Heterophyllous Shoots of Japanese Larch Trees: The Seasonal and Yearly Variation in CO2 Assimilation Capacity of the Canopy Top with Changing Environment

 , and
, and 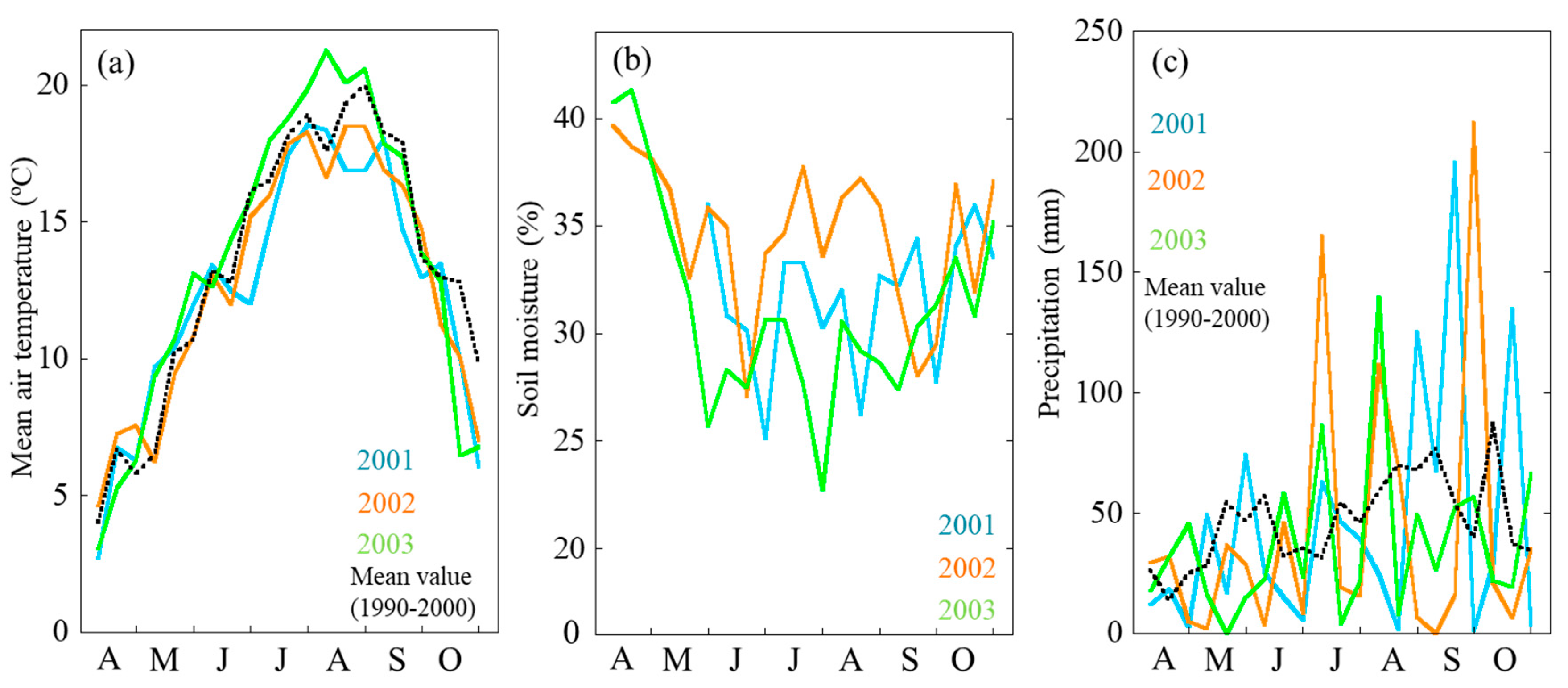{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Seasonal Changes of Air Temperature and Soil Moisture
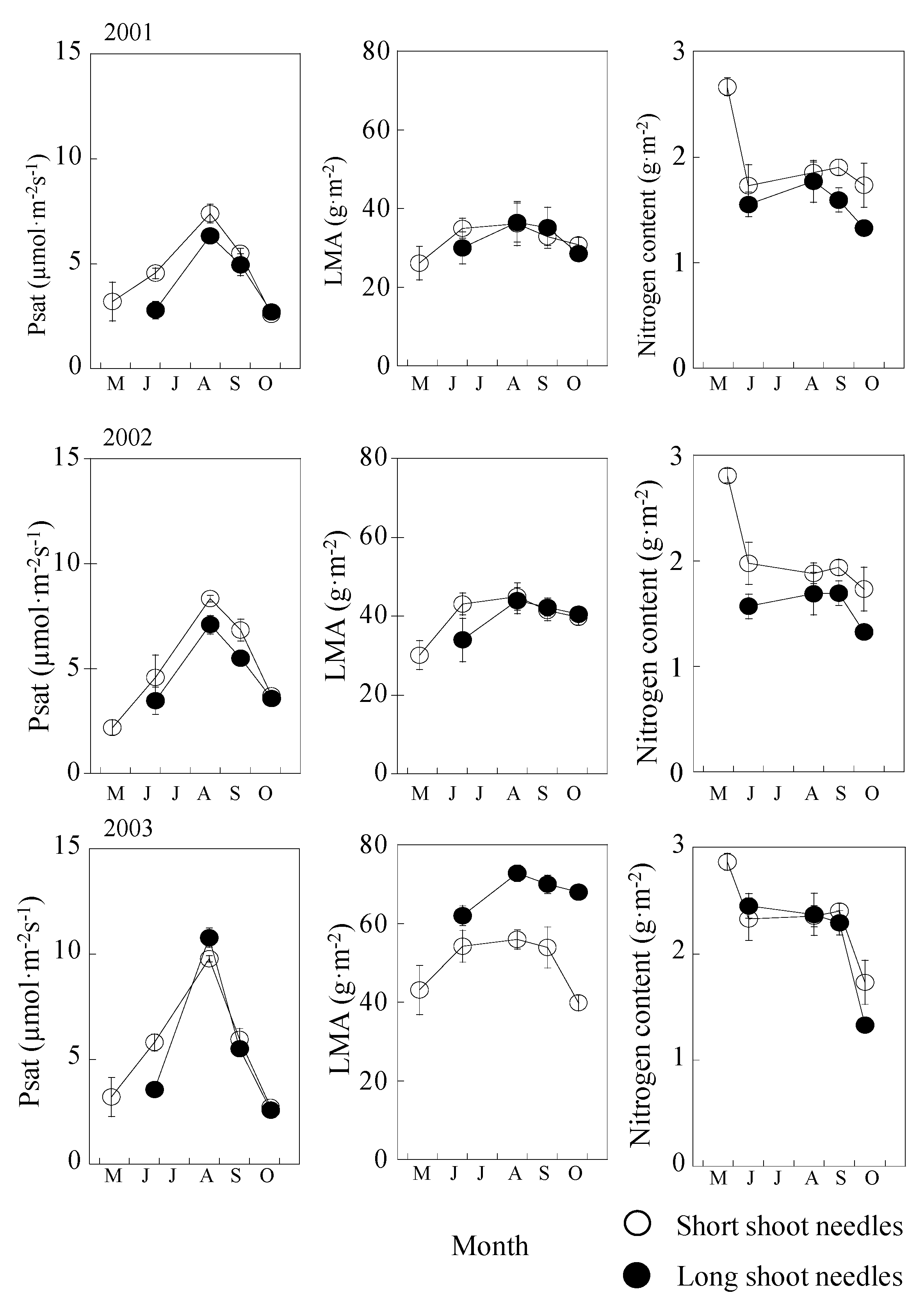
2.2. Seasonal and Annual Changes in Psat, LMA, and Nitrogen Content
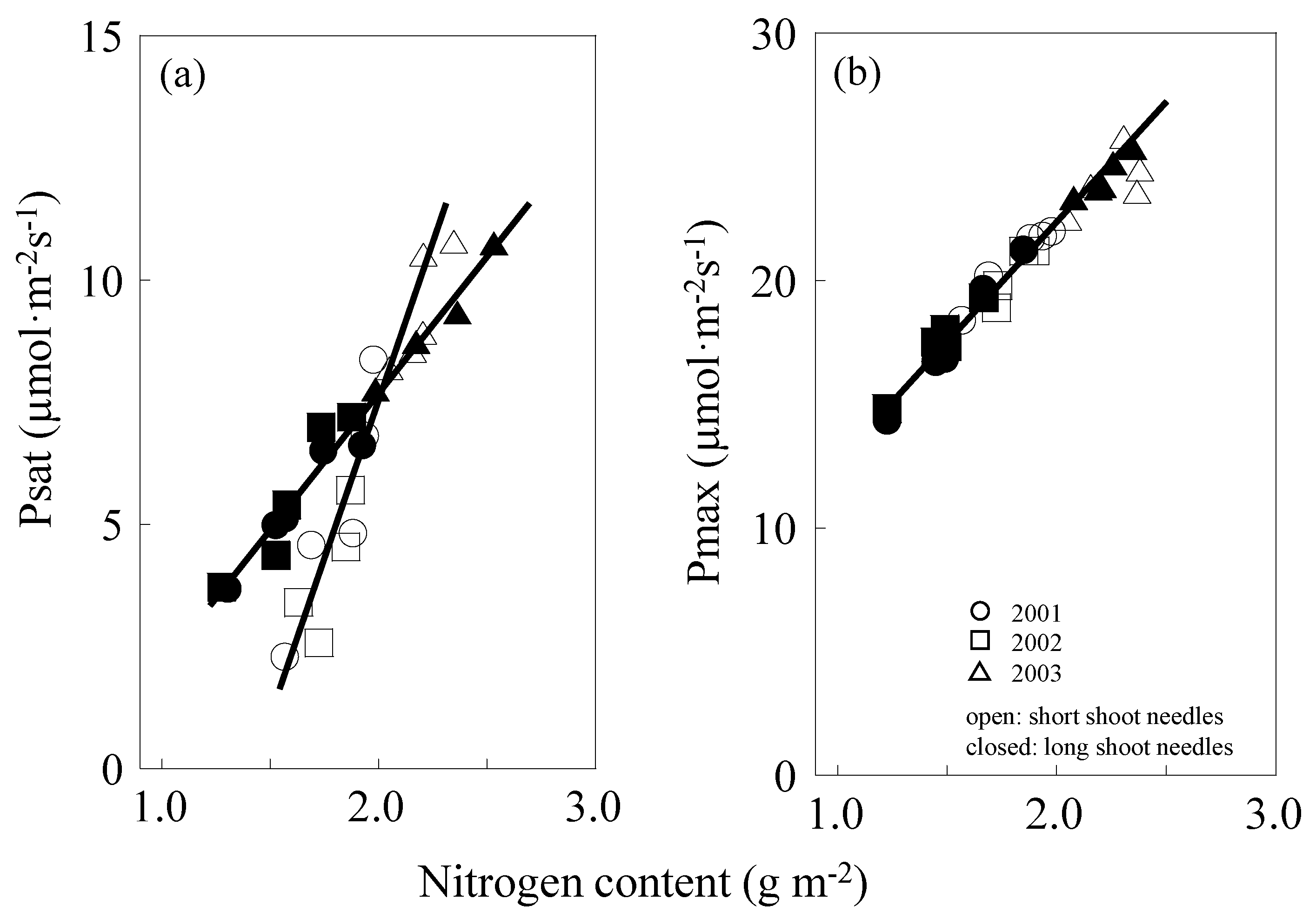
2.3. Photosynthesis–Nitrogen Relation
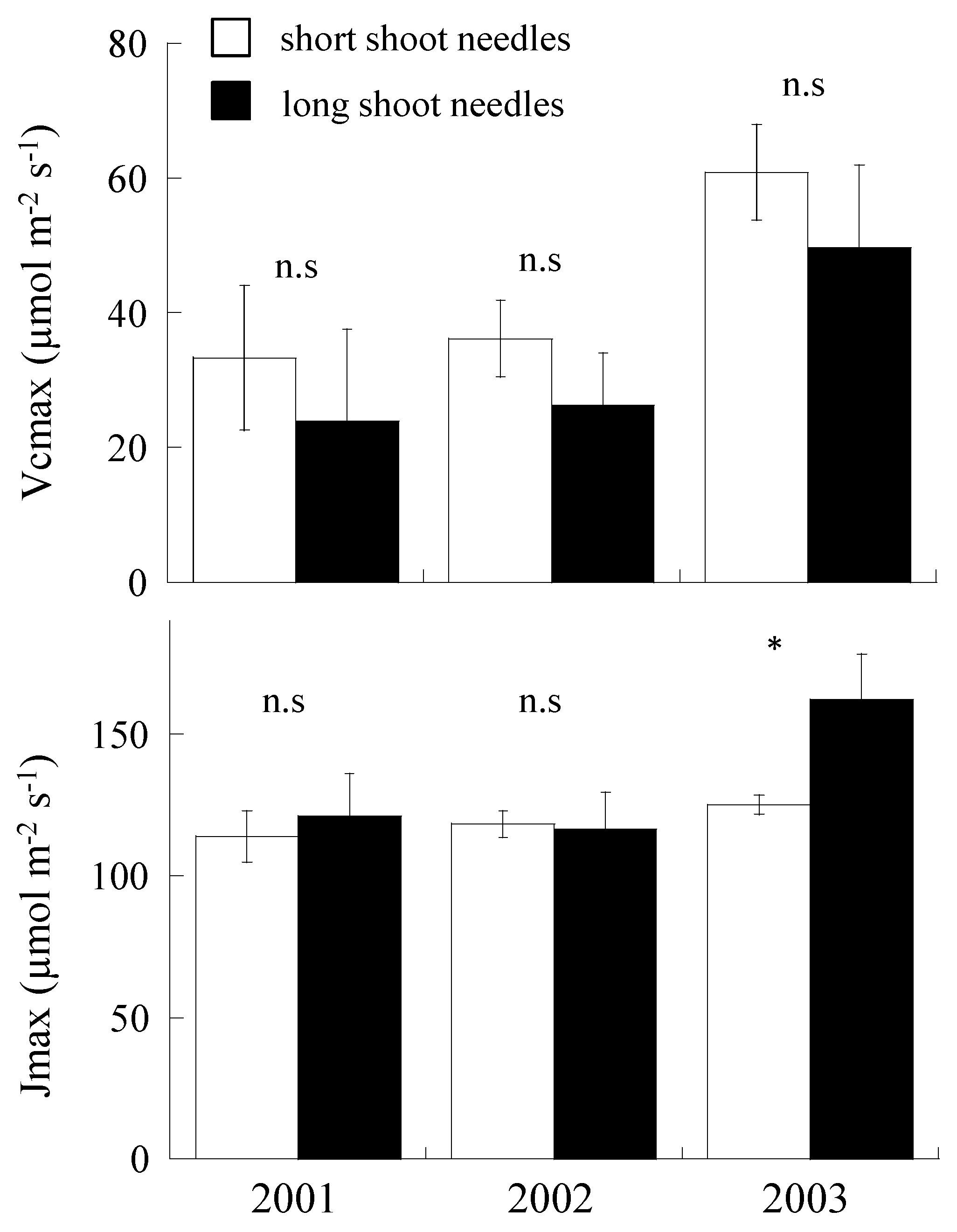
2.4. Variation of Vcmax and Jmax
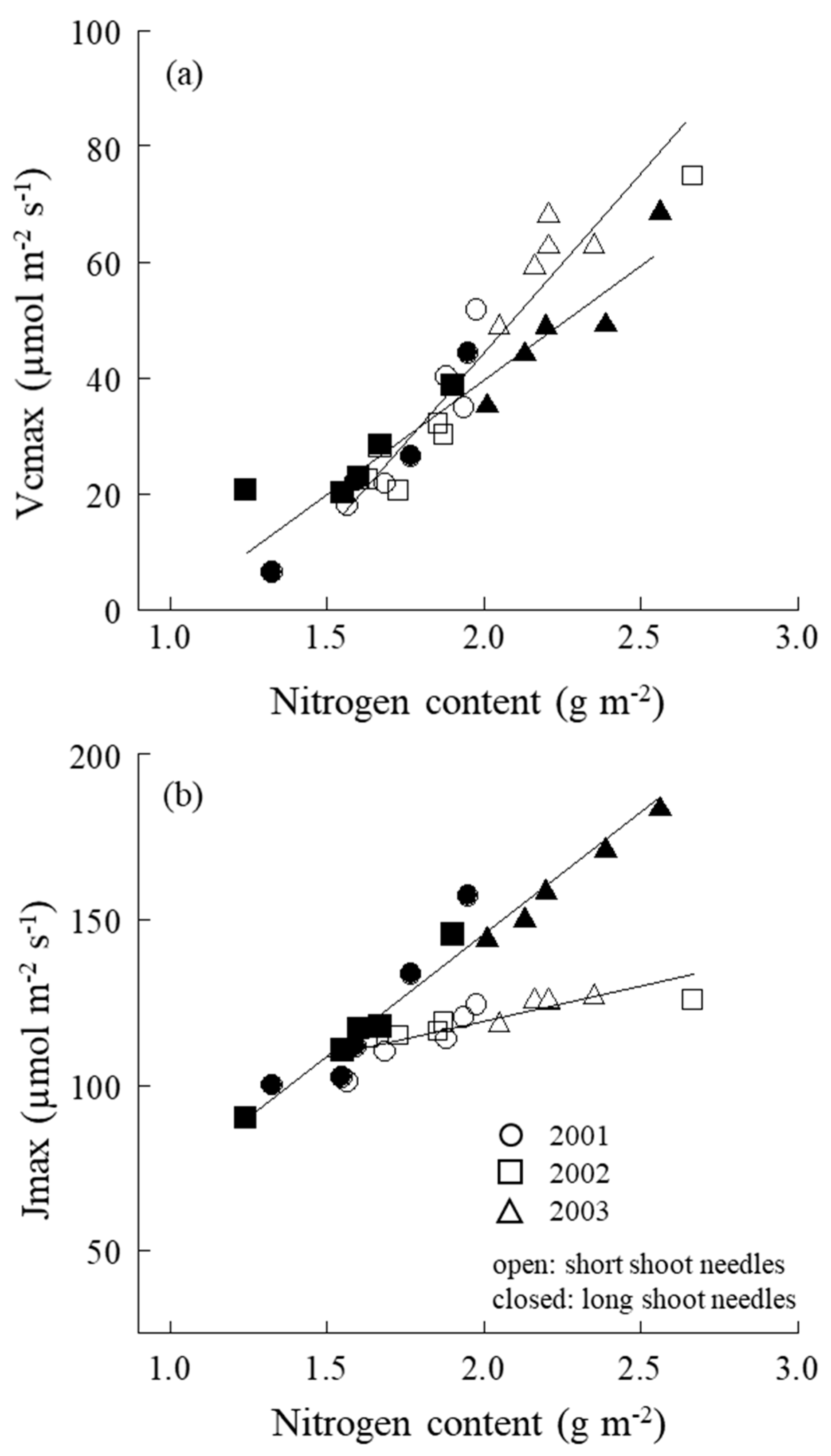
2.5. Vcmax, Jmax–N Relation
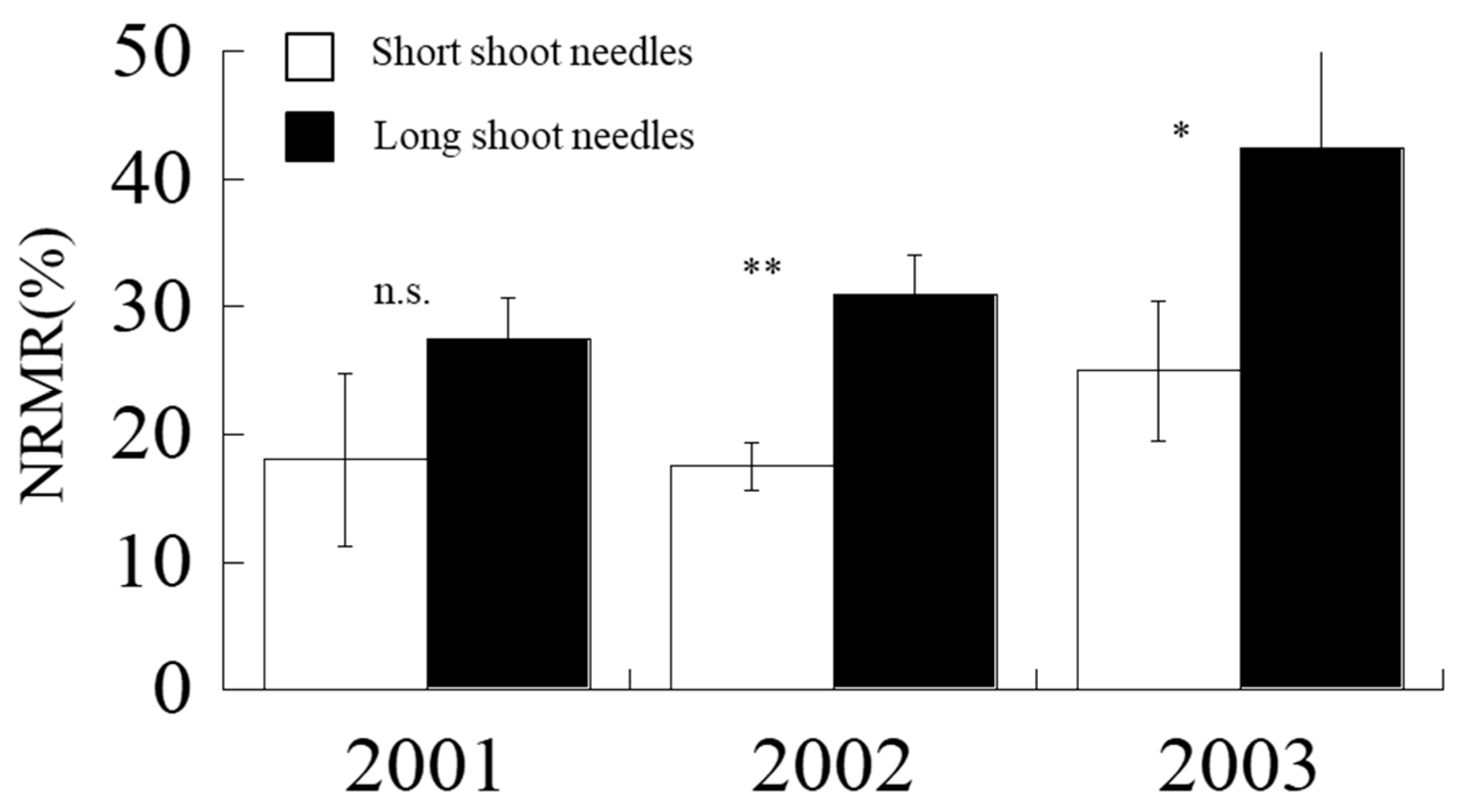
2.6. Nitrogen Remobilization Rate (NRMR)
3. Discussion
3.1. Seasonal and Yearly Variation in Photosynthetic Rate: Climatic Limitation
3.2. Differences in the Photosynthetic Nitrogen Relations Between Short and Long Shoots
4. Materials and Methods
4.1. Study Site
4.2. Plant Materials
4.3. Air Temperature and Soil Moisture Monitoring
4.4. Photosynthesis and Nitrogen Measurement of Needles
4.5. Analysis of A-Ci Curves
4.6. Calculation of Nitrogen Remobilization During Autumn Senescence
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lambers, H.; Chapin, F.; Stuart, I.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; ISBN 978-0-387-78340-6. [Google Scholar]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; ISBN 3-540-43516-6. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. (Eds.) Climate Change 2013: The Physical Science Basis; Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar] [CrossRef] [Green Version]
- Japan Meteorological Agency. Available online: http://www.jma.go.jp/jma/indexe.html (accessed on 17 May 2020).
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Chang. Biol. 2006, 12, 343–351. [Google Scholar] [CrossRef]
- Abrams, M.D.; Kubiske, M.E. Photosynthesis and water relations during drought in Acer rubrum L. genotypes from contrasting sites in central Pennsylvania. Funct. Ecol. 1990, 4, 727–733. [Google Scholar] [CrossRef]
- Kitaoka, S. Ecophysiological study on the growth and photosynthetic responses of deciduous broad-leaved tree seedlings invading into unmanaged larch [Larix leptolepis] plantations. Res. Bull. Hokkaido Univ. For. 2007, 64, 37–90. [Google Scholar]
- Kitaoka, S.; Koike, T. Seasonal and yearly variations in light use and nitrogen use by seedlings of four deciduous broad-leaved tree species invading larch plantations. Tree Physiol. 2005, 25, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Environment 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Kikuzawa, K.; Lechowicz, M.J. Ecology of Leaf Longevity; Ecological Research Monographs; Springer: Tokyo, Japan, 2011; ISBN 978-4-431-53917-9. [Google Scholar]
- Kozlowski, T.T.; Clausen, J.J. Shoot growth characteristics of heterophyllous woody plants. Can. J. Bot. 1966, 44, 827–843. [Google Scholar] [CrossRef]
- Matsuki, S.; Sano, Y.; Koike, T. Chemical and physical defence in early and late leaves in three heterophyllous birch species native to northern Japan. Ann. Bot. 2004, 93, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gower, S.T.; Richards, J.H. Larches: Deciduous conifers in an evergreen world. BioScience 1990, 40, 818–826. [Google Scholar] [CrossRef]
- Kitaoka, S.; Koike, T.; Quoreshi, A.M.; Takagi, K.; Wang, W.; Shi, F.; Kayama, M.; Ishida, N.; Mamiya, H.; Sasa, K. Seasonal changes in the photosynthetic capacity of Japanese larch trees planted on the Tomakomai National Forest, Northern Japan. Proc. Asia Flux Netw. 2001, 1, 109–112. [Google Scholar]
- Kurachi, N.; Hagihara, A.; Hozumi, K. Photosynthetic production in a Larix leptolepis plantation (IV). Photosynthetic characteristics of long and short shoot leaves. Trans. Jpn. Soc. Chubu Br. 1984, 32, 131–134. [Google Scholar]
- Kurachi, N. Distribution of Leaf—And Branch-Biomass Density within a Crown of Japanese Larch and its Relationship to Primary Production: Analysis by Sainome Cutting. In Crown and Canopy Structure in Relation to Productivity; Fujimori, T., Whitehead, D., Eds.; Forestry and Forest Products Research Institute: Tsukuba, Japan, 1986; pp. 308–322. [Google Scholar]
- Dörken, V.M. The Evolutionary Relevance of Vegetative Long-Shoot/Short-Shoot Differentiation in Gymnospermous Tree Species. Available online: https://www.schweizerbart.de/publications/detail/isbn/9783510480326/The_evolutionary_relevance_of_vegetative_long_shoot_short_shoot_differentiation_in_gymnospermous_tree_species (accessed on 18 May 2020).
- Takenaka, A. Structural variation in current-year shoots of broad-leaved evergreen tree saplings under forest canopies in warm temperate Japan. Tree Physiol. 1997, 17, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, T.; Kitaoka, S.; Ichie, T.; Lei, T.T.; Kitao, M. Photosynthetic characteristics of mixed deciduous-broadleaf forests from leaf to stand. Glob. Environ. Chang. Ocean Land Tokyo Terrapub 2004, 453–472. [Google Scholar]
- Koike, T.; Yazaki, K.; Eguchi, N.; Kitaoka, S.; Funada, R. Effects of elevated CO2 on ecophysiological responses of larch species native to northeast Eurasia. In Permafrost Ecosystems: Siberian Larch Forests; Ecological Studies (Analysis and Synthesis); Osawa, A., Zyryanova, O.A., Matsuura, Y., Kajimoto, T., Wein, R., Eds.; Springer: Dordrecht, The Netherland, 2010; Volume 209, pp. 447–456. ISBN 978-1-4020-9692-1. [Google Scholar]
- Nelson, G.C.; Valin, H.; Sands, R.D.; Havlík, P.; Ahammad, H.; Deryng, D.; Elliott, J.; Fujimori, S.; Hasegawa, T.; Heyhoe, E.; et al. Climate change effects on agriculture: Economic responses to biophysical shocks. Proc. Natl. Acad. Sci. USA 2014, 111, 3274–3279. [Google Scholar] [CrossRef] [Green Version]
- Hoshika, Y.; Paoletti, E.; Agathokleous, E.; Sugai, T.; Koike, T. Developing ozone risk assessment for larch species. Front. For. Glob. Chang. 2020, 3. [Google Scholar] [CrossRef]
- Fry, D.J.; Phillips, I.D.J. Photosynthesis of conifers in relation to annual growth cycles and dry matter production: I. some C4 characteristics in photosynthesis of Japanese larch (Larix leptolepis). Physiol. Plant. 1976, 37, 185–190. [Google Scholar] [CrossRef]
- Kurachi, N.; Hagihara, A.; Hozumi, K. Canopy photosynthetic production in a Japanese larch stand. I. Seasonal and vertical changes of leaf characteristics along the light gradient in a canopy. Ecol. Res. 1992, 7, 255–265. [Google Scholar] [CrossRef]
- Matyssek, R. Carbon, water and nitrogen relations in evergreen and deciduous conifers. Tree Physiol. 1986, 2, 177–187. [Google Scholar] [CrossRef]
- Matyssek, R.; Schulze, E.-D. Heterosis in hybrid larch (Larix decidua x leptolepis) I. The role of leaf characteristics. Trees 1987, 1, 219–224. [Google Scholar] [CrossRef]
- Matyssek, R.; Schulze, E.-D. Heterosis in hybrid larch (Larix decidua x leptolepis) II. Growth characteristics. Trees 1987, 1, 225–231. [Google Scholar] [CrossRef]
- Richards, J.H.; Teeri, J.A. Re-evaluation of proposed C4 photosynthetic characteristics in the genus Larix. Physiol. Plant. 1982, 55, 117–120. [Google Scholar] [CrossRef]
- Hozumi, K.; Kurachi, N. Estimation of seasonal changes in translocation rates in leaves of a Japanese larch stand. Bot. Mag. Shokubutsu-Gaku-Zasshi 1991, 104, 25–36. [Google Scholar] [CrossRef]
- Fujimoto, S. Studies on the shoot formation in Larix leptolepis Gordon. Res. Bull. Hokkaido Univ. For. Jpn. 1978, 35, 1–28. [Google Scholar]
- Wang, W.; Zu, Y.; Wang, H.; Matsuura, Y.; Sasa, K.; Koike, T. Plant biomass and productivity of Larix gmelinii forest ecosystems in Northeast China: Intra- and inter-species comparison. Eurasian J. For. Res. 2005, 8, 21–41. [Google Scholar]
- Ellsworth, D.S. Seasonal CO2 assimilation and stomatal limitations in a Pinus taeda canopy. Tree Physiol. 2000, 20, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M. Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats. Funct. Ecol. 2001, 15, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Westoby, M. Leaves at low versus high rainfall: Coordination of structure, lifespan and physiology. New Phytol. 2002, 155, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Bassow, S.L.; Bazzaz, F.A. Intra- and inter-specific variation in canopy photosynthesis in a mixed deciduous forest. Oecologia 1997, 109, 507–515. [Google Scholar] [CrossRef]
- Koike, T. Leaf structure and photosynthetic performance as related to the forest succession of deciduous broad-leaved trees. Plant Species Biol. 1988, 3, 77–87. [Google Scholar] [CrossRef]
- Nobel, P.S.; Zaragoza, L.J.; Smith, W.K. Relation between mesophyll surface area, photosynthetic rate, and illumination level during development for leaves of Plectranthus parviflorus Henckel. Plant Physiol. 1975, 55, 1067–1070. [Google Scholar] [CrossRef] [Green Version]
- Terashima, I.; Miyazawa, S.-I.; Hanba, Y.T. Why are sun leaves thicker than shade leaves?—Consideration based on analyses of CO2 diffusion in the leaf. J. Plant Res. 2001, 114, 93–105. [Google Scholar] [CrossRef]
- Evans, J.R. Leaf anatomy enables more equal access to light and CO2 between chloroplasts. New Phytol. 1999, 143, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Adams, W.W., III; Terashima, I. (Eds.) The Leaf: A Platform for Performing Photosynthesis; Advances in photosynthesis and respiration; Springer International Publishing: New York, NY, USA, 2018; ISBN 978-3-319-93592-8. [Google Scholar]
- Evans, J.R.; von Caemmerer, S. Carbon dioxide diffusion inside leaves. Plant Physiol. 1996, 110, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Manter, D.K.; Kerrigan, J. A/C(i) curve analysis across a range of woody plant species: Influence of regression analysis parameters and mesophyll conductance. J. Exp. Bot. 2004, 55, 2581–2588. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B.; Mooney, H.A. The photosynthesis-nitrogen relationship in wild plants. In On the Economy of Plant Form and Function; Givnish, T.J., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 25–55. ISBN 978-0-521-02249-1. [Google Scholar]
- Kitaoka, S.; Koike, T. Invasion of broad-leaf tree species into a larch plantation: Seasonal light environment, photosynthesis and nitrogen allocation. Physiol. Plant. 2004, 121, 604–611. [Google Scholar] [CrossRef]
- Koike, T.; Miyashita, N.; Toda, H. Effects of shading on leaf structural characteristics in successional deciduous broadleaved tree seedlings and their silvicultural meaning. For. Resour. Environ. Tokyo Univ. Agric. Technol. 1997, 35, 9–25. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Marchi, S.; Tognetti, R.; Minnocci, A.; Borghi, M.; Sebastiani, L. Variation in mesophyll anatomy and photosynthetic capacity during leaf development in a deciduous mesophyte fruit tree (Prunus persica) and an evergreen sclerophyllous Mediterranean shrub (Olea europaea). Trees 2008, 22, 559. [Google Scholar] [CrossRef]
- Hikosaka, K.; Nagamatsu, D.; Ishii, H.S.; Hirose, T. Photosynthesis-nitrogen relationships in species at different altitudes on Mount Kinabalu, Malaysia. Ecol. Res. 2002, 17, 305–313. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Kloeppel, B.D.; Ellsworth, D.S. Different photosynthesis-nitrogen relations in deciduous hardwood and evergreen coniferous tree species. Oecologia 1995, 104, 24–30. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Tenhunen, J.D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant Cell Environ. 1997, 20, 845–866. [Google Scholar] [CrossRef]
- Niinemets, Ü. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 2002, 22, 515–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Hikosaka, K.; Hanba, Y.T.; Hirose, T.; Terashima, I. Photosynthetic nitrogen-use efficiency in leaves of woody and herbaceous species. Funct. Ecol. 1998, 12, 896–905. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Yoneda, R.; Kitahashi, Y.; Watanabe, Y.; Ninomiya, I.; Koike, T. Interspecific variation of photosynthesis and leaf characteristics in canopy trees of five species of Dipterocarpaceae in a tropical rain forest. Tree Physiol. 2004, 24, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Onoda, Y.; Hirose, T.; Hikosaka, K. Effect of elevated CO2 levels on leaf starch, nitrogen and photosynthesis of plants growing at three natural CO2 springs in Japan. Ecol. Res. 2007, 22, 475–484. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Watanabe, M.; Watanabe, Y.; Kitaoka, S.; Utsugi, H.; Kita, K.; Koike, T. Growth and photosynthetic traits of hybrid larch F1 (Larix gmelinii var. japonica x L. kaempferi) under elevated CO2 concentration with low nutrient availability. Tree Physiol. 2011, 31, 965–975. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Kedrowski, R.A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Herrick, J.D.; Thomas, R.B. Leaf senescence and late-season net photosynthesis of sun and shade leaves of overstory sweetgum (Liquidambar styraciflua) grown in elevated and ambient carbon dioxide concentrations. Tree Physiol. 2003, 23, 109–118. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitaoka, S.; Laiye, Q.; Watanabe, Y.; Watanabe, M.; Watanabe, T.; Koike, T. Heterophyllous Shoots of Japanese Larch Trees: The Seasonal and Yearly Variation in CO2 Assimilation Capacity of the Canopy Top with Changing Environment. Plants 2020, 9, 1278. https://doi.org/10.3390/plants9101278
Kitaoka S, Laiye Q, Watanabe Y, Watanabe M, Watanabe T, Koike T. Heterophyllous Shoots of Japanese Larch Trees: The Seasonal and Yearly Variation in CO2 Assimilation Capacity of the Canopy Top with Changing Environment. Plants. 2020; 9(10):1278. https://doi.org/10.3390/plants9101278
Chicago/Turabian StyleKitaoka, Satoshi, Qu Laiye, Yoko Watanabe, Makoto Watanabe, Toshihiro Watanabe, and Takayoshi Koike. 2020. "Heterophyllous Shoots of Japanese Larch Trees: The Seasonal and Yearly Variation in CO2 Assimilation Capacity of the Canopy Top with Changing Environment" Plants 9, no. 10: 1278. https://doi.org/10.3390/plants9101278