Prediction of Dhurrin Metabolism by Transcriptome and Metabolome Analyses in Sorghum

 , and
, and
Abstract
:1. Introduction
2. Results
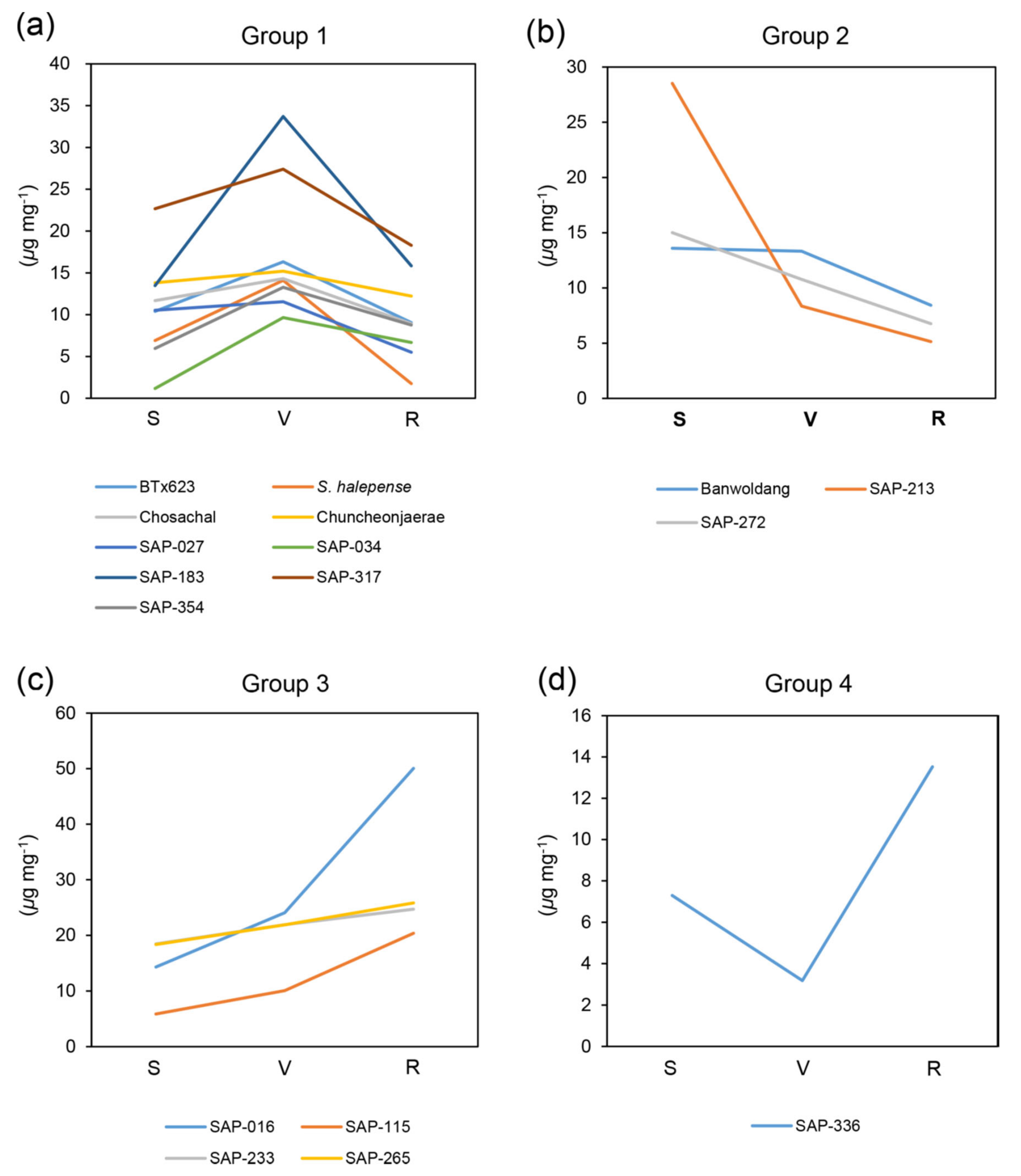
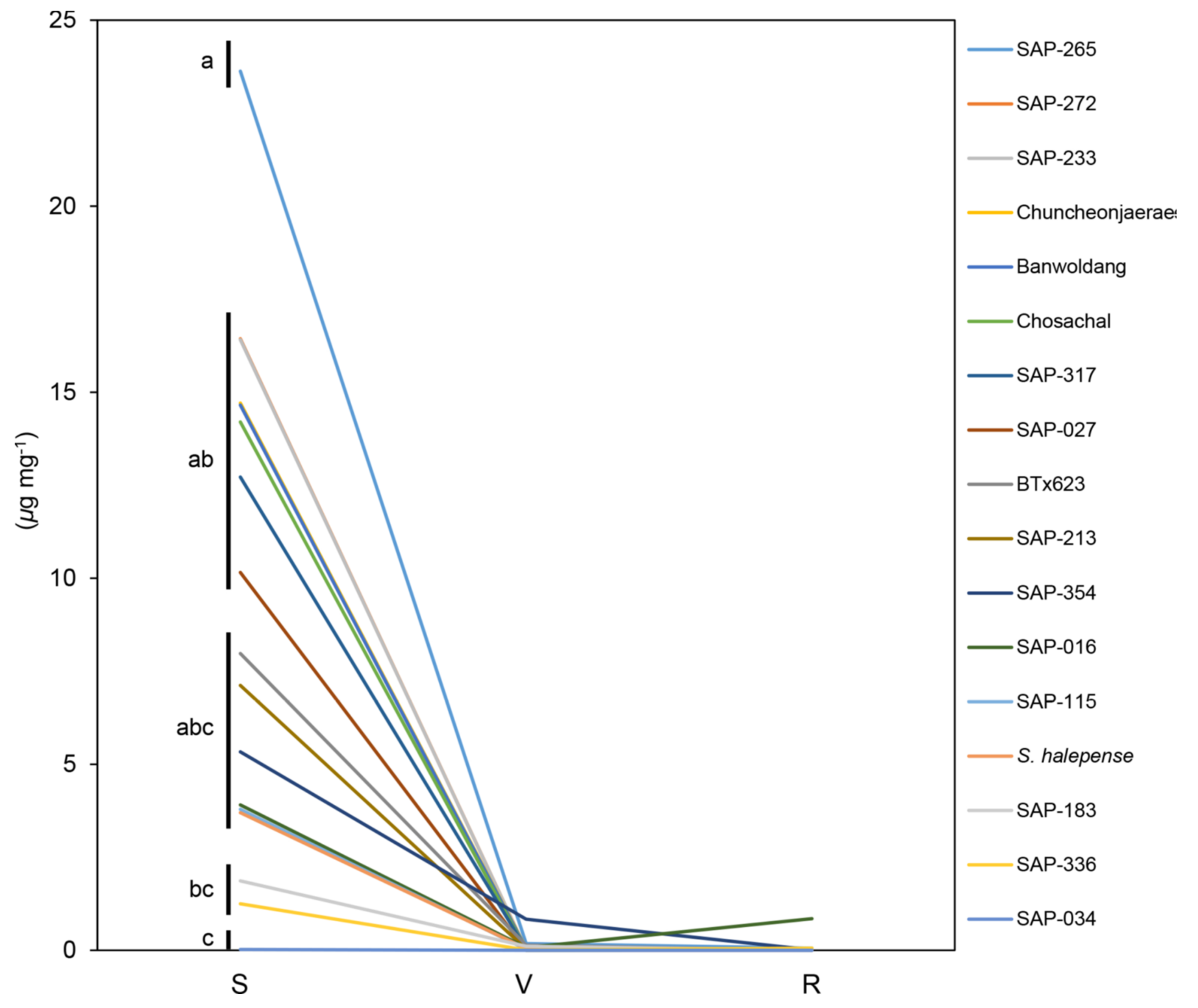
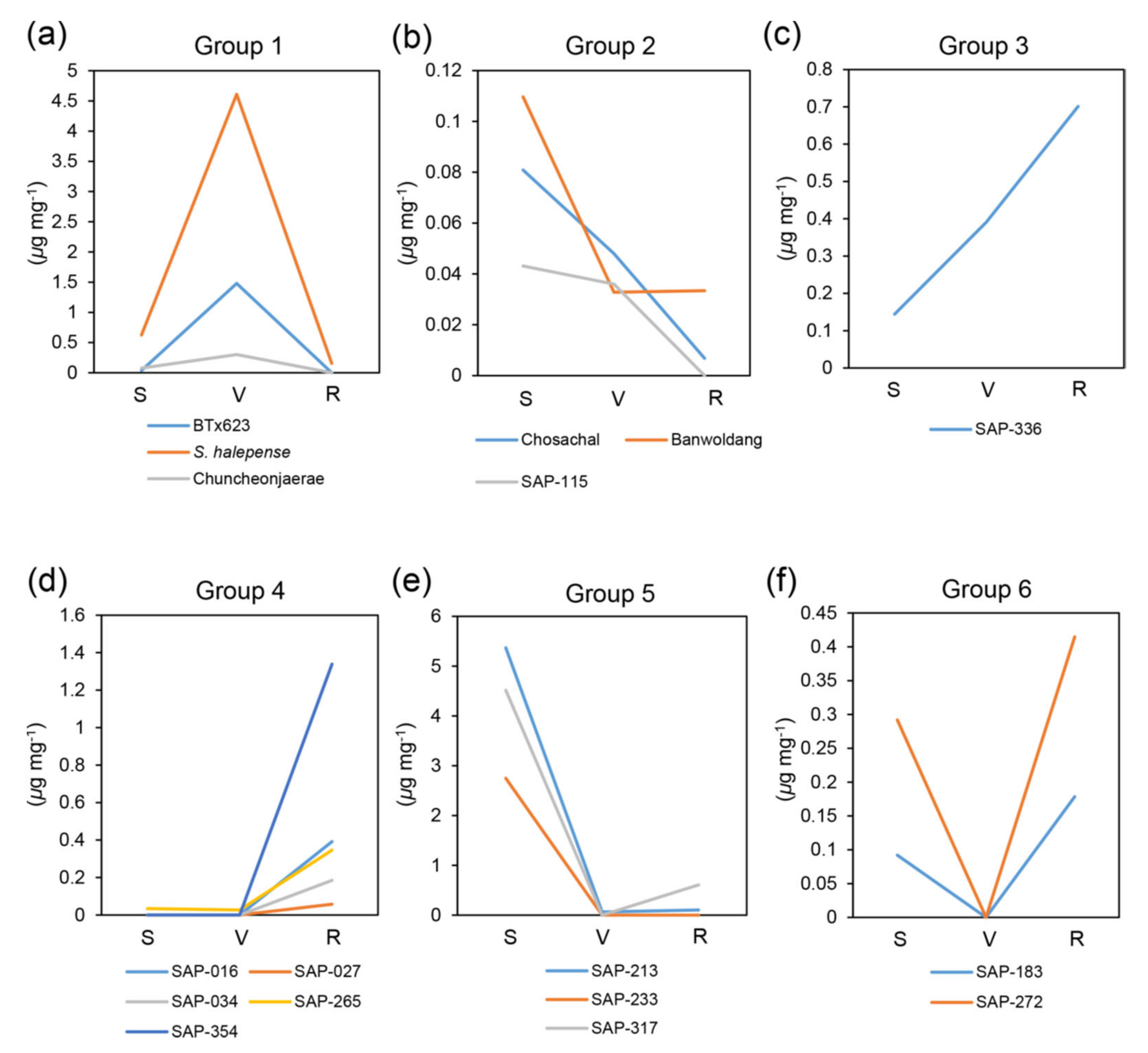
2.1. Metabolite Analysis at Different Developmental Stages
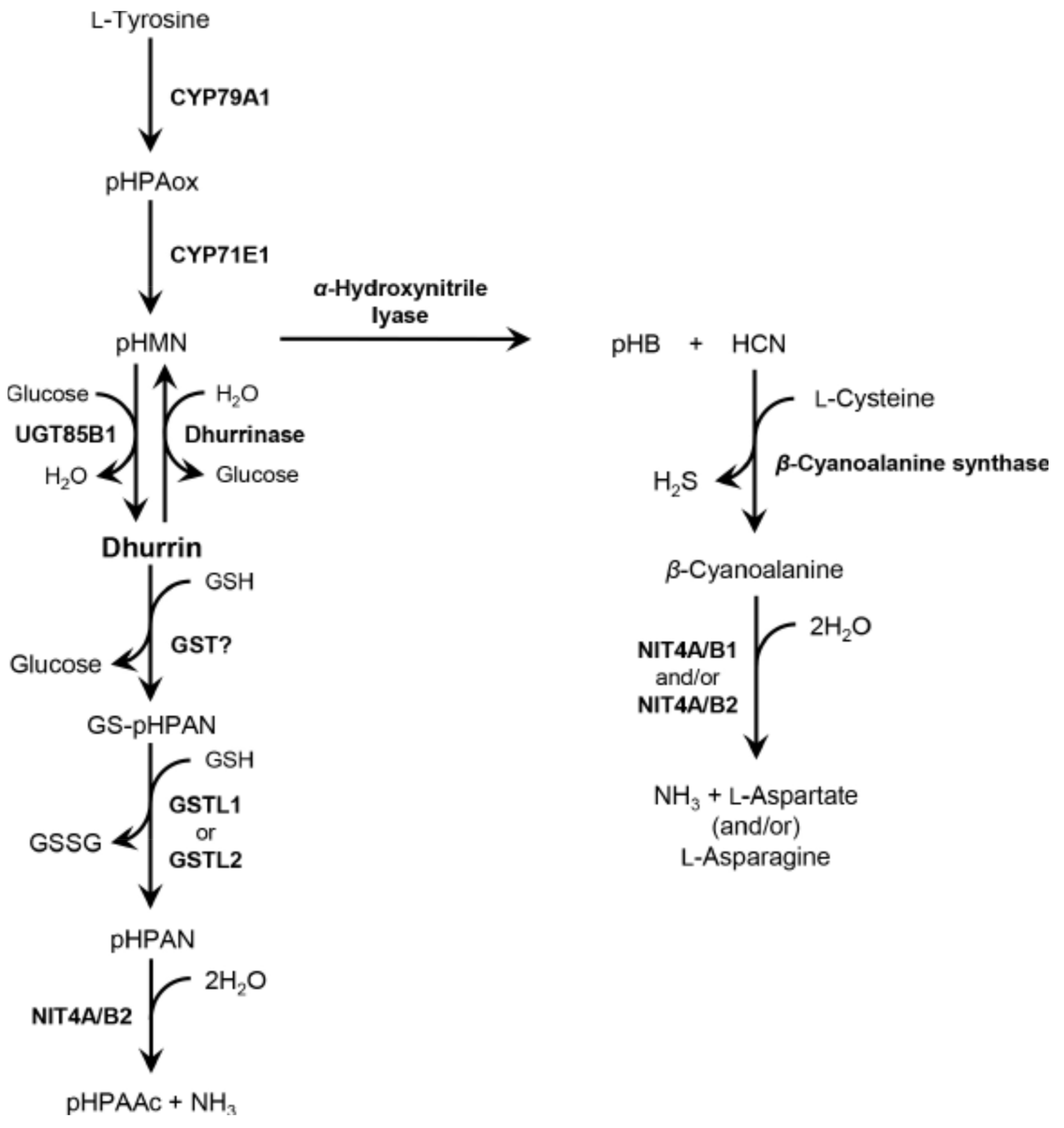
2.2. Identification of Genes Involved in Dhurrin Metabolism
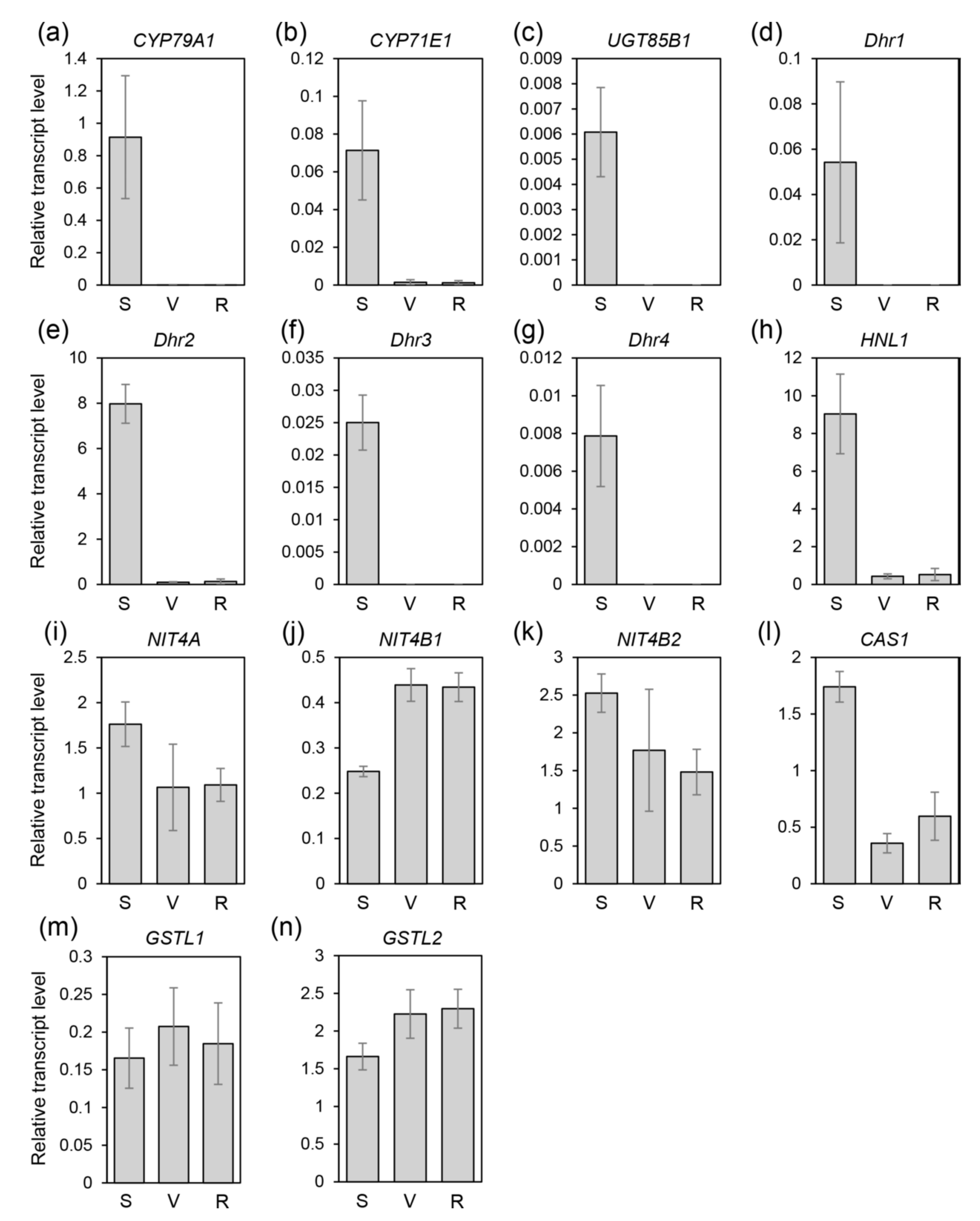
2.3. Quantitative RT-PCR Analysis of Candidate Genes
2.4. Correlation of Metabolite Contents and Gene Expression Levels
2.5. SNP Analysis through RNAseq
3. Discussion
3.1. Correlation of Metabolite Contents and Expression Profiles
3.2. Function of Dhurrinases
3.3. Complex Dhurrin Metabolism
3.4. The Effect of Gene Expressions on HCN Detoxification
3.5. Functional Differentiation of NIT4 Enzymes
3.6. Identification of SNPs in Dhurrin Degradation
3.7. Perspectives of Sorghum Materials and Breeding
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Metabolite Extraction
4.3. HPLC Analysis
4.4. RNA Extraction and cDNA Synthesis
4.5. Quantitative RT-PCR Analysis
4.6. Statistical Analysis
4.7. Library Construction and Sequencing
4.8. Transcript Assembly and Abundance Estimation Using Cufflinks
4.9. Analysis of SNPs Associated with the Candidate Genes
4.10. Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kimber, C.T.; Dahlberg, J.A.; Kresovich, S. The gene pool of Sorghum bicolor and its improvement. In Genomics of the Saccharinae; Springer: New York, NY, USA, 2013; pp. 23–41. [Google Scholar]
- Etuk, E.; Ifeduba, A.; Okata, U.; Chiaka, I.; Okoli, I.C.; Okeudo, N.; Esonu, B.; Udedibie, A.; Moreki, J. Nutrient composition and feeding value of sorghum for livestock and poultry: A review. J. Anim. Sci. Adv. 2012, 2, 510–524. [Google Scholar]
- Proietti, I.; Frazzoli, C.; Mantovani, A. Exploiting nutritional value of staple foods in the world’s semi-arid areas: Risks, benefits, challenges and opportunities of sorghum. Healthcare 2015, 3, 172–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, N.W.; Mickan, F.J.; Kaiser, A.G. Crops and by-products for silage. In TopFodder Successful Silage, 2nd ed.; Dairy Australia and New South Wales Department of Primary Industries: Orange, NSW, Australia, 2004; pp. 109–142. [Google Scholar]
- Zagrobelny, M.; Bak, S.; Møller, B.L. Cyanogenesis in plants and arthropods. Phytochemistry 2008, 69, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.J.; Stuart, P.; Pičmanová, M.; Rasmussen, S.; Olsen, C.E.; Harholt, J.; Møller, B.L.; Bjarnholt, N. Dhurrin metabolism in the developing grain of Sorghum bicolor (L.) Moench investigated by metabolite profiling and novel clustering analyses of time-resolved transcriptomic data. BMC Genom. 2016, 17, 1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleadow, R.M.; Møller, B.L. Cyanogenic glycosides: Synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant. Biol. 2014, 65, 155–185. [Google Scholar] [CrossRef] [PubMed]
- Sibbesen, O.; Koch, B.; Halkier, B.A.; Møller, B.L. Isolation of the heme-thiolate enzyme cytochrome P-450TYR, which catalyzes the committed step in the biosynthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor (L.) Moench. Proc. Natl. Acad. Sci. USA 1994, 91, 9740–9744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, S.; Kahn, R.A.; Nielsen, H.L.; Møller, B.L.; Halkier, B.A. Cloning of three A-type cytochromes P450, CYP71E1, CYP98, and CYP99 from Sorghum bicolor (L.) Moench by a PCR approach and identification by expression in Escherichia coli of CYP71E1 as a multifunctional cytochrome P450 in the biosynthesis of the cyanogenic glucoside dhurrin. Plant. Mol. Biol. 1998, 36, 393–405. [Google Scholar]
- Jones, P.R.; Møller, B.L.; Høj, P.B. The UDP-glucose: P-hydroxymandelonitrile-O-glucosyltransferase that catalyzes the last step in synthesis of the cyanogenic glucoside dhurrin in sorghum bicolor isolation, cloning, heterologous expression, and substrate specificity. J. Biol. Chem. 1999, 274, 35483–35491. [Google Scholar] [CrossRef] [Green Version]
- Kahn, R.A.; Fahrendorf, T.; Halkier, B.A.; Møller, B.L. Substrate Specificity of the Cytochrome P450 Enzymes CYP79A1 and CYP71E1 Involved in the Biosynthesis of the Cyanogenic Glucoside Dhurrin in Sorghum bicolor (L.) Moench. Arch. Biochem. Biophys. 1999, 363, 9–18. [Google Scholar] [CrossRef]
- Poulton, J.E. Cyanogenesis in plants. Plant Physiol. 1990, 94, 401–405. [Google Scholar] [CrossRef] [Green Version]
- Wajant, H.; Mundry, K.-W.; Pfizenmaier, K. Molecular cloning of hydroxynitrile lyase from Sorghum bicolor (L.). Homologies to serine carboxypeptidases. Plant Mol. Biol. 1994, 26, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Poulton, J.E.; Thayer, S.S.; Conn, E.E. Tissue distributions of dhurrin and of enzymes involved in its metabolism in leaves of Sorghum bicolor. Plant Physiol. 1979, 63, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Jenrich, R.; Trompetter, I.; Bak, S.; Olsen, C.E.; Møller, B.L.; Piotrowski, M. Evolution of heteromeric nitrilase complexes in Poaceae with new functions in nitrile metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 18848–18853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Kliebenstein, D.J.; Gershenzon, J. The Arabidopsis epithiospecifier protein promotes the hydrolysis of glucosinolates to nitriles and influences Trichoplusia ni herbivory. Plant Cell 2001, 13, 2793–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnholt, N.; Neilson, E.H.; Crocoll, C.; Jørgensen, K.; Motawia, M.S.; Olsen, C.E.; Dixon, D.P.; Edwards, R.; Møller, B.L. Glutathione transferases catalyze recycling of auto-toxic cyanogenic glucosides in sorghum. Plant J. 2018, 94, 1109–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowski, M.; Schönfelder, S.; Weiler, E.W. The Arabidopsis thaliana isogene NIT4 and its orthologs in tobacco encode β-cyano-L-alanine hydratase/nitrilase. J. Biol. Chem. 2001, 276, 2616–2621. [Google Scholar] [CrossRef] [Green Version]
- Pičmanová, M.; Neilson, E.H.; Motawia, M.S.; Olsen, C.E.; Agerbirk, N.; Gray, C.J.; Flitsch, S.; Meier, S.; Silvestro, D.; Jørgensen, K. A recycling pathway for cyanogenic glycosides evidenced by the comparative metabolic profiling in three cyanogenic plant species. Biochem. J. 2015, 469, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Mizuno, H.; Kawahigashi, H.; Kawahara, Y.; Kanamori, H.; Ogata, J.; Minami, H.; Itoh, T.; Matsumoto, T. Global transcriptome analysis reveals distinct expression among duplicated genes during sorghum-Bipolaris sorghicola interaction. BMC Plant Biol. 2012, 12, 121. [Google Scholar] [CrossRef] [Green Version]
- Wurtele, E.S.; Nikolau, B.J.; Conn, E.E. Tissue distribution of β-cyanoalanine synthase in leaves. Plant Physiol. 1984, 75, 979–982. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (Austin) 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halkier, B.A.; Møller, B.L. Biosynthesis of the cyanogenic glucoside dhurrin in seedlings of Sorghum bicolor (L.) Moench and partial purification of the enzyme system involved. Plant Physiol. 1989, 90, 1552–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busk, P.K.; Møller, B.L. Dhurrin synthesis in sorghum is regulated at the transcriptional level and induced by nitrogen fertilization in older plants. Plant Physiol. 2002, 129, 1222–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicollier, G.F.; Pope, D.F.; Thompson, A.C. Biological activity of dhurrin and other compounds from Johnson grass (Sorghum halepense). J. Agric. Food. Chem. 1983, 31, 744–748. [Google Scholar] [CrossRef]
- Hösel, W.; Tober, I.; Eklund, S.H.; Conn, E.E. Characterization of β-glucosidases with high specificity for the cyanogenic glucoside dhurrin in Sorghum bicolor (L.) moench seedlings. Arch. Biochem. Biophys. 1987, 252, 152–162. [Google Scholar] [CrossRef]
- Cicek, M.; Esen, A. Structure and expression of a dhurrinase (β-glucosidase) from sorghum. Plant Physiol. 1998, 116, 1469–1478. [Google Scholar] [CrossRef] [Green Version]
- Krothapalli, K.; Buescher, E.M.; Li, X.; Brown, E.; Chapple, C.; Dilkes, B.P.; Tuinstra, M.R. Forward genetics by genome sequencing reveals that rapid cyanide release deters insect herbivory of Sorghum bicolor. Genetics 2013, 195, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Hayes, C.M.; Burow, G.B.; Brown, P.J.; Thurber, C.; Xin, Z.; Burke, J.J. Natural variation in synthesis and catabolism genes influences dhurrin content in sorghum. Plant Genome 2015, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, C.; Patel, K.; Khan, B.M.; Rawal, S.S. In silico ligand binding studies of cyanogenic β-glucosidase, dhurrinase-2 from Sorghum bicolor. J. Mol. Model 2015, 21, 184. [Google Scholar] [CrossRef]
- Blanchard, D.J.; Cicek, M.; Chen, J.; Esen, A. Identification of β-glucosidase aggregating factor (BGAF) and mapping of BGAF binding regions on maize β-glucosidase. J. Biol. Chem. 2001, 276, 11895–11901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittur, F.S.; Yu, H.Y.; Bevan, D.R.; Esen, A. Homolog of the maize β-glucosidase aggregating factor from sorghum is a jacalin-related GalNAc-specific lectin but lacks protein aggregating activity. Glycobiology 2009, 19, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Kittur, F.S.; Yu, H.Y.; Bevan, D.R.; Esen, A. Deletion of the N-terminal dirigent domain in maize β-glucosidase aggregating factor and its homolog sorghum lectin dramatically alters the sugar-specificities of their lectin domains. Plant Physiol. Biochem. 2010, 48, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Siegień, I.; Bogatek, R. Cyanide action in plants—From toxic to regulatory. Acta Physiol. Plant. 2006, 28, 483–497. [Google Scholar] [CrossRef]
- Paterson, A.H.; Chapman, B.A.; Kissinger, J.C.; Bowers, J.E.; Feltus, F.A.; Estill, J.C. Many gene and domain families have convergent fates following independent whole-genome duplication events in Arabidopsis, Oryza, Saccharomyces and Tetraodon. Trends Genet. 2006, 22, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, J.; Miller, F.; Rosenow, D.T. Conversion of alien Sorghums to early combine genotypes. Crop. Sci. 1967, 7, 396. [Google Scholar] [CrossRef]
- Gabriel, K. A Study of Heterotic Relationships in Sorghum. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2006. [Google Scholar]
- Casa, A.M.; Pressoir, G.; Brown, P.J.; Mitchell, S.E.; Rooney, W.L.; Tuinstra, M.R.; Franks, C.D.; Kresovich, S. Community resources and strategies for association mapping in sorghum. Crop Sci. 2008, 48, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D.; Riera-Lizarazu, O.; Brown, P.J.; Acharya, C.B.; Mitchell, S.E. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- De Nicola, G.R.; Leoni, O.; Malaguti, L.; Bernardi, R.; Lazzeri, L. A simple analytical method for dhurrin content evaluation in cyanogenic plants for their utilization in fodder and biofumigation. J. Agric. Food Chem. 2011, 59, 8065–8069. [Google Scholar] [CrossRef]
- Haskins, F.A.; Gorz, H.J.; Hill, R.M. Colorimetric determination of cyanide in enzyme-hydrolyzed extracts of dried sorghum leaves. J. Agric. Food Chem. 1988, 36, 775–778. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Zhang, C.-H.; Cao, X.L. RP-HPLC Analysis of Phenolic Acids in Different Varieties of Cereal Bran. Food Sci. 2010, 10. Available online: https://en.cnki.com.cn/Article_en/CJFDTotal-SPKX201010058.htm (accessed on 1 October 2020).
- Reddy, P.S.; Reddy, D.S.; Sivasakthi, K.; Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Evaluation of sorghum [Sorghum bicolor (L.)] reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dhurrin | pHB | pHPAAc | CYP79A1 | CYP71E1 | UGT85B1 | Dhr1 | Dhr2 | Dhr3 | Dhr4 | HNL1 | NIT4A | NIT4B1 | NIT4B2 | CAS1 | GSTL1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pHB | 0.071 | |||||||||||||||
| pHPAAc | 0.232 | 0.212 | ||||||||||||||
| CYP79A1 | −0.103 | 0.534 ** | 0.185 | |||||||||||||
| CYP71E1 | −0.098 | 0.419 ** | 0.107 | 0.956 ** | ||||||||||||
| UGT85B1 | 0.019 | 0.206 | 0.043 | 0.588 ** | 0.690 ** | |||||||||||
| Dhr1 | −0.112 | 0.363 ** | 0.284 * | 0.757 ** | 0.685 ** | 0.412 ** | ||||||||||
| Dhr2 | −0.020 | 0.639 ** | 0.253 | 0.639 ** | 0.519 ** | 0.191 | 0.650 ** | |||||||||
| Dhr3 | 0.001 | 0.752 ** | 0.270 | 0.505 ** | 0.364 ** | 0.184 | 0.521 ** | 0.762 ** | ||||||||
| Dhr4 | −0.124 | 0.696 ** | −0.042 | 0.565 ** | 0.496 ** | 0.146 | 0.373 ** | 0.497 ** | 0.503 ** | |||||||
| HNL1 | −0.103 | 0.463 ** | 0.180 | 0.577 ** | 0.454 ** | 0.246 | 0.464 ** | 0.582 ** | 0.617 ** | 0.433 ** | ||||||
| NIT4A | −0.282 * | 0.539 ** | 0.239 | 0.348 * | 0.261 | 0.016 | 0.323 * | 0.649 ** | 0.486 ** | 0.411 ** | 0.290 * | |||||
| NIT4B1 | 0.173 | −0.069 | −0.098 | 0.111 | 0.117 | 0.155 | −0.006 | 0.064 | −0.009 | −0.062 | 0.273 | −0.132 | ||||
| NIT4B2 | −0.092 | 0.470 ** | 0.387 ** | 0.414 ** | 0.305 * | 0.215 | 0.504 ** | 0.687 ** | 0.546 ** | 0.345 * | 0.466 ** | 0.758 ** | 0.048 | |||
| CAS1 | −0.173 | 0.635 ** | 0.140 | 0.534 ** | 0.490 ** | 0.380 ** | 0.498 ** | 0.475 ** | 0.524 ** | 0.531 ** | 0.383 ** | 0.571 ** | 0.119 | 0.575 ** | ||
| GSTL1 | −0.049 | 0.394 ** | 0.105 | 0.017 | −0.015 | −0.093 | 0.140 | 0.480 ** | 0.378 ** | 0.115 | 0.012 | 0.548 ** | 0.002 | 0.481 ** | 0.378 ** | |
| GSTL2 | 0.145 | −0.253 | −0.081 | −0.392 ** | −0.330 * | −0.100 | −0.352 * | −0.234 | −0.195 | −0.394 ** | −0.357 * | −0.218 | −0.037 | −0.326 * | −0.284 * | 0.195 |
| Location of SNP Marker | Reference | Substitute | Related Gene | Position | Associated Accession |
|---|---|---|---|---|---|
| Chr4:57544430 | C | T | NIT4B2 | 4th intron | S. halepense, SAP-336, SAP-354 |
| Chr6:2666100 | C | T | CAS1 | 5′ UTR | S. halepense, SAP-336 |
| Chr6:2666092 | CCACCAAAG | G | CAS1 | 5′ UTR | S. halepense |
| Chr6:2666088 | TGA | A | CAS1 | 5′ UTR | S. halepense |
| Chr6:2666085 | CT | T | CAS1 | 5′ UTR | S. halepense |
| Chr6:2666028 | GCTAGT | T | CAS1 | 5′ UTR | SAP-336 |
| Chr6:2666028 | GCTAAGCTAGT | T | CAS1 | 5′ UTR | S. halepense |
| Chr6:2665933 | G | ACGG | CAS1 | 1st exon | S. halepense |
| Chr6:2665870 | GC | C | CAS1 | 1st exon | S. halepense |
| Chr6:2665867 | GC | C | CAS1 | 1st exon | S. halepense |
| Chr6:2665864 | AGC | C | CAS1 | 1st exon | S. halepense |
| Chr6:2665840 | G | A | CAS1 | 1st exon | S. halepense |
| Chr6:2665335 | G | T | CAS1 | 2nd exon | S. halepense |
| Chr6:2665167 | C | AGC | CAS1 | 2nd intron | SAP-336 |
| Chr6:2665136 | A | GTTAA | CAS1 | 2nd intron | S. halepense |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.C.; Chung, Y.S.; Lee, Y.G.; Kang, Y.; Park, Y.J.; Park, S.U.; Kim, C. Prediction of Dhurrin Metabolism by Transcriptome and Metabolome Analyses in Sorghum. Plants 2020, 9, 1390. https://doi.org/10.3390/plants9101390
Choi SC, Chung YS, Lee YG, Kang Y, Park YJ, Park SU, Kim C. Prediction of Dhurrin Metabolism by Transcriptome and Metabolome Analyses in Sorghum. Plants. 2020; 9(10):1390. https://doi.org/10.3390/plants9101390
Chicago/Turabian StyleChoi, Sang Chul, Yong Suk Chung, Yun Gyeong Lee, Yuna Kang, Yun Ji Park, Sang Un Park, and Changsoo Kim. 2020. "Prediction of Dhurrin Metabolism by Transcriptome and Metabolome Analyses in Sorghum" Plants 9, no. 10: 1390. https://doi.org/10.3390/plants9101390