C. Callose-associated silica deposition in Arabidopsis. J. Trace Elem. Med. Biol. 2017, 39, 86–90. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genome Version | Species | Genome Version |
|---|---|---|---|
| Amaranthus hypochondriacus | V2.1 | Kalanchoe laxiflora | V1.1 |
| Amborella trichopoda | V1.0 | Lactuca sativa | V8 |
| Anacardium occidentale | V0.9 | Linum usitatissimum | V1.0 |
| Ananas comosus | V3 | Malus domestica | V1.0 |
| Aquilegia coerulea | V3.1 | Manihot esculenta | V6.1 |
| Arabidopsis halleri | V1.1 | Marchantia polymorpha | V3.1 |
| Arabidopsis lyrata | V2.1 | Medicago truncatula | 4.0V1 |
| Arabidopsis thaliana | TAIR10 | Micromonas pusilla CCMP1545 | V3.0 |
| Asparagus officinalis | V1.1 | Mimulus guttatus | V2.0 |
| Boechera stricta | V1.2 | Miscanthus sinensis | V7.1 |
| Botryococcus braunii | V2.1 | Musa acuminata | V1 |
| Brachypodium distachyon | V3.1 | Olea europaea Var. sylVestris | V1.0 |
| Brachypodium hybridum | V1.1 | Oropetium thomaeum | V1.0 |
| Brachypodium stacei | V1.1 | Oryza sativa | V7_JGI |
| Brachypodium sylVaticum | V1.1 | Ostreococcus lucimarinus | V2.0 |
| Brassica oleracea capitata | V1.0 | Panicum hallii | V3.1 |
| Brassica rapa FPsc | V1.3 | Panicum Virgatum | V4.1 |
| Capsella grandiflora | V1.1 | Phaseolus Vulgaris | V2.1 |
| Capsella rubella | V1.0 | Physcomitrella patens | V3.3 |
| Carica papaya | V0.4 | Populus deltoides WV94 | V2.1 |
| Chenopodium quinoa | V1.0 | Populus trichocarpa | V3.1 |
| Chlamydomonas reinhardtii | V5.5 | Prunus persica | V2.1 |
| Chromochloris zofingiensis | V5.2.3.2 | Ricinus communis | V0.1 |
| Cicer arietinum | V1.0 | Salix purpurea | V1.0 |
| Citrus clementina | V1.0 | Selaginella moellendorffii | V1.0 |
| Citrus sinensis | V1.1 | Setaria italica | V2.2 |
| Coccomyxa subellipsoidea C-169 | V2.0 | Setaria Viridis | V2.1 |
| Coffea arabica | UCDV0.5 | Solanum lycopersicum iTAG | 2.4 |
| Cucumis sativus | V1.0 | Solanum tuberosum | V4.03 |
| Daucus carota | V2.0 | Sorghum bicolor | V3.1.1 |
| Dunaliella salina | V1.0 | Sphagnum fallax | V0.5 |
| Eucalyptus grandis | V2.0 | Spirodela polyrhiza | V2 |
| Eutrema salsugineum | V1.0 | Theobroma cacao | V1.1 |
| Fragaria Vesca | V1.1 | Trifolium pratense | V2 |
| Glycine max Wm82 | a2.V1 | Triticum aestiVum | V2.2 |
| Gossypium hirsutum | V1.1 | Vigna unguiculata | V1.1 |
| Gossypium raimondii | V2.1 | Vitis Vinifera | 12X |
| Helianthus annuus | r1.2 | VolVox carteri | V2.1 |
| Hordeum Vulgare | r1 | Zea mays | Ensembl-18 |
| Kalanchoe fedtschenkoi | V1.1 | Zostera marina | V2.2 |
| Class | Order | Family | Species | Lsi1 | Lsi2 | Slp1 |
|---|---|---|---|---|---|---|
| Trebouxiophyceae | Trebouxiophyceae | Coccomyxaceae | Coccomyxa subellipsoidea | |||
| Trebouxiales | Botryococcaceae | Botryococcus braunii | ||||
| Mamiellophyceae | Mamiellales | Mamiellaceae | Micromonas PUSILLA ccmp1545 | 1 | ||
| Bathycoccaceae | Osteococcus lucimarinus | 1 | ||||
| Chlorophyceae | Sphaeropleales | Chromochloridaceae | Chromochloris zofingiensis | 2 | ||
| Chlamydomonadales | Volvocaceae | Volvox carteri | 2 | |||
| Chlamydomonadaceae | Chlamydomonas reinhardtii | 1 | ||||
| Dunaliellaceae | Dunaliella salina | 1 | ||||
| Lycopodiopsida | Selaginellales | Selaginellaceae | Selaginella moellendorffii | 3 | ||
| Sphagnopsida | Sphagnales | Sphagnaceae | Sphagnum fallax | 1 | 1 | |
| Bryopsida | Funariales | Funariaceae | Physcomitrella patens | 2 | ||
| Marchantiopsida | Marchantiales | Marchantiaceae | Marchantia polymorpha | 1 | ||
| Liliopsida | Alismatales | Araceae | Spirodela polyrhiza | 1 | 1 | |
| Zosteraceae | Zostera marina | 1 | ||||
| Asparagales | Asparagaceae | Asparagus officinalis | 1 | 2 | ||
| Zingiberales | Musaceae | Musa acuminata | 4 | 4 | 1 | |
| Poales | Bromeliaceae | Ananas comosus | 4 | 4 | ||
| Poaceae | Oropetium thomaeum | 1 | 4 | 3 | ||
| Panicum halli | 1 | 3 | 4 | |||
| Panicum virgatum | 3 | 8 | 8 | |||
| Brachypodium stacei | 2 | 5 | 6 | |||
| Brachypodium hybridum | 4 | 10 | 11 | |||
| Brachypodium sylvaticum | 2 | 4 | 8 | |||
| Brachypodium distachyon | 2 | 5 | 5 | |||
| Zea mays | 4 | 5 | 4 | |||
| Triticum aestivum | 3 | 6 | 20 | |||
| Sorghum bicolor | 2 | 5 | 3 | |||
| Setaria viridis | 2 | 5 | 5 | |||
| Setaria italica | 2 | 5 | 4 | |||
| Oryza sativa | 2 | 4 | 7 | |||
| Hordeum vulgare | 3 | 5 | 2 | |||
| Miscanthus sinensis | 4 | 10 | ||||
| Magnoliopsida | Ranunculales | Ranunculaceae | Aquilegia coerulea | 1 | 1 | 2 |
| Myrtales | Myrtaceae | Eucalyptus grandis | 2 | 2 | ||
| Saxifragales | Crassulaceae | Kalanchoe laxiflora | 1 | 2 | ||
| Kalanchoe fedtschenkoi | 1 | 1 | ||||
| Vitales | Vitaceae | Vitis vinifera | 1 | 1 | 2 | |
| Caryophyllales | Chenopodiaceae | Chenopodium quinoa | 2 | 2 | 2 | |
| Amaranthaceae | Amaranthus hypochondriacus | 1 | 2 | 2 | ||
| Gentianales | Rubiaceae | Coffea arabica | 1 | 3 | ||
| Amborellales | Amborellaceae | Amborella trichopoda | 1 | 4 | 1 | |
| Asterales | Asteraceae | Lactuca sativa | 2 | 1 | 1 | |
| Helianthus annuus | 7 | 6 | 3 | |||
| Lamiales | Lamiaceae | Capsella grandiflora | 1 | 5 | ||
| Phrymaceae | Mimulus guttatus | 1 | 1 | |||
| Oleaceae | Olea europaea | 1 | 5 | |||
| Solanales | Solanaceae | Solanum tuberosum | 1 | 2 | ||
| Solanum lycopersicum | 1 | 1 | 1 | |||
| Apiales | Apiaceae | Daucus carota | 1 | 1 | 3 | |
| Fabales | Fabaceae | Trifolium pratense | 1 | 3 | 1 | |
| Vigna unguiculata | 3 | 2 | ||||
| Phaseolus vulgaris | 2 | 2 | ||||
| Medicago truncatula | 2 | 1 | ||||
| Glycine max | 2 | 1 | 1 | |||
| Cicer arietinum | 1 | 1 | ||||
| Rosales | Rosaceae | Fragaria vesca | 3 | 2 | 2 | |
| Prunus persica | 1 | 1 | 1 | |||
| Malus domestica | 1 | 2 | 1 | |||
| Malpighiales | Linaceae | Linum usitatissimum | 2 | 2 | ||
| Euphorbiaceae | Ricinum communis | 1 | 1 | |||
| Mahihot esculenta | 1 | 1 | 1 | |||
| Salicaceae | Salix purpurea | 1 | 2 | 3 | ||
| Populus deltoides | 1 | 1 | 2 | |||
| Populus trichocarpa | 1 | 2 | 2 | |||
| Cucurbitales | Cucurbitaceae | Cucumis sativus | 1 | 1 | 2 | |
| Brassicales | Brassicaceae | Capsella rubella | 1 | 6 | ||
| Eutrema salugineum | 1 | 6 | ||||
| Boechera stricta | 1 | 4 | ||||
| Brassica oleracea | 1 | 5 | ||||
| Brassica rapa | 1 | 6 | ||||
| Arabidopsis halleri | 1 | 2 | ||||
| Arabidopsis lyrata | 1 | 7 | ||||
| Arabidopsis thaliana | 1 | 6 | ||||
| Caricaceae | Carica papaya | 1 | 1 | 1 | ||
| Malvales | Malvaceae | Theobroma cacao | 1 | 4 | 1 | |
| Gossypium raimondii | 3 | 2 | ||||
| Gossypium hirsutum | 2 | 3 | 4 | |||
| Sapindales | Anacardiacea | Anacardium occidentale | 1 | 1 | ||
| Rutaceae | Citrus clementia | 1 | 3 | |||
| Citrus sinensis | 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, M.A.; Azeem, F.; Zakharenko, A.M.; Lin, X.; Atif, R.M.; Baloch, F.S.; Chan, T.-F.; Chung, G.; Ham, J.; Sun, S.; et al. In-silico Exploration of Channel Type and Efflux Silicon Transporters and Silicification Proteins in 80 Sequenced Viridiplantae Genomes. Plants 2020, 9, 1612. https://doi.org/10.3390/plants9111612
Nawaz MA, Azeem F, Zakharenko AM, Lin X, Atif RM, Baloch FS, Chan T-F, Chung G, Ham J, Sun S, et al. In-silico Exploration of Channel Type and Efflux Silicon Transporters and Silicification Proteins in 80 Sequenced Viridiplantae Genomes. Plants. 2020; 9(11):1612. https://doi.org/10.3390/plants9111612
Chicago/Turabian StyleNawaz, Muhammad Amjad, Farrukh Azeem, Alexander Mikhailovich Zakharenko, Xiao Lin, Rana Muhammad Atif, Faheem Shehzad Baloch, Ting-Fung Chan, Gyuhwa Chung, Junghee Ham, Sangmi Sun, and et al. 2020. "In-silico Exploration of Channel Type and Efflux Silicon Transporters and Silicification Proteins in 80 Sequenced Viridiplantae Genomes" Plants 9, no. 11: 1612. https://doi.org/10.3390/plants9111612