Temporal Distinction between Male and Female Floral Organ Development in Nicotiana tabacum cv. Xanthi (Solanaceae)


Abstract
:1. Introduction
2. Results
2.1. Corolla Growth Analysis
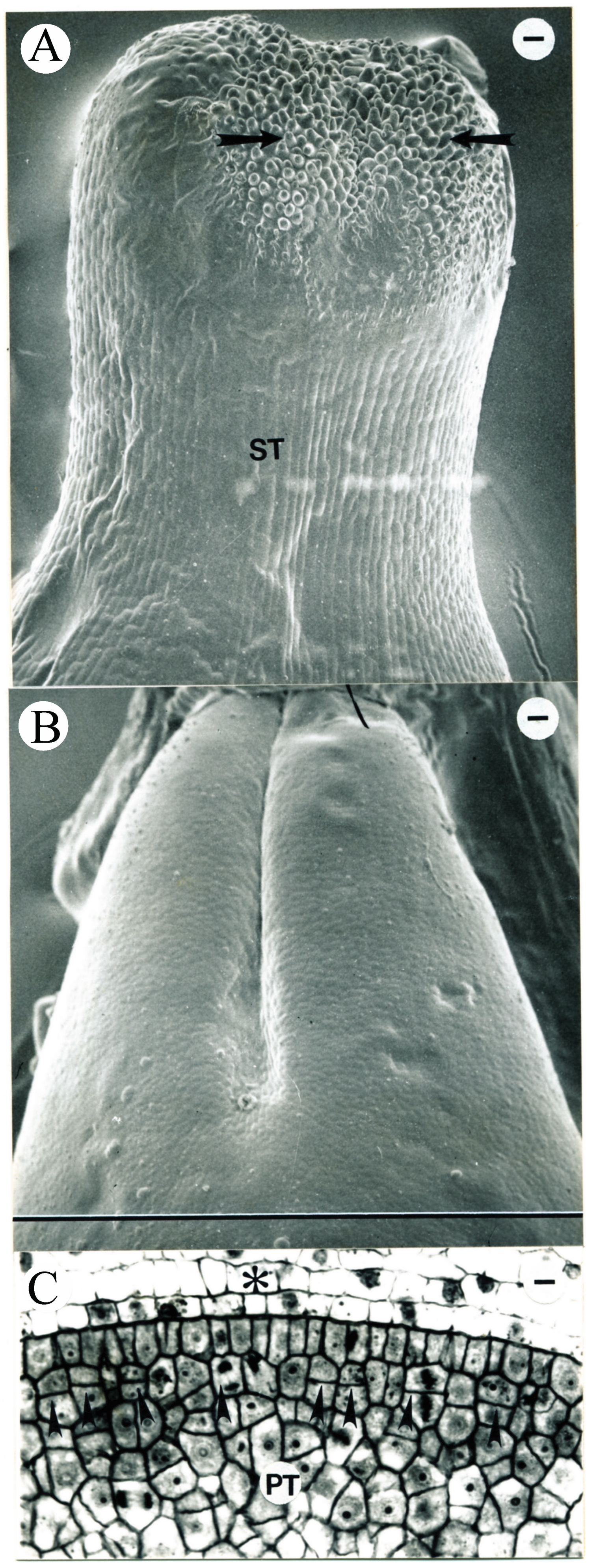
2.2. Pistil and Stamen Development
3. Discussion
3.1. Photoperiod Cycle Affects the Floral Initiation and Development
3.2. Unequal Duration of Male and Female Floral Organ Developmental Stages
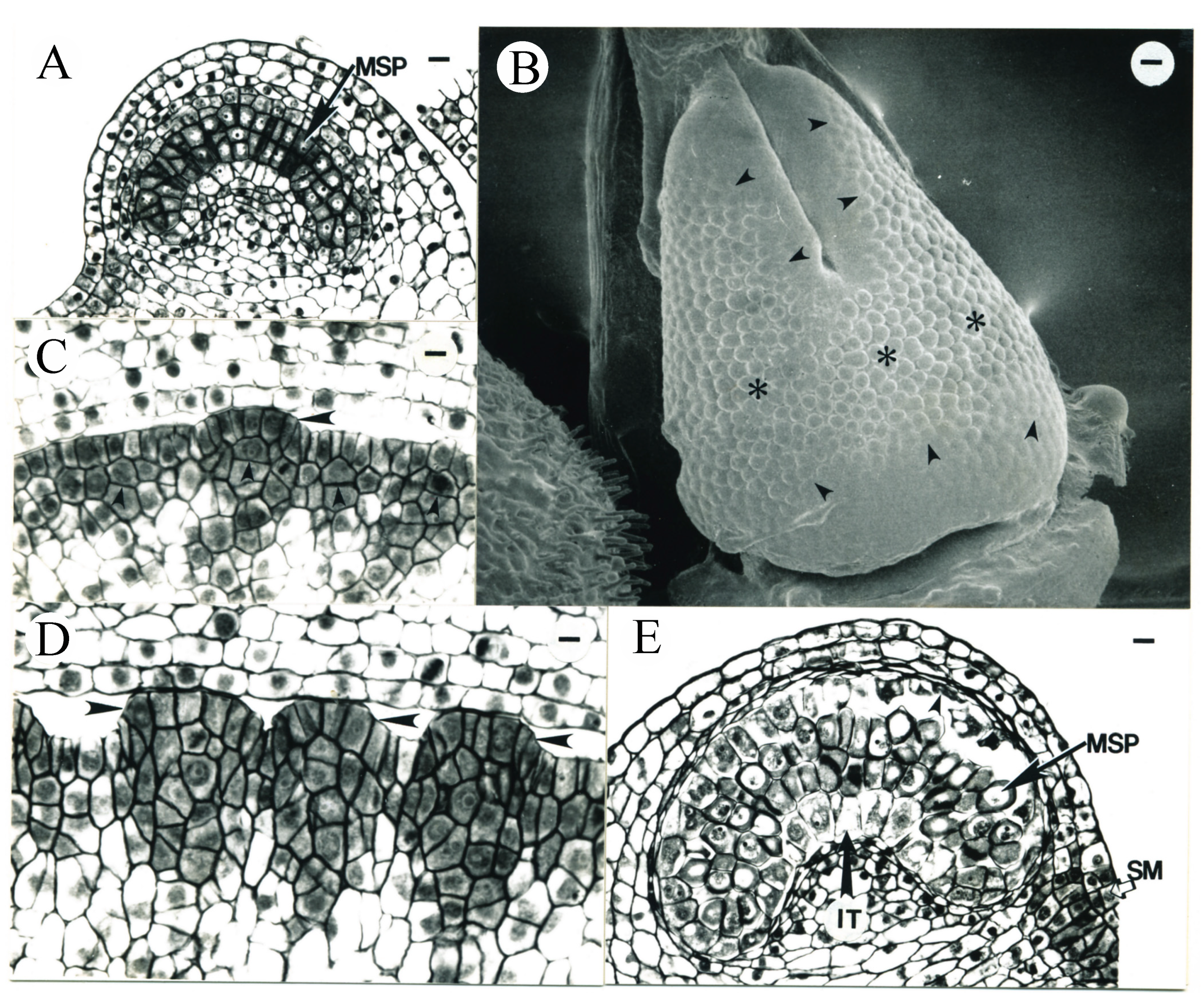
3.3. Carpel Development
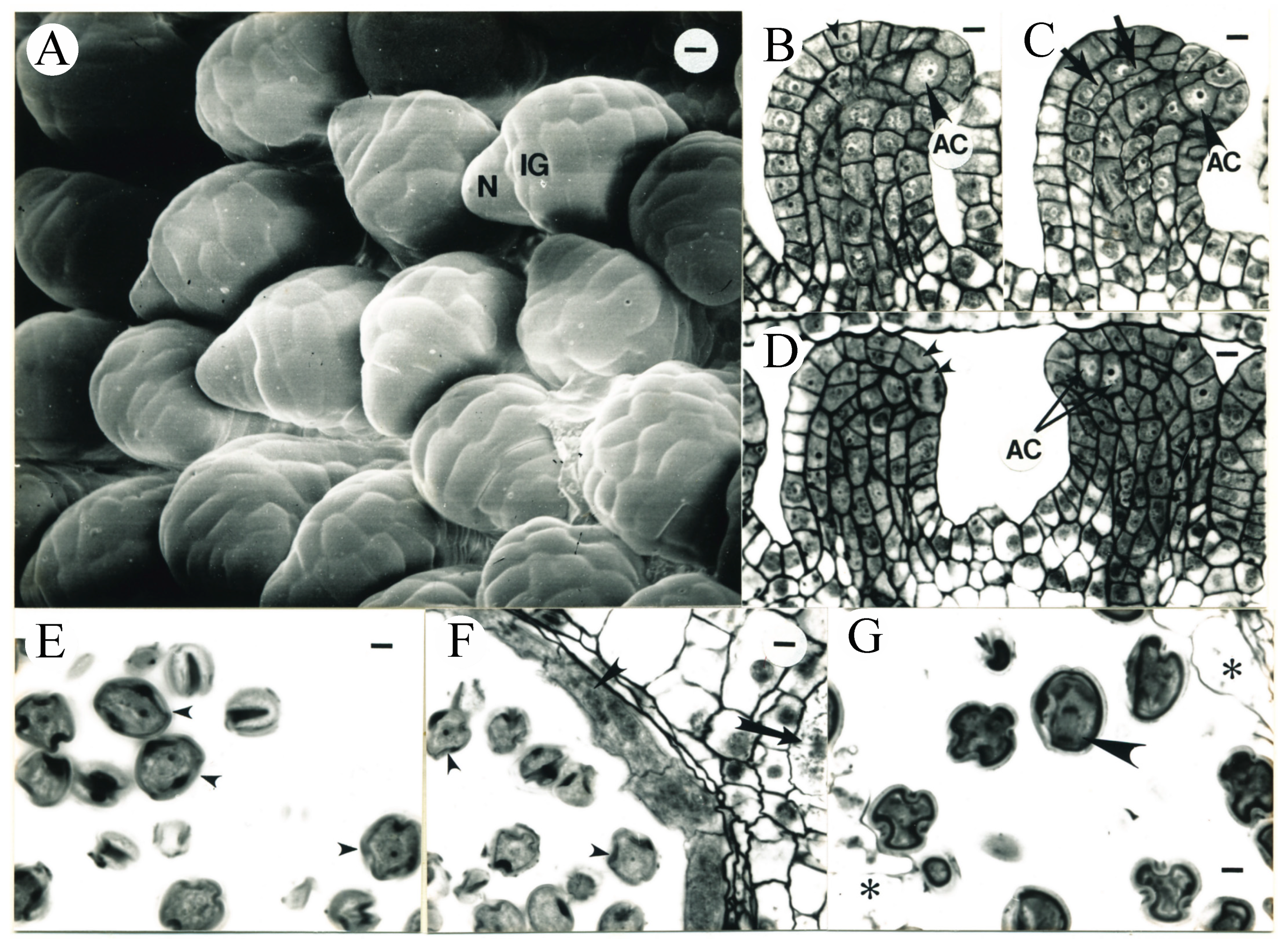
3.4. Placenta Development and Ovule Differentiation
3.5. Formation of Megasporocytes
3.6. Pistil and Stamen Development in Tobacco
4. Materials and Methods
4.1. Growth of Tobacco Plants
4.2. Determination of Inflorescence Plastochron
4.3. Microscopy and Histology
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lloyd, D.G.; Webb, C.J. The avoidance of interference between the presentation of pollen and stigmas in angiosperms. I. Dichogamy. N. Z. J. Bot. 1986, 24, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.J.; Lloyd, D.G. The avoidance of interference between the presentation of pollen and stigmas in angiosperms. II. Herkogamy. N. Z. J. Bot. 1986, 24, 163–178. [Google Scholar] [CrossRef]
- Hill, J.P.; Lord, E.M. A method for determining plastochron indices during heteroblastic shoot growth. Am. J. Bot. 1990, 77, 1491–1497. [Google Scholar] [CrossRef]
- Hill, J.P.; Lord, E.M.; Shaw, R.G. Morphological and growth rate differences among outcrossing and self-pollinating races of Arenaria uniflora (Caryophyllaceae). J. Evol. Biol. 1992, 5, 559–573. [Google Scholar] [CrossRef]
- Fenster, C.B.; Diggle, P.K.; Barrett, S.C.H.; Ritland, K. The genetics of floral development differentiating two species of Mimulus (Scrophulariaceae). Heredity 1995, 74, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.P.; Malmberg, R.L. Timing of morphological and histological development in premeiotic anthers of Nicotiana tabacum cv. Xanthi (Solanaceae). Am. J. Bot. 1996, 83, 285–295. [Google Scholar] [CrossRef]
- Runions, C.J.; Geber, M.A. Evolution of the self-pollinating flower in Clarkia xantiana (Onagraceae). I. Size and development of floral organs. Am. J. Bot. 2000, 87, 1439–1451. [Google Scholar] [CrossRef] [Green Version]
- Sherry, R.A.; Lord, E.M. A comparative developmental study of the selfing and outcrossing flowers of Clarkia tembloriensis (Onagraceae). Int. J. Plant Sci. 2000, 161, 563–574. [Google Scholar] [CrossRef]
- Lord, E.M. Cleistogamy: A tool for the study of floral morphogenesis, function and evolution. Bot. Rev. 1981, 47, 421–449. [Google Scholar] [CrossRef]
- Cohen, J.C.; Litt, A.; Davis, J.I. Comparative floral development in Lithospermum (Boraginaceae) and implications for the evolution and development of heterostyly. Am. J. Bot. 2012, 99, 797–805. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, A.; Li, W.; Jiang, Y.; Song, S.; Li, Y.; Chen, R. Comparative proteomic analysis of eggplant (Solanum melongena L.) heterostylous pistil development. PLoS ONE 2017, 12, e0179018. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.C.H. Mating strategies in flowering plants: The outcrossing-selfing paradigm and beyond. Philos. Trans. R. Soc. 2003, 358, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Dejong, T.J.; Waser, N.M.; Klinkhamer, P.G.L. Geitonogamy-the neglected side of selfing. Trends Ecol. Evol. 1993, 8, 321–325. [Google Scholar] [CrossRef]
- Zhong, L.; Barrett, S.C.H.; Wang, X.; Wu, Z.; Sun, H.; Li, D.; Wang, H.; Zhou, W. Phylogenomic analysis reveals multiple evolutionary origins of selfing from outcrossing in a lineage of heterostylous plants. New Phytol. 2019, 224, 1290–1303. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.C.H.; Charlesworth, D. Effects of a change in the level of inbreeding on the genetic load. Nature 1991, 352, 522–524. [Google Scholar] [CrossRef]
- Carr, D.E.; Dudash, M.R. Recent approaches into the genetic basis of inbreeding depression in plants. Philos. Trans. R. Soc. 2003, 358, 1071–1084. [Google Scholar] [CrossRef]
- Lande, R.; Schemske, D.W. The evolution of self-fertilization and inbreeding depression in plants. I. Genetic Models. Evolution 1985, 39, 24–40. [Google Scholar] [CrossRef]
- Willis, J.H. Inbreeding Load, Average dominance and the mutation rate for mildly deleterious alleles in Mimulus guttatus. Genetics 1999, 153, 1885–1898. [Google Scholar]
- Li, P.; Johnston, M.O. Heterochrony in plant evolutionary studies through the twentieth century. Bot. Rev. 2000, 66, 57–88. [Google Scholar] [CrossRef]
- Mazer, S.J.; Paz, H.; Bell, M.D. Life history, floral development and mating system in Clarkia xantiana (Onagraceae): Do floral and whole-plant rates of development evolve independently. Am. J. Bot. 2004, 91, 2041–2050. [Google Scholar] [CrossRef]
- Holtsford, T.P.; Ellstrand, N.C. Genetic and environmental variation in floral traits affecting outcrossing rate in Clarkia tembloriensis (Onagraceae). Evolution 1992, 46, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Mazer, S.J.; Delesalle, V.A. Floral trait variation in Spergularia marina (Caryophyllaceae): Ontogenetic, maternal family, and population effects. Heredity 1996, 77, 269–281. [Google Scholar] [CrossRef]
- Hill, J.P.; Lord, E.M. The role of developmental timing in the evolution of floral form. Semin. Dev. Biol. 1990, 1, 281–287. [Google Scholar]
- Alves, C.M.L.; Noyszewski, A.K.; Smith, A.G. Nicotiana tabacum pollen-pistil interactions show unexpected spatial and temporal differences in pollen tube growth among genotypes. Plant Reprod. 2019, 32, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Rocha, S.; Vázquez, N.; López-Fernández, H.; Fdez-Riverola, F.; Reboiro-Jato, M.; Vieira, C.P. Predicting specificities under the non-self gametophytic self-Incompatibility recognition model. Front. Plant Sci. 2019, 10, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, D.; Baraket, G.; Perez, V.; Ben Mustapha, S.; Salhi-Hannachi, A.; Hormaza, J.I. Analysis of self-incompatibility and genetic diversity in diploid and hexaploid plum genotypes. Front. Plant Sci. 2019, 10, 896. [Google Scholar] [CrossRef] [Green Version]
- Kamalay, J.C.; Goldberg, R.B. Regulation of structural gene expression in tobacco. Cell 1980, 19, 935–946. [Google Scholar] [CrossRef]
- Roberts, W.R.; Roalson, E.H. Comparative transcriptome analyses of flower development in four species of Achimenes (Gesneriaceae). BMC Genom. 2017, 18, 240. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, M.E. Changing MADS-Box transcription factor protein–protein interactions as a mechanism for generating floral morphological diversity. J. Integr. Comp. Biol. 2017, 57, 1312–1321. [Google Scholar] [CrossRef]
- Koltunow, A.M.; Truettner, J.; Cox, K.H.; Wallroth, M.; Goldberg, R.B. Different temporal and spatial gene expression patterns occur during anther development. Plant Cell 1990, 2, 1201–1224. [Google Scholar] [CrossRef]
- Erickson, R.O. Cytological and growth correlations in the flower bud and anther of Lilium longiflorum. Am. J. Bot. 1948, 35, 729–739. [Google Scholar] [CrossRef]
- Tanaka, I.; Taguchi, T.; Ito, M. Studies on microspore development in Liliaceous plants. I. The duration of the cell cycle and developmental aspects in lily microspores. Bot. Mag. 1979, 92, 291–298. [Google Scholar] [CrossRef]
- Erickson, R.O.; Michelini, F.J. The plastochron index. Am. J. Bot. 1957, 44, 297–305. [Google Scholar] [CrossRef]
- Poyatos-Pertíñez, S.; Quinet, M.; Ortíz-Atienza, A.; Yuste-Lisbona, F.J.; Pons, C.; Giménez, E.; Angosto, T.; Granell, A.; Capel, J.; Lozano, R. A factor linking floral organ identity and growth revealed by characterization of the tomato mutant unfinished flower development (ufd). Front. Plant Sci. 2016, 7, 1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelán-Muñoz, N.; Herrera, J.; Cajero-Sánchez, W.; Arrizubieta, M.; Trejo, C.; García-Ponce, B.; de la Paz Sánchez, M.; Álvarez-Buylla, E.R.; Garay-Arroyo, A. MADS-Box genes are key components of genetic regulatory networks involved in abiotic stress and plastic developmental responses in plants. Front. Plant Sci. 2019, 10, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meicenheimer, R.D. The plastochron index: Still useful after nearly six decades. Am. J. Bot. 2014, 101, 1821–1835. [Google Scholar] [CrossRef] [Green Version]
- Verbeke, J.A. Fusion events during floral morphogenesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 583–598. [Google Scholar] [CrossRef]
- Hill, J.P.; Malmberg, R.L. Rates of corolla growth in tobacco determined with the plastochron index. Planta 1991, 185, 472–478. [Google Scholar] [CrossRef]
- Quinby, J.R.; Hesketh, J.D.; Voigt, R.L. Influence of temperature and photoperiod on floral initiation and leaf number in Sorghum. Crop Sci. 1973, 13, 243–246. [Google Scholar] [CrossRef]
- Sandoval-Oliveros, R.; Guevara-Olvera, L.; Beltran, J.P.; Gomez-Mena, C.; Acosta-Garcıa, G. Developmental landmarks during floral ontogeny of jalapeno chili pepper (Capsicum annuum L.) and the effect of gibberellin on ovary growth. Plant Reprod. 2017, 30, 119–129. [Google Scholar] [CrossRef]
- Hicks, G.S. Initiation of floral organs in Nicotiana tabacum. Can. J. Bot. 1973, 51, 1611–1617. [Google Scholar] [CrossRef]
- Hill, J.P.; Lord, E.M. Floral development in Arabidopsis thaliana—A comparison of the wild type and the homeotic pistillata mutant. Can. J. Bot. 1989, 67, 2922–2936. [Google Scholar] [CrossRef]
- Crone, W.; Lord, E.M. Floral organ initiation and development in wild type Arabidopsis thaliana (Brassicaceae) and in the organ identity mutants apetala2-1 and agamous1. Can. J. Bot. 1994, 72, 384–401. [Google Scholar] [CrossRef]
- Satina, S.; Blakeslee, A.F. Periclinal chimeras in Datura in relation to the development of the carpel. Am. J. Bot. 1943, 30, 453–462. [Google Scholar] [CrossRef]
- Szymkowiak, E.J.; Sussex, I.M. The internal meristem layer (L3) determines floral meristem size and carpel number in tomato periclinal chimeras. Plant Cell 1992, 4, 1089–1100. [Google Scholar] [PubMed] [Green Version]
- Stewart, R.N.; Burk, L.G. Independence of tissues derived from apical layers in ontogeny of the tobacco leaf and ovary. Am. J. Bot. 1970, 57, 1010–1016. [Google Scholar] [CrossRef]
- Cheung, M.; Sattler, R. Early floral development of Lythrum salicaria. Can. J. Bot. 1967, 45, 1609–1623. [Google Scholar] [CrossRef]
- Evans, P.T.; Malmberg, R.L. Alternative pathways of tobacco placental development: Time of commitment and analysis of a mutant. Dev. Biol. 1989, 136, 273–283. [Google Scholar] [CrossRef]
- Reiser, L.; Modrusan, Z.; Margossian, L.; Samach, A.; Ohad, N.; Haughn, G.W.; Fischer, R.L. The BELL1 gene encodes a homeodomain protein involved in pattern formation in the Arabidopsis ovule primordium. Cell 1995, 83, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Colombo, L.; Franken, J.; Koetje, E.; van Went, J.; Dons, H.J.M.; Angenent, G.C.; van Tunen, A.J. The Petunia MADS box gene FBP11 determines ovule identity. Plant Cell 1995, 7, 1859–1868. [Google Scholar]
- Lesley, M.M. Maturation in diploid and triploid tomatoes. Genetics 1926, 11, 267–279. [Google Scholar] [PubMed]
- Satina, S. Periclinal chimeras in Datura in relation to the development and structure of the ovule. Am. J. Bot. 1945, 32, 72–81. [Google Scholar] [CrossRef]
- Goodspeed, T.H. The Genus Nicotiana: Origins, Relationships and Evolution of Its Species in the Light of Their Distribution, Morphology and Cytogenetics; Chronica Botanica Co.: Waltham, MA, USA, 1954; pp. 167–176. [Google Scholar]
- George, G.P.; George, R.A.; Herr, J.M., Jr. A comparative study of ovule and megagametophyte development in field-grown and greenhouse-grown plants of Glycine max and Phaseolus aureus (Papilionaceae). Am. J. Bot. 1979, 69, 1033–1043. [Google Scholar] [CrossRef]
- Moço, M.C.C.; Mariath, J.E.A. Ovule ontogenesis and megasporogenesis in Adesmia latifolia (Spreng.) Vog. (Leguminosae-Papilionoideae). Rev. Bras. Bot. 2003, 26, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Bachelier, J.B.; Friedman, W.E. Female gamete competition in an ancient angiosperm lineage. Proc. Natl. Acad. Sci. USA 2011, 108, 12360–12365. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, K.-I. The MSP1 gene is necessary to restrict the number of cells entering into male and female sporogenesis and to initiate anther wall formation in rice. Plant Cell 2003, 15, 1728–1739. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Zhao, L.; Zhao, Y.; Li, S.; Won, S.; Cai, H.; Wang, L.; Li, Z.; Chen, P.; Qin, Y.; et al. The THO complex non-cell- autonomously represses female germline specification through the TAS3-ARF3 module. Curr. Biol. 2017, 27, 1597–1609. [Google Scholar] [CrossRef] [Green Version]
- Reiser, L.; Fischer, R.L. The ovule and the embryo sac. Plant Cell 1993, 5, 1291–1301. [Google Scholar] [CrossRef]
- Lora, J.; Yang, X.; Tucker, M.R. Establishing a framework for female germline initiation in the plant ovule. J. Exp. Bot. 2019, 70, 2937–2949. [Google Scholar] [CrossRef]
- Shirley, N.J.; Aubert, M.; Wilkinson, L.; Bird, D.; Lora, J.; Yang, X.; Tucker, M.R. Translating auxin responses into ovules, seeds and yield: Insight from Arabidopsis and the cereals. J. Integr. Comp. Biol. 2019, 61, 310–336. [Google Scholar] [CrossRef] [Green Version]
- Willemse, M.T.M.; van Went, J.L. The female gametophyte. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 159–196. [Google Scholar]
- Soliman, M.; Espinoza, F.; Ortiz, J.P.O.; Delgado, L. Heterochronic reproductive developmental processes between diploid and tetraploid cytotypes of Paspalum rufum. Ann. Bot. 2019, 123, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Green, P.B.; Linstead, P. A procedure for SEM of complex shoot structures applied to the inflorescence of snapdragon (Antirrhinum). Protoplasma 1990, 158, 33–38. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flower Position | Plastochron ± Standard Deviation |
|---|---|
| 2 | 1.45 ± 0.26 |
| 3 | 1.57 ± 0.31 |
| 4 | 1.84 ± 0.10 |
| 5 | 2.06 ± 0.35 |
| 6 | 1.98 ± 0.19 |
| 7 | 2.55 ± 0.38 |
| 8 | 2.85 ± 0.45 |
| Cyme # | PI |
|---|---|
| 1 (2.0 cm) | 1.61 |
| 2 (2.5 cm) | 1.78 |
| 3 (3.0 cm) | 2.17 |
| 4 (3.5 cm) | 2.54 |
| 5 (4.0 cm) | 2.65 |
| 6 (4.5 cm) | 3.00 |
| Anther Age (Days after ECP) and Developmental Characterization | Stage Defined in [30] | Age (Day) Reported in [6] |
|---|---|---|
| 0 (Figure 1A,B): Filaments and anthers differentiated in stamen primordia. Periclinal divisions in the hypodermal layer (L2) in the anther were observed. A single procambial strand was shown in the region of the anther connective. | Stage –7 | 3.0 |
| 1.3 ± 1.5 (Figure 1C): Four separate microsporangia were evident. Periclinal divisions of L2 cells were observed. | Stage –6 | 3.7 |
| 3.0 ± 0.2 (Figure 2I): The tetrasporangiate anthers was evident. Most of the archesporial cells completed the first round of periclinal divisions, while the inner tapetal cells were identified beneath a layer of archesporial cells. The majority of cells in L2 completed a recent periclinal division. | Stages –6, –5 | 4.4 |
| 4.8 ± 1.3 (Figure 3K): The anther generally contained 3–4 layers of cells between the inner tapetum and the epidermis. The region of 1–2 layers of sporogenous cells immediately adjacent to the inner tapetum became cytologically distinct, while no outer tapetum differentiated. | Stage –5 | 4.7–5.0 |
| 5.4 (Figure 4D): The anther contained 3–4 layers of cells between the layer of sporogenous tissue and the epidermis. The outer tapetum differentiated as 1–2 layers of cells immediately outside the sporogenous tissue. | Stages –4, –3 | 5.9 |
| 6.1 ± 0.6 (Figure 4E): Sporogenous cells in anthers were in mitotic proliferation. The outer tapetum was evident as a layer of 1–2 smaller cells immediately outside the sporogenous layer. Some cells in the stomium and circular cell cluster (CCC, a group of cells underneath the stomium layer that accumulate crystals and degenerate when the two microsporangia fuse into one locule prior to anther dehiscence) were densely stained. Recent periclinal divisions were observed in the CCC region without any crystal accumulation in these cells. | Stages –3, –2 | 6.0 |
| 8.9 ± 1.1 (Figure 6A): Microsporocytes entered prophase I of meiosis. More periclinal divisions were observed in the cells of CCC and some CCC cells contained crystals. | Stages –2, –1 | 6.6 |
| 10.0 ± 0.5 (Figure 6E): Microsporocyptes were separated from the anther wall. Periclinal divisions were observed in stomium cells, while some outer tapetal cells began to degrade. | Stage –1 | N/A |
| 10.9 ± 1.1 (Figure 7D): Microspores were in tetrads and tapetum was degenerating. | Stage 1 | N/A |
| 12.9 ± 1.2 (Figure 8E,F): Microspores were separated from each other. Cells of CCC contained crystals and lost staining. Tapetal cells were degenerating. | Stages 2–4 | N/A |
| 14.0 ± 1.2 (Figure 8G): Microspores were in mitotic divisions in anaphase. Tapetal cells were further degraded, while the remnants of the tapetum were still present. Secondary thickening on the cell walls of the endothecial layer in the anthers was observed, while the CCC cells began to degrade and some stomium cells collapsed. | Stages 5–6 | N/A |
| 16.0 ± 0.4 (Figure 9E): Both the vegetative and generative cells were observed in pollen grains. The cells of CCC completely degraded, while the two pollen sacs fused into one locule. Most connective cells were still intact, while the tapetal cells almost completely degraded. The anthers did not open yet. | Stages 7–8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, H.; Sun, F. Temporal Distinction between Male and Female Floral Organ Development in Nicotiana tabacum cv. Xanthi (Solanaceae). Plants 2020, 9, 127. https://doi.org/10.3390/plants9010127
Chang H, Sun F. Temporal Distinction between Male and Female Floral Organ Development in Nicotiana tabacum cv. Xanthi (Solanaceae). Plants. 2020; 9(1):127. https://doi.org/10.3390/plants9010127
Chicago/Turabian StyleChang, Hongli, and Fengjie Sun. 2020. "Temporal Distinction between Male and Female Floral Organ Development in Nicotiana tabacum cv. Xanthi (Solanaceae)" Plants 9, no. 1: 127. https://doi.org/10.3390/plants9010127