1. Introduction
Specialized orchards of the arid or semiarid regions of the Mediterranean basin are often characterized by low levels of soil organic matter and severe weed infestations, which need a frequent use of chemical inputs for their management [
1]. In these agroecosystems, weeds represent the most serious constraint to agricultural production, causing serious yield losses due to their highly competitive capacity and allelopathic activity [
2,
3]. For many decades, they have been controlled almost exclusively through an irrational use of herbicides that, in addition to the negative effects on the environment, humans and animals [
4,
5], caused a significant reduction of biodiversity [
6]. Low biodiversity in agroecosystems is associated not only to the development of a selective weed flora more difficult to manage, but also to a greater vulnerability to new invasive species [
7]. Both weed abundance and diversity are closely influenced by agricultural practices, mainly soil tillage systems, crop rotation and fertilization [
8], with a central role played also by environmental conditions [
9,
10]. The effects (positive or negative) of agronomic techniques on weed diversity are unclear and contradictory, depending on the specific conditions of field experiments, while conservation tillage systems are commonly reported to increase weed abundance [
11]. Nowadays, given the increasing interest in limiting the dependence on herbicides, weed control in croplands is addressing to find ecologically-based practices (e.g., crop rotation, stale seedbed, cover cropping, mechanical and physical methods, etc.) under an integrated approach in a medium–long-term strategy [
3]. The basic principle is that weeds are an integral part of the agroecosystem and, thus, they should be managed to reduce their harmful effects and increase benefits [
12]. Integrated weed management systems are not absolute, but may vary in relation to the context-specific requirements and from year to year.
One of the most common eco-friendly practices, commonly adopted in organic and low-input agricultural systems, is cover cropping, which in the present study is going to include the techniques of mulching, intercropping and green manuring. Indeed, cover crops can be used as living mulches when intercropped between rows in herbaceous crops or on the whole field surface in tree crops, as well as dead mulches either on the soil surface or buried into the soil [
3]. In both cases, they prevent weed germination and emergence physically by increasing the competition with weeds and chemically through allelopathic mechanisms [
13]. In addition to the phytotoxic activity, cover crops are referred to increase soil fertility by reducing erosion and nutrient leaching, while improving the organic matter content, soil structure and microbial activities [
14]. Among the high number of cover crops used in agroecosystems, the
Trifolium genus and subterranean clover (
T. subterraneum) in particular, play a key role in Mediterranean orchards thanks to N-fixation ability, rapid growth, rusticity, allelopathic activity and resistance to low radiation levels [
15,
16]. Subterranean clover originated in the Mediterranean basin, from where it spread throughout western Europe, northern Africa and other world regions with Mediterranean-type climates including Americas, New Zealand and mainly in southern Australia, where it is actually the major pasture legume [
17]. It is a free-seeding annual legume, diploid (2n = 16) and predominantly self-pollinated, with remarkable geocarpism. Despite the dispute about the intraspecific taxonomy of
T. subterraneum, recent genetic studies have confirmed the original classification provided by Katznelson and Morley [
18], according to whom the species includes three subspecies with different ecological behavior: subsp.
subterraneum, subsp.
yanninicum and subsp.
brachycalycinum.
In a recent study, Scavo et al. [
1] demonstrated that
T. subterraneum cover cropping significantly reduced the size of the weed seed bank, enhanced the amount of soil nitrogen bacteria and increased the levels of ammoniacal and nitric soil nitrogen. Developed as a continuation of the above-mentioned study, in this research we hypothesized that the observed changes in the potential weed flora (soil seed bank) could reflect on the real one in terms of abundance, richness and diversity, all key aspects for the development of an optimal integrated weed management strategy. Therefore, the objective of this work was to determine the long-term effect of
T. subterraneum and spontaneous flora cover crops, with respect to a conventional management, on the aboveground weed populations and species composition in an apricot orchard.
3. Discussion
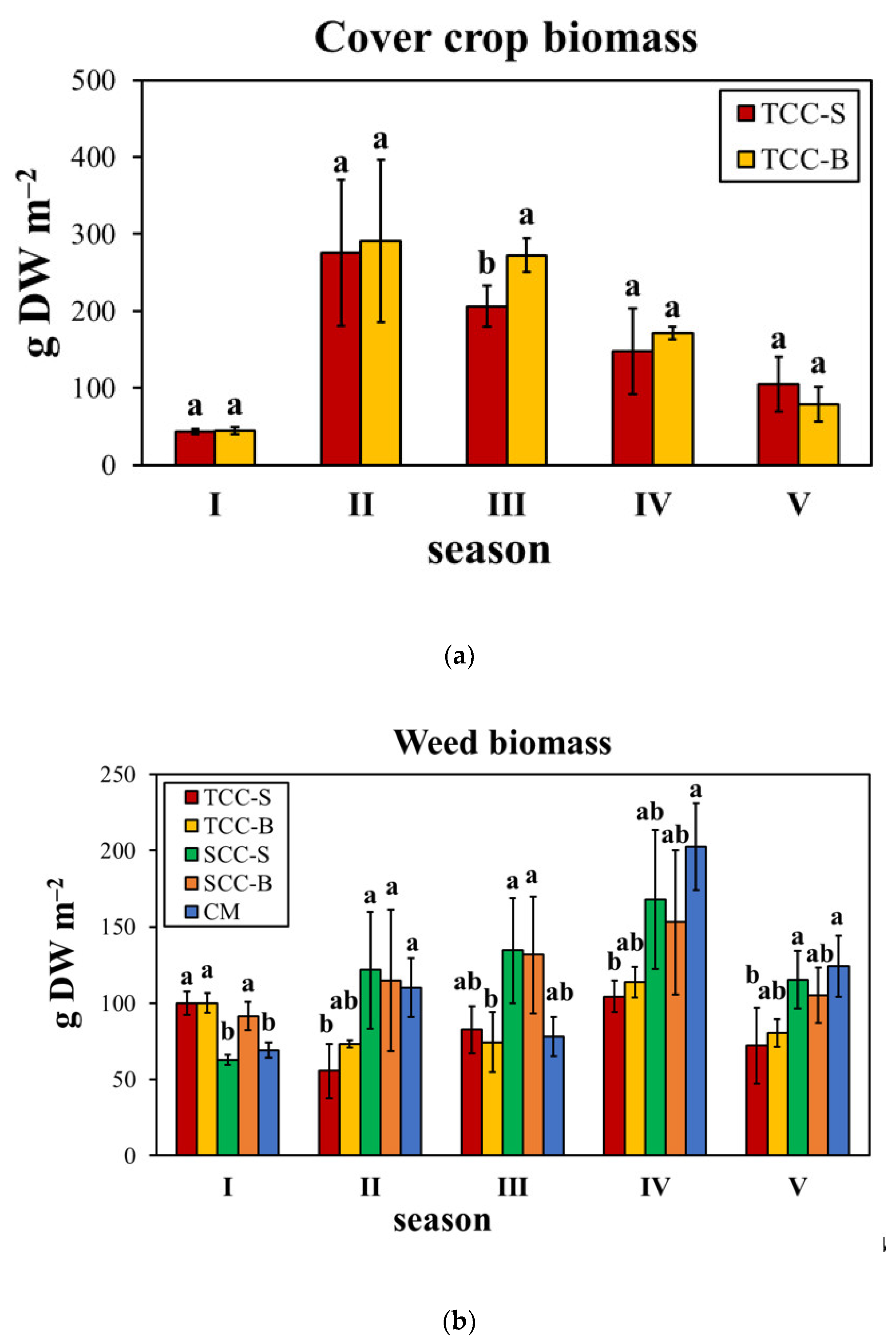
The present study aimed to evaluate the influence of 5 years of cover cropping, by subterranean clover and spontaneous flora, both buried and living dead mulches on the soil surface, on diversity and abundance of the real weed flora. In our previous research [
1], we found a 70% reduction of the weed soil seed bank, compared to CM, after 3-years of
T. subterraneum green manuring (TCC-B). Given that the real weed flora generally reflects the spectrum of the potential one, the effects on the emerged weeds were evaluated for a further two years on a medium–long-term period. We found that subterranean clover, in some seasons, significantly decreased the mean weed biomass up to 86%, contrariwise to spontaneous flora cover crop. The intensity of such a decrease was season-dependent, likely due to a combined effect of climatic conditions and cover crop biomass. In contrast with Moonen and Bàrberi [
19], in our study cover crop biomass highly varied between the seasons from 44 to more than 290 g of DW m
‒2. Weed biomass decrease caused by subterranean clover was higher in seasons when cover crop biomass was higher (seasons II, IV and V), except for season III. Our results are similar to those obtained by the study of Bàrberi and Mazzoncini [
20], in which subterranean clover was found to reduce weed biomass from 21% to 67%, with a positive correlation between weed growth suppression and cover crop biomass and with seasonal effects. The results obtained on the real flora were corroborated by the effects on the soil seed bank, in which all the cover cropping systems decreased the number of weed seeds. TCC-S and TCC-B showed the highest weed suppressive ability after a further two years, although with a lower degree than the third year [
1].
Weed suppressive ability of subterranean clover may be attributed to competitive or allelopathic effects, or even to a combination of them.
Trifolium subterraneum, in fact, competes well with weeds thanks to its rapid growth, developed canopy, length of biological cycle and development of root system [
21]. Generally, weed suppression increases with increasing cover crop biomass and cycle length, as found in the present study. Furthermore, subterranean clover is recognized as allelopathic species and allelochemicals responsible for such phytotoxic effects have been indicated as phenols and isoflavonoids [
22]. These secondary metabolites can be directly exuded into the soil or released by decomposition of plant residues. Once present into the rhizosphere, allelochemicals interact with the complex of physical, chemical and biological soil characteristics, which altogether fix their availability [
23]. Unfortunately, competitive and allelopathic effects are very difficult to distinguish in field experiments.
The emerged flora reflected the composition of the seed bank, given that weed communities were dominated by Asteraceae members, therophytes and annual spring–summer weeds. As previously observed on the seed bank [
1], weed species richness was significantly affected by TCC-S, while, interestingly, TCC-B increased it, suggesting no clear influence of cover cropping. On the contrary, the effect of season was more noticeable, with a much higher number of weed species detected in season V, showing an increase in weed biodiversity. Conflicting reports have been provided by authors concerning the effects of cover crops on species richness. Ngouajio et al. [
24], for example, observed no significant relationships, with results depending on cover crop type and season, while a reduction of weed density was found by Moonen and Bàrberi [
19] using rye (
Secale cereale L.) cover crop.
Since the contradictory results, many authors agree in not considering species richness as the only parameter to evaluate the herbicidal activity of cover crops. In this regard, the composition of weed communities plays a key role in shifting the phytotoxic effects. It should be pointed out, in fact, that the sensitivity of weed species to cover crop residue is highly variable, mainly depending on weed community structure. On one side, annual weeds with small seed sizes are more susceptible to surface residues than large seeded species, and on the other side, large seeds have a greater metabolic capacity for allelochemical detoxification [
25]. In this study, the β-diversity indices of Jaccard and Sørensen were applied the compare the areas in terms of composition of the weed communities [
26]. These indices are closely influenced by agronomic practices. Here, the highest similarity was found between spontaneous flora (SCC-S × SCC-B) and between subterranean clover (TCC-S × TCC-B) cover crop, often with values across seasons higher than 50%, at which an elevated similarity can be interpreted. Instead, a general low similarity was found between the conventional management and cover cropping systems. Therefore, it is reasonable to assume that
T. subterraneum treatments (TCC-S and TCC-B) determined similar weed communities based on presence/absence, as well as spontaneous flora cover crops (SCC-S and SCC-B), both different with respect to CM. ANOVA performed on RF data of single species, however, pointed out any significant effect among treatments under study, demonstrating that weeds were able to establish independently of cover type and season. To overcome the complexity of weed data, species composition was studied by PCA on major weed species. In addition to a reduction in weed seed bank density and aboveground biomass, TCC-B did not show any association with weeds, contrariwise to SCC-S and SCC-B. No evident weed patterns emerged in this study, as observed also in the seed bank [
1]. Overall, treatments were quite similar also with reference to frequency, botanical families, life cycle, biological and ecophysiological groups. The lack of consistent associations between cover crop and weeds has been reported in many other studies [
19,
27], since species composition can be influenced by abiotic and biotic factors. In the 9-year research study carried out by Shrestha et al. [
28] on winter wheat and three beans, rye and maize cover crop were also indicated to have differential effects on weed densities, species composition and associations depending on crop type and interaction with agronomic management.
In conclusion, this research suggests that long-term changes in weed flora are linked to the soil seed bank. On one hand, the adoption of 5 years of cover cropping with subterranean clover was found to reduce not only the number of weed seeds in the soil, but also the aboveground weed biomass and the number of species, with significant variations by season. On the other hand, instead, no clear shifts in weed populations were observed. These results are very useful in view of reducing intensive tillage and the frequent application of herbicides, thus allowing multiple benefits for the environment. The benefits in using subterranean clover in Mediterranean agroecosystems are further increased considering its self-reseed capacity, N-fixation ability and high adaptability in such contexts [
1]. Future studies may consider the evaluation of subterranean clover cover cropping in combination with other control techniques under an integrated weed management system, as well as a better knowledge of the mechanisms involved in its phytotoxicity.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}