Antitumor Potential and Phytochemical Profile of Plants from Sardinia (Italy), a Hotspot for Biodiversity in the Mediterranean Basin

 , , , ,
, , , ,
Abstract
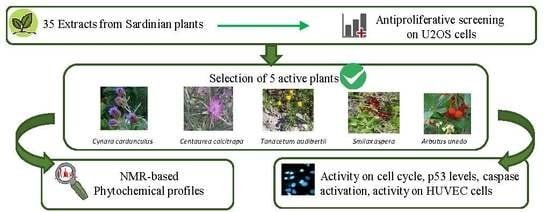
:
1. Introduction
2. Results and Discussion
2.1. Plant Traditional Uses and Screening of Antiproliferative Effect
2.2. Effect of the Selected Plants on the Cell Cycle Progression
2.3. Assessment of Apoptotic Effect of Selected Extracts
2.4. Comparison of the Selected Plants’ Effects on U2OS and HUVEC Cells
2.5. Phytochemical Analyses
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Preparation of Plant Extracts for Bioactivity Tests and for 1H NMR Profiling
3.4. Fractionation and Cynaropicrin Identification
3.5. NMR Measurement and Analysis
3.6. Phenolic and Flavonoid Content
3.7. Cell Culture and Treatment
3.8. MTT Assay
3.9. Cell Cycle Analysis
3.10. Analysis of p53 Expression Levels by Flow Cytometry
3.11. Caspase Activity Assay
3.12. Fluorescence Microscopy
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1H (δ; J) | 13C (δ) | HMBC | COSY |
|---|---|---|---|---|
| 1 | 3.00 (ddd; J = 11.53; 10.65 Hz; 6.84) | 46.22 | 39.78; 51.32; 74.18; 118.11; 142.84 | 2.09; 1.74; 2.88 |
| 2a | 2.09 (ddd; J = 13.14; 7.50; 6.84 Hz) | 39.78 | 46.22; 51.32; 74.18; 142.84 | 1.74; 3.00; 4.49 |
| 2b | 1.74 (ddd; J = 13.14; 11.53; 8.80 Hz) | 39.78 | 46.22; 74.18; 152.92 | 2.09; 3.00; 4.49 |
| 3 | 4.49 (dd; J = 8.80; 7.60 Hz) | 74.18 | 152.92 | 2.09; 1.74; 5.43; 5.64 |
| 4 | - | 152.92 | - | - |
| 5 | 2.88 (ddd; J = 10.65; 9.21 Hz) | 51.32 | 79.81 | 3.00; 4.33; 4.91; 5.33 |
| 6 | 4.33 (dd; J = 11.06; 8.89 Hz) | 79.81 | 75.15 | 2.88; 3.27 |
| 7 | 3.27 (m) | 41.48 | ov. | 4.33; 5.15; 6.12; 5.64 |
| 8 | 5.15 (m) | 75.15 | ov. | 3.27 |
| 9a | 2.73 (dd; J = 14.65; 5.26 Hz) | 37.14 | 118.11; 142.84 | 2.40; 4.91; 5.15 |
| 9b | 2.40 (dd; J = 14.65; 3.50 Hz) | 37.14 | 118.11; 142.84 | 2.73; 4.91; 5.15 |
| 10 | - | 142.84 | - | - |
| 11 | - | 138.77 | - | - |
| 12 | - | 170.01 | - | - |
| 13a | 6.12 (d; J = 3.30 Hz) | 121.96 | 41.48; 138.77; 170.01 | 3.27; 5.64 |
| 13b | 5.64 (d; J = 3.30 Hz) | 121.96 | 41.48; 138.77; 170.01 | 3.27; 6.12 |
| 14a | 5.14 (d; J = 2.09 Hz) | 118.11 | 37.14; 46.22; 142.84 | 2.40; 2.73; 4.91 |
| 14b | 4.91 (d; J = 2.09 Hz) | 118.11 | 37.14; 46.22; 142.84 | 2.40; 2.73; 5.14 |
| 15a | 5.43 (d; J = 1.97 Hz) | 122.36 | 51.32; 74.18; 152.92 | 2.88; 4.49; 5.33 |
| 15b | 5.33 (d; J = 1.97 Hz) | 122.36 | 51.32; 74.18; 152.92 | 2.88; 4.49; 5.43 |
| 1’ | - | 165.28 | - | - |
| 2’ | - | 140.80 | - | - |
| 3’a | 6.30 (d; J = 1.40 Hz) | 125.70 | 61.73; 140.80; 165.28 | 4.30; 5.97 |
| 3’b | 5.97 (d; J = 1.40 Hz) | 125.70 | 61.73; 140.80; 165.28 | 4.30; 6.30 |
| 4’ | 4.30 (s) | 61.73 | 125.70;140.80; 165.28 | 5.97; 6.30 |
References
- Bolzani da Silva, V.; Valli, M.; Pivatto, M.; Viegas, C. Natural products from Brazilian biodiversity as a source of new models for medicinal chemistry. Pure Appl. Chem. 2012, 84, 1837–1846. [Google Scholar] [CrossRef]
- Henrich, C.J.; Beutler, J.A. Matching the power of high throughput screening to the chemical diversity of natural products. Nat. Prod. Rep. 2013, 30, 1284–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cañadas, E.; Giuseppe, F.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Cañadas, E.M.; Bacchetta, G. Disentangling the influence of environmental and anthropogenic factors on the distribution of endemic vascular plants in Sardinia. PLoS ONE 2017, 12, e0182539. [Google Scholar] [CrossRef] [Green Version]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). Syst. Biodivers. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Cagno, V.; Sgorbini, B.; Sanna, C.; Cagliero, C.; Ballero, M.; Civra, A.; Donalisio, M.; Bicchi, C.; Lembo, D.; Rubiolo, P. In vitro anti-herpes simplex virus-2 activity of Salvia desoleana Atzei & V. Picci essential oil. PLoS ONE 2017, 12, e0172322. [Google Scholar] [CrossRef] [Green Version]
- Mandrone, M.; Bonvicini, F.; Lianza, M.; Sanna, C.; Maxia, A.; Gentilomi, G.A.; Poli, F. Sardinian plants with antimicrobial potential. Biological screening with multivariate data treatment of thirty-six extracts. Ind. Crop. Prod. 2019, 137, 557–565. [Google Scholar] [CrossRef]
- Sanna, C.; Scognamiglio, M.; Fiorentino, A.; Corona, A.; Graziani, V.; Caredda, A.; Cortis, P.; Montisci, M.; Ceresola, E.; Canducci, F.; et al. Prenylated phloroglucinols from Hypericum scruglii, an endemic species of Sardinia (Italy), as new dual HIV-1 inhibitors effective on HIV-1 replication. PLoS ONE 2018, 13, e0195168. [Google Scholar] [CrossRef] [Green Version]
- Sanna, C.; Rigano, D.; Corona, A.; Piano, D.; Formisano, C.; Farci, D.; Franzini, G.; Ballero, M.; Chianese, G.; Tramontano, E.; et al. Dual HIV-1 reverse transcriptase and integrase inhibitors from Limonium morisianum Arrigoni, an endemic species of Sardinia (Italy). Nat. Prod. Res. 2019, 33, 1798–1803. [Google Scholar] [CrossRef]
- Roy, A.; Ahuja, S.; Bharadvaja, N. A review on medicinal plants against cancer. J. Plant. Sci. Agric. Res. 2017, 2, 1008. [Google Scholar]
- Moten, A.; Schafer, D.; Ferrari, M. Redefining global health priorities: Improving cancer care in developing settings. J. Glob. Health 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuazo-Gaztelu, I.; Casanovas, O. Unraveling the role of angiogenesis in cancer ecosystems. Front. Oncol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.R.; Zhou, W.; Zhang, H.M.; Guo, Q.S.; Yang, W.; Li, B.J.; Sun, Z.H.; Gao, S.H.; Cui, R.J. Modulation of multiple signaling pathways of the plant-derived natural products in cancer. Front. Oncol. 2019, 9, 1153. [Google Scholar] [CrossRef] [Green Version]
- Dutta, R.; Khalil, R.; Green, R.; Mohapatra, S.S.; Mohapatra, S. Withania somnifera (Ashwagandha) and withaferin A: Potential in integrative oncology. Int. J. Mol. Sci. 2019, 20, 5310. [Google Scholar] [CrossRef] [Green Version]
- Remadevi, V.; Mohan Lathika, L.; Sasikumar Sujatha, A.; Sreeharshan, S. Ficus extract—A promising agent for antimammary tumorigenesis: A review on current status and future possibilities. Phytother. Res. 2019, 33, 1597–1603. [Google Scholar] [CrossRef]
- Chen, P.; Li, C.; Li, X.; Li, J.; Chu, R.; Wang, H. Higher dietary folate intake reduces the breast cancer risk: A systematic review and meta-analysis. Br. J. Cancer 2014, 110, 2327–2338. [Google Scholar] [CrossRef]
- Toledo, E.; Salas-Salvadó, J.; Donat-Vargas, C.; Buil-Cosiales, P.; Estruch, R.; Ros, E.; Corella, D.; Fitó, M.; Hu, F.B.; Arós, F.; et al. Mediterranean diet and invasive breast cancer risk among women at high cardiovascular risk in the PREDIMED Trial: A randomized clinical trial. JAMA Intern. Med. 2015, 175, 1752–1760. [Google Scholar] [CrossRef]
- Crew, K.D.; Brown, P.; Greenlee, H.; Bevers, T.B.; Arun, B.; Hudis, C.; McArthur, H.L.; Chang, J.; Rimawi, M.; Vornik, L.; et al. Phase IB randomized, double-blinded, placebo-controlled, dose escalation study of polyphenon E in women with hormone receptor-negative breast cancer. Cancer Prev. Res. (Phila) 2012, 5, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Hui, C.; Qi, X.; Qianyong, Z.; Xiaoli, P.; Jundong, Z.; Mantian, M. Flavonoids, flavonoid subclasses and breast cancer risk: A meta-analysis of epidemiologic studies. PLoS ONE 2013, 8, e54318. [Google Scholar] [CrossRef] [PubMed]
- Ballero, M.; Floris, R.; Poli, F. Le piante utilizzate nella medicina popolare nel territorio di Laconi (Sardegna centrale). Boll. Soc. Sarda Sci. Nat. 1997, 31, 207–229. [Google Scholar]
- Loi, M.C.; Maxia, L.; Maxia, A. Ethnobotanical Comparison between the Villages of Escolca and Lotzorai (Sardinia, Italy). J. Herbs Spices Med. Plants 2005, 11, 67–84. [Google Scholar] [CrossRef]
- Ballero, M.; Poli, F.; Sacchetti, G.; Loi, M.C. Ethnobotanical research in the territory of Fluminimaggiore (south-western Sardinia). Fitoterapia 2001, 72, 788–801. [Google Scholar] [CrossRef]
- Ballero, M.; Sacchetti, G.; Poli, F. Plants in folk medicine in the territory of Perdasdefogu (Central Sardinia, Italy). Allonia 1997, 35, 57–164. [Google Scholar]
- Bruni, A.; Ballero, M.; Poli, F. Quantitative ethnopharmacological study of the Campidano Valley and Urzulei district, Sardinia, Italy. J. Ethnopharmacol. 1997, 57, 97–124. [Google Scholar] [CrossRef]
- Loi, M.C.; Frailis, L.; Maxia, A. Le piante utilizzate nella medicina popolare nel territorio di Gesturi (Sardegna centro-meridionale). Atti. Soc. Tosc. Sci. Nat. Mem. Ser. B 2002, 109, 167–176. [Google Scholar]
- Loi, M.C.; Poli, F.; Sacchetti, G.; Selenu, M.B.; Ballero, M. Ethnopharmacology of Ogliastra (Villagrande Strisaili, Sardinia, Italy). Fitoterapia 2004, 75, 277–295. [Google Scholar] [CrossRef]
- Leonti, M.; Casu, L.; Sanna, F.; Bonsignore, L. A comparison of medicinal plant use in Sardinia and Sicily-De Materia Medica revisited? J. Ethnopharmacol. 2009, 121, 255–267. [Google Scholar] [CrossRef]
- Ballero, M.; Bruni, A.; Sacchetti, G.; Poli, F. Le piante utilizzate nella medicina popolare nel comune di Tempio Pausania (Sardegna settentrionale). Acta Phytother. 1997, 1, 23–29. [Google Scholar]
- Ballero, M.; Poli, F. Plants used in folk medicine of Monteleone (Northern Sardinia). Fitoterapia 1998, 69, 52–64. [Google Scholar]
- Atzei, A.D. Le Piante Nella Tradizione Popolare Della Sardegna; Carlo Delfino: Sassari, Italy, 2003. [Google Scholar]
- Palmese, M.T.; Manganelli, R.E.; Tomei, P.E. An ethno-pharmacobotanical survey in the Sarrabus district (south-east Sardinia). Fitoterapia 2001, 72, 619–643. [Google Scholar] [CrossRef]
- Hientz, K.; Mohr, A.; Bhakta-Guha, D.; Efferth, T. The role of p53 in cancer drug resistance and targeted chemotherapy. Oncotarget 2016, 8, 8921–8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Vu, T.; Cook, W.; Naseri, M.; Zhan, K.; Nakajima, W.; Harada, H. p53-independent Noxa induction by cisplatin is regulated by ATF3/ATF4 in head and neck squamous cell carcinoma cells. Mol. Oncol. 2018, 12, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Thakur, V.S.; Bhaskaran, N.; Nawab, A.; Babcook, M.A.; Jackson, M.W.; Gupta, S. Green Tea Polyphenols Induce p53-Dependent and p53- Independent Apoptosis in Prostate Cancer Cells through Two Distinct Mechanisms. PLoS ONE 2012, 7, e52572. [Google Scholar] [CrossRef] [PubMed]
- Tsang, W.P.; Chau, S.P.Y.; Kong, S.K.; Fung, K.P.; Kwok, T.T. Reactive oxygen species mediate doxorubicin induced p53-independent apoptosis. Life Sci. 2003, 73, 2047–2058. [Google Scholar] [CrossRef]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J. Fighting cancer by attacking its blood supply. Sci. Am. 1996, 275, 150–154. [Google Scholar] [CrossRef]
- Wu, J.-G.; Ma, L.; Lin, S.; Wu, Y.; Yi, J.; Yang, B.; Wu, J.-Z.; Wong, K. Anticancer and anti-angiogenic activities of extract from Actinidia eriantha Benth root. J. Ethnopharmacol. 2017, 203, 1–10. [Google Scholar] [CrossRef]
- Morgado, S.; Morgado, M.; Plácido, A.I.; Roque, F.; Duarte, A.P. Arbutus unedo L.: From traditional medicine to potential uses in modern pharmacotherapy. J. Ethnopharmacol. 2018, 225, 90–102. [Google Scholar] [CrossRef]
- Curti, V.; Di Lorenzo, A.; Dacrema, M.; Xiao, J.; Nabavi, S.M.; Daglia, M. In vitro polyphenol effects on apoptosis: An update of literature data. SeminCancer Biol. 2017, 46, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, D.; Zhang, Y.; Li, J.; Wu, Z.; Wang, Z.; Wang, D. Investigation of the pro-apoptotic effects of arbutin and its acetylated derivative on murine melanoma cells. Int. J. Mol. Med. 2018, 41, 1048–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.H.; van Spronsen, J.; Dai, Y.; Verberne, M.; Hollmann, F.; Arends, I.W.C.E.; Witkamp, G.-J.; Verpoorte, R. Are natural deep eutectic solvents the missing link in understanding cellular metabolism and physiology? Plant Physiol. 2011, 156, 1701–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepore, S.M.; Maggisano, V.; Lombardo, G.E.; Maiuolo, J.; Mollace, V.; Bulotta, S.; Russo, D.; Celano, M. Antiproliferative effects of cynaropicrin on anaplastic thyroid cancer cells. Endocr. Metab. Immune 2019, 19, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.; Bancheva, S.; Rosselli, S.; Maggio, A. Sesquiterpenoids in subtribe Centaureinae (Cass.) Dumort (tribe Cardueae, Asteraceae): Distribution, (13) C NMR spectral data and biological properties. Phytochemistry 2013, 95, 19–93. [Google Scholar] [CrossRef] [Green Version]
- Genovese, C.; Brundo, M.; Toscano, V.; Tibullo, D.; Puglisi, F.; Raccuia, S. Effect of Cynara extracts on multiple myeloma cell lines. Acta Hortic. 2016, 1147, 113–118. [Google Scholar] [CrossRef]
- Ramos, P.A.B.; Guerra, Â.R.; Guerreiro, O.; Santos, S.A.O.; Oliveira, H.; Freire, C.S.R.; Silvestre, A.J.D.; Duarte, M.F. Antiproliferative effects of Cynara cardunculus L. var. altilis (DC) lipophilic extracts. Int. J. Mol. Sci. 2016, 18, 63. [Google Scholar] [CrossRef]
- Erol-Dayi, Ö.; Pekmez, M.; Bona, M.; Aras-Perk, A.; Arda, N. Total phenolic contents, antioxidant activities cytotoxicity of three Centaurea species: C. calcitrapa subsp. calcitrapa, C. ptosimopappa, C. spicata. Free Radic. Antiox. 2011, 1, 31–36. [Google Scholar] [CrossRef]
- Ivanova, A.; Mikhova, B.; Batsalova, T.; Dzhambazov, B.; Kostova, I. New furostanol saponins from Smilax aspera L. and their in vitro cytotoxicity. Fitoterapia 2011, 82, 282–287. [Google Scholar] [CrossRef]
- Longo, L.; Vasapollo, G. Extraction and identification of anthocyanins from Smilax aspera L. berries. Food Chem. 2006, 94, 226–231. [Google Scholar] [CrossRef]
- Maxia, A.; Sanna, C.; Piras, A.; Porcedda, S.; Danilo, F.; Gonçalves, M.; Cavaleiro, C.; Salgueiro, L. Chemical composition and biological activity of Tanacetum audibertii (Req.) DC. (Asteraceae), an endemic species of Sardinia Island, Italy. Ind. Crop. Prod. 2015, 65, 472–476. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Mandrone, M.; Coqueiro, A.; Poli, F.; Antognoni, F.; Choi, Y.H. Identification of a collagenase-inhibiting flavonoid from Alchemilla vulgaris using NMR-based metabolomics. Planta Med. 2018, 84, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrone, M.; Scognamiglio, M.; Fiorentino, A.; Sanna, C.; Cornioli, L.; Antognoni, F.; Bonvicini, F.; Poli, F. Phytochemical profile and α-glucosidase inhibitory activity of Sardinian Hypericum scruglii and Hypericum Hircinum. Fitoter 2017, 120, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Mandrone, M.; Antognoni, F.; Aloisi, I.; Potente, G.; Poli, F.; Cai, G.; Faleri, C.; Parrotta, L.; Del Duca, S. Compatible and incompatible pollen-styles interaction in Pyrus communis L. show different transglutaminase features, polyamine pattern and metabolomics profiles. Front. Plant. Sci. 2019, 10, 741. [Google Scholar] [CrossRef] [PubMed]




| Plant Species | Plant Organ and Label | Ethnobotanical Use in Sardinia | Preparation |
|---|---|---|---|
| Arbutus unedo L. | Fruits (AuF) | Astringent [22,23], blood circulation (atherosclerosis) [24] | Decoction |
| wound healing [25,26] | Cataplasm | ||
| Leaves (AuL) | Inflammations of intestine, kidney, bladder [25] | Decoction (together with fruits and roots) | |
| Antipyretic, diarrhea [26,27,28], intestinal pains, as a vulnerary [29], diuretic, against cystitis and nephritis [22], asthma and bronchitis [28] | Decoction | ||
| Asphodelus ramosus L. subsp ramosus | Rhizome (ArRh) | Sore throat, skin diseases [24] | Decoction |
| Skin disease (chilblain) [24,29], hemorrhoids and impetigo [30] | Cataplasm | ||
| Leaves (ArL) | Diuretic (not recommended for patients affected by rheumatisms) [22] | Decoction | |
| Carlina gummifera (L.) Less. | Leaves (CgL) | Diuretic [27,30], cholagogue, stomachic, and diaphoretic [30] | Decoction and infusion |
| Centaurea calcitrapa L. | Aerial parts (CcA) | Antipyretic, digestive, for constipation and diarrhea [22,31] | Decoction |
| Antiseptic [31] | Juice | ||
| Centaurea horrida Badarò a | Aerial parts (ChA) | N.D. * | - |
| Centaurea napifolia L. | Aerial parts (CnA) | Nutritional purposes: aerial parts are often included in the diet [32] | Direct ingestion |
| Cistus monspeliensis L. | Aerial parts (CmA) | Topically for wound healing [30] | Poultice obtained by pressing fresh leaves between two pieces of wood |
| Cistus salvifolius L. | Aerial parts (CsA) | N.D. * | - |
| Cynara cardunculus L. | Aerial parts (CycA) | Hepatoprotective, blood depurative, hypocholesterolemic, digestive, intestinal spasmolytic [24,33] | Decoction |
| Liver diseases [28] | Infusion | ||
| Ferula arrigonii Bocchieri b | Leaves (FaL) | N.D. * | - |
| Roots (FaR) | N.D. * | - | |
| Galactites tomentosa Moench | Aerial parts (GtA) | Nutritional purposes: aerial parts are often included in the diet [32] | Direct ingestion |
| Genista corsica (Loisel.) D b | Aerial parts, flowers (GcA) | In Corse, flowers were used as disinfectant of wounds and abrasions [32] | Infusion |
| Glechoma sardoa (Bég.) Bég. b | Aerial parts (GsA) | Treatment of respiratory diseases, chronic catarrh, bronchitis, asthma, and to heal wounds [22] | Infusion of steam and flowers in water or milk |
| Hypericum hircinum L. ssp hircinum c | Aerial parts (HhA) | Burns and wounds healing [22] | Macerated in olive oil |
| For rheumatic and sciatic pains and for dislocations and sprains [22] | Macerated in olive oil and white wine, followed by evaporation of the wine | ||
| Hypericum scruglii Bacch., Brullo & Salmeri a | Aerial parts (HsA) | N.D. * | - |
| Lavandula stoechas L. | Aerial parts (LsA) | Against ringworm and skin diseases, and for wounds healing [22] | Macerated in spirit |
| Treatment of migraine, vertigo, asthma, palpitation, whooping cough, laryngitis, bronchitis, rheumatism [22,23], sedative, and skin diseases [24] | Infusion | ||
| Treatment of skin diseases [24] | Direct application of leaves | ||
| Limonium morisianum Arrigoni a | Aerial parts (LmA) | N.D. * | - |
| Myrtus communis L. | Fruits (McF) | Vulnerary, cough, sedative, digestive [21] | Decoction |
| Against cough and catarrh [25] and eupeptic [30] | Decoction (together with leaves) | ||
| Digestive [25] | Macerated in spirit | ||
| Leaves (McL) | Wound healing [30] | Dried and powdered for topical application | |
| Digestive and as an agent to treat respiratory ailments, as vulnerary, against hemorrhoids, to treat sweaty feet [22], catarrhal cough [26] | Infusion | ||
| Digestive, treatment of respiratory inflammations and hemorrhoids [22] | Fresh leaves pack | ||
| Vulnerary, cough, sedative, digestive [23], bronchitis, and asthma [28] | Decoction | ||
| Pistacia lentiscus L. | Fruits (PlF) | Cutaneous inflammations [30] | Fresh-squeezed and heated for topical application |
| Halitosis [22] | Fresh fruits | ||
| Catarrhal cough, gingivitis, sore throat [30], stomachache [28] | Decoction | ||
| Leaves (PlL) | Treatment of gingivitis, sore throat [30] | Decoction of fresh leaves to use as mouthwash | |
| Stomatitis, cough sedative, skin diseases [24] | Decoction | ||
| Against ticks [25] | Fumigation | ||
| Anticatarrhal [22], against cough and against bad breath and as an antisudorific [29] | Infusion | ||
| Pistacia terebinthus L. ssp. terebinthus | Leaves (PtL) | Catarrhal cough [26] | Decoction |
| Plagius flosculosus (L.) Alavi & Heywood b | Aerial parts (PfA) | N.D. * | - |
| Ptilostermon casabonae (L.) Greuter d | Aerial parts (PcA) | Antispasmodic [32] | Direct ingestion |
| Rosmarinus officinalis L. | Aerial parts (RoA) | Stomachache [30], cholagogue, general tonic, against common cold, hair loss [25], inappetence, digestive, diuretic, sedative, headache, pruritus [21] | Infusion |
| Hepatic [24], diarrhea [30], mucolytic, anti-inflammatory, tooth care, colic, tonic for blood pressure, joint pains [33], antitussive, antispasmodic, migraine, digestive [25], taenifuge, asthma, bronchitis; stomachic [28] | Decoction | ||
| Antirheumatic [28] | Cataplasm | ||
| Santolina corsica Jord. & Fourr b | Aerial parts (ScA) | N.D. * | |
| Scolymus hispanicus L. subsp. hispanicus | Aerial parts (ShA) | Nutritional purposes: young stems are often included in the diet [32] | Direct ingestion |
| Silybum marianum (L.) Gaertn. | Aerial parts (SmA) | Treatment of bleeding, diuretic, hypotensive, sudorifer in case of pneumonia and chronic catarrh [22] | Decoction (together with the seeds) |
| Smilax aspera L. | Aerial parts (SaA) | Treatment of rheumatisms, skin diseases [24], hemorrhoids [28] | Cataplasm |
| Sudorific and blood cleanser [24,27] | Decoction | ||
| Toothache [28] | Drops of fresh-squeezed juice applied on the gums | ||
| Stachys glutinosa L. c | Aerial parts (SgA) | Antiseptic, antispasmodic [32] | Infusion |
| Cholagogue, diuretic, and hepatoprotective [27], common cold [28] | Decoction | ||
| Tanacetum audibertii (Req.) DC b | Aerial parts (TaA) | Digestive, vermifuge, antiarthritic and to treat menstrual disorders [30] | Decoction |
| Thymus herba barona Loisel. e | Aerial parts (ThA) | Antitussive, expectorant, antispasmodic, collutory [22], anthelmintic, treatment of stomachache [24], sore throat, common cold, tonic and antianemic, diuretic [28] | Decoction or infusion |
| Against foot perspiration and urticaria [25] | Powder obtained by crushing aerial parts | ||
| Rheumatisms [24] | Cataplasm | ||
| Catarrhal, antipyretic [24] | Macerated in wine | ||
| Lung diseases [24] | Vaporization |
| Extracts | U2OS Cells | HUVEC Cells | ||
|---|---|---|---|---|
| 50 μg/mL | 100 μg/mL | 50 μg/mL | 100 μg/mL | |
| AuL | 79.6 ± 7.7 | 56 ± 5.5 | 111.8 ± 0.63 | 104.7 ± 4.6 |
| CcA | 90 ± 2 | 50 ± 6.5 | 6.5 ± 0.29 | 6.4 ± 0.05 |
| CycA | 45 ± 1.4 | 29 ± 9.9 | 6.5 ± 0.02 | 7.7 ± 0.37 |
| SaA | 85.4 ± 5.5 | 68.3 ± 0.7 | 114.2 ± 0.08 | 77.3 ± 1.11 |
| TaA | 96.9 ± 12.9 | 56.9 ± 3.9 | 98.9 ± 0.07 | 50.4 ± 1.5 |
| Metabolite | Diagnostic 1H NMR Signal (δ) Used for the Quantification and Number of Underlying Proton/s (in Brackets) | Metabolite Quantity in the Extract (μg/mg of Extract) | ||||
|---|---|---|---|---|---|---|
| AuL | CycA | CcA | TaA | SaA | ||
| alanine | 1.48 (3H) | 1.4 | 2.0 | - | 8.9 | 3.7 |
| arbutin | 7.07 (2H) | 75 | - | - | - | - |
| aspartate | 2.96 (1H) | - | - | 17 | 33 | - |
| caffeic acid | 7.62 (1H) | - | - | - | - | 13 |
| cynaropicrin | 6.16 (1H) | - | 159 | - | - | - |
| α-glucose | 5.2 (1H) | 22 | 24 | - | - | 40 |
| β-glucose | 4.59 (1H) | 36 | 49 | - | 34 | 88 |
| isoleucine | 1.06 (3H) | - | - | - | - | 19 |
| quinic acid | 1.87 (1H) | 142 | 43 | - | - | 193 |
| shikimic acid | 6.45 (1H) | - | - | - | - | 141 |
| sucrose | 5.4 (1H) | 138 | - | - | 45 | 79 |
| Plant Name | Family | Plant Organ and Sample Label in Brackets | GPS Coordinates | HARVESTING DATE | Voucher |
|---|---|---|---|---|---|
| Arbutus unedo L. | Ericaceae | Fruits (AuF) | 39°45′37.8″ N 9°30′31.0″ E | December 2017 | Herbarium CAG 878 |
| Leaves (AuL) | 39°45′37.8″ N 9°30′31.0″ E | December 2017 | |||
| Asphodelus ramosus L. subsp. ramosus | Asphodelaceae | Rhizome (ArRh) | 39°10′38.7″ N 9°22′50.3″ E | April 2017 | Herbarium CAG 1405 |
| Leaves (ArL) | 39°10′38.7″ N 9°22′50.3″ E | April 2017 | |||
| Carlina gummifera (L.) Less. | Asteraceae | Leaves (CgL) | 39°45′44.2″ N 9°40′16.9″ E | July 2018 | Herbarium CAG 770 |
| Centaurea calcitrapa L. | Asteraceae | Aerial parts (CcA) | 39°18′02.3″ N 8°53′39.4″ E | June 2017 | Herbarium CAG 781 |
| Centaurea horrida Badarò * | Asteraceae | Aerial parts (ChA) | 40°57′51.6″ N 8°12′05.0″ E | June 2017 | Herbarium CAG 777 |
| Centaurea napifolia L. | Asteraceae | Aerial parts (CnA) | 39°16′51.5″ N 8°56′01.5″ E | June 2017 | Herbarium CAG 784 |
| Cistus monspeliensis L. | Cistaceae | Aerial parts (CmA) | 39°45′44.2″ N 9°40′16.9″ E | April 2018 | Herbarium CAG 135 |
| Cistus salviifolius L. | Cistaceae | Aerial parts (CsA) | 39°45′44.2″ N 9°40′16.9″ E | April 2018 | Herbarium CAG 135/C |
| Cynara cardunculus L. | Asteraceae | Aerial parts (CycA) | 39°18′02.3″ N 8°53′39.4″ E | April 2017 | Herbarium CAG 790 |
| Ferula arrigonii Bocchieri ** | Apiaceae | Leaves (FaL) | 39°51′37.9″ N 8°26′05.2″ E | April 2017 | Herbarium CAG 612/A |
| Roots (FaR) | 39°51′37.9″ N 8°26′05.2″ E | April 2017 | |||
| Galactites tomentosa Moench | Asteraceae | Aerial parts (GtA) | 39°46′16.7″ N 9°30′41.6″ E | September 2018 | Herbarium CAG 789 |
| Genista corsica (Loisel.) DC ** | Fabaceae | Aerial parts (GcA) | 39°49′35.0″ N 9°20′27.5″ E | May 2017 | Herbarium CAG 286 |
| Glechoma sardoa (Bég.) Bég ** | Lamiaceae | Aerial parts (GsA) | 39°57′33.5″ N 9°19′13.3″ E | June 2017 | Herbarium CAG 1104 |
| Hypericum hircinum L. ssp hircinum+ | Hypericaceae | Aerial parts (HhA) | 39°46′55.8″ N 9°30′52.5″ E | June 2018 | Herbarium CAG 232 |
| Hypericum scruglii Bacch., Brullo & Salmeri * | Hypericaceae | Aerial parts (HsA) | 39°45′57.4″ N 9°30′41.8″ E | June 2018 | Herbarium CAG 239/C |
| Lavandula stoechas L. | Lamiaceae | Aerial parts (LsA) | 39°45′44.2″ N 9°40′16.9″ E | April 2017 | Herbarium CAG 1067 |
| Limonium morisianum Arrigoni * | Plumbaginaceae | Aerial parts (LmA) | 39°54′33.3″ N 9°24′41.0″ E | December 2017 | Herbarium CAG 909/G |
| Myrtus communis L. | Myrtaceae | Fruits (McF) | 39°45′44.2″ N 9°40′16.9″ E | December 2018 | Herbarium CAG 514 |
| Leaves (McL) | 39°08′22.2″ N 8°58′08.9″ E | April 2018 | |||
| Pistacia lentiscus L. | Anacardiaceae | Fruits (PlF) | 39°45′44.2″ N 9°40′16.9″ E | December 2017 | Herbarium CAG 280 |
| Leaves (PlL) | December 2017 | ||||
| Pistacia terebinthus L. ssp. terebinthus | Anacardiaceae | Leaves (PtL) | 39°47′38.8″ N 9°30′38.3″ E | June 2018 | Herbarium CAG 279 |
| Plagius flosculosus ( L.) Alavi & Heywood ** | Asteraceae | Aerial parts (PfA) | 39°21′45.2″ N 8°32′24.1″ E | July 2017 | Herbarium CAG 743 |
| Ptilostemon casabonae (L.) Greuter ++ | Asteraceae | Aerial parts (PcA) | 39°53′52.7″ N 9°26′31.8″ E | June 2018 | Herbarium CAG 796 |
| Rosmarinus officinalis L. | Lamiaceae | Aerial parts (RoA) | 40°34′10.1″ N 8°22′57.0″ E | May 2017 | Herbarium CAG 1091 |
| Santolina corsica Jord. & Fourr** | Asteraceae | Aerial parts (ScA) | 40°32′30.6″ N 9°36′09.4″ E | November 2017 | Herbarium CAG 732/A |
| Scolymus hispanicus L. subsp. hispanicus | Asteraceae | Aerial parts (ShA) | 39°03′25.9″ N 8°58′46.3″ E | June 2018 | Herbarium CAG 812 |
| Silybum marianum (L.) Gaertn. | Asteraceae | Aerial parts (SmA) | 39°16′51.5″ N 8°56′01.5″ E | May 2017 | Herbarium CAG 801 |
| Smilax aspera L. | Smilacaceae | Aerial parts (SaA) | 39°10′38.7″ N 9°22′50.3″ E | May 2017 | Herbarium CAG 1414 |
| Stachys glutinosa L.+ | Lamiaceae | Aerial parts (SgA) | 39°55′46.1″ N 9°27′10.7″ E | June 2017 | Herbarium CAG 1099 |
| Tanacetum audibertii (Req.) DC** | Asteraceae | Aerial parts (TaA) | 40°02′07.9″ N 9°17′59.1″ E | August 2018 | Herbarium CAG 737/A |
| Thymus herba barona Loisel § | Lamiaceae | Aerial parts (ThA) | 39°56′01.2″ N 9°19′56.9″ E | June 2017 | Herbarium CAG 1065 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappadone, C.; Mandrone, M.; Chiocchio, I.; Sanna, C.; Malucelli, E.; Bassi, V.; Picone, G.; Poli, F. Antitumor Potential and Phytochemical Profile of Plants from Sardinia (Italy), a Hotspot for Biodiversity in the Mediterranean Basin. Plants 2020, 9, 26. https://doi.org/10.3390/plants9010026
Cappadone C, Mandrone M, Chiocchio I, Sanna C, Malucelli E, Bassi V, Picone G, Poli F. Antitumor Potential and Phytochemical Profile of Plants from Sardinia (Italy), a Hotspot for Biodiversity in the Mediterranean Basin. Plants. 2020; 9(1):26. https://doi.org/10.3390/plants9010026
Chicago/Turabian StyleCappadone, Concettina, Manuela Mandrone, Ilaria Chiocchio, Cinzia Sanna, Emil Malucelli, Vincenza Bassi, Giovanna Picone, and Ferruccio Poli. 2020. "Antitumor Potential and Phytochemical Profile of Plants from Sardinia (Italy), a Hotspot for Biodiversity in the Mediterranean Basin" Plants 9, no. 1: 26. https://doi.org/10.3390/plants9010026