Phenolic Compounds as Unambiguous Chemical Markers for the Identification of Keystone Plant Species in the Bale Mountains, Ethiopia

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
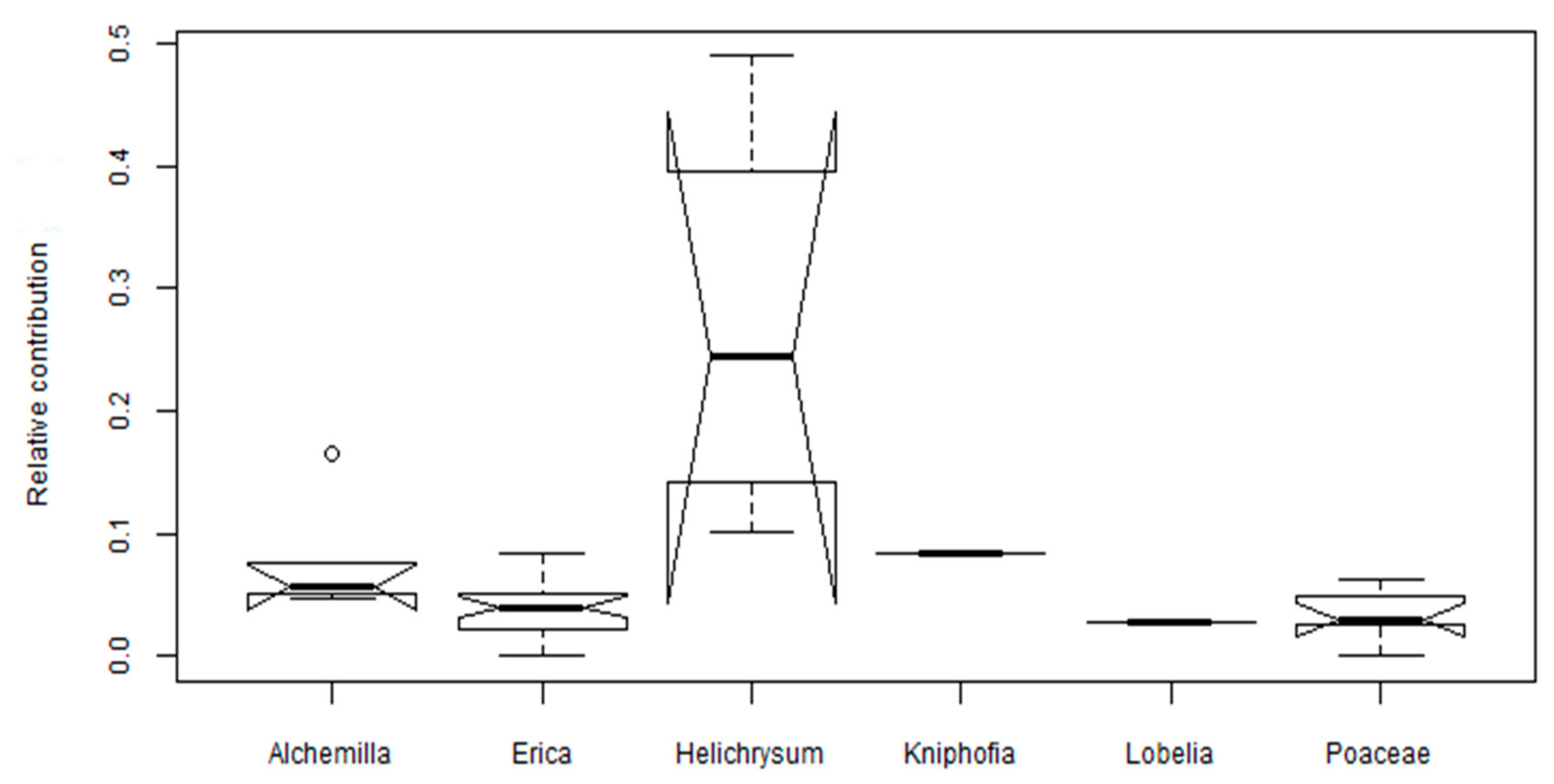
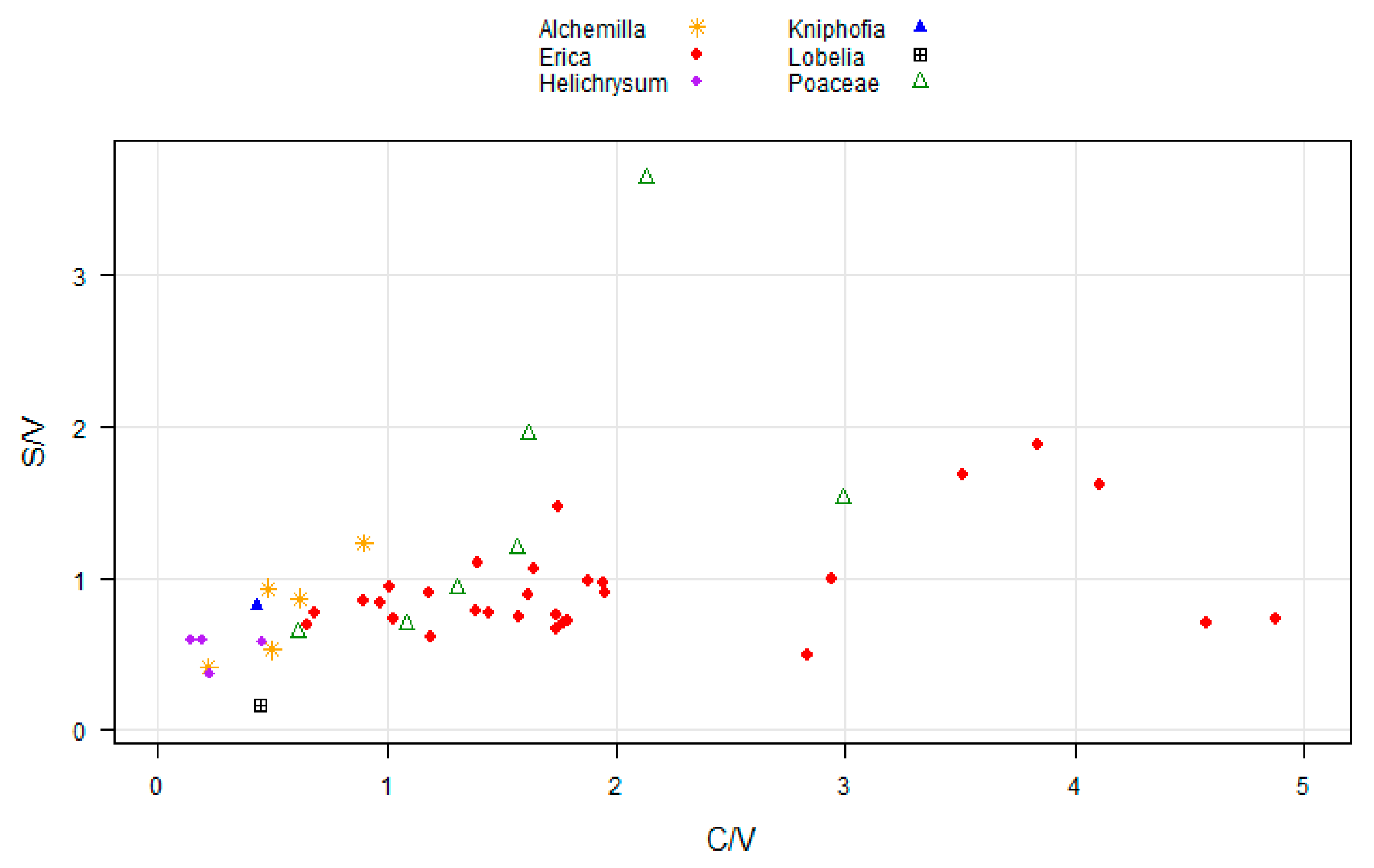
2.1. Distribution and Diversity of Phenols
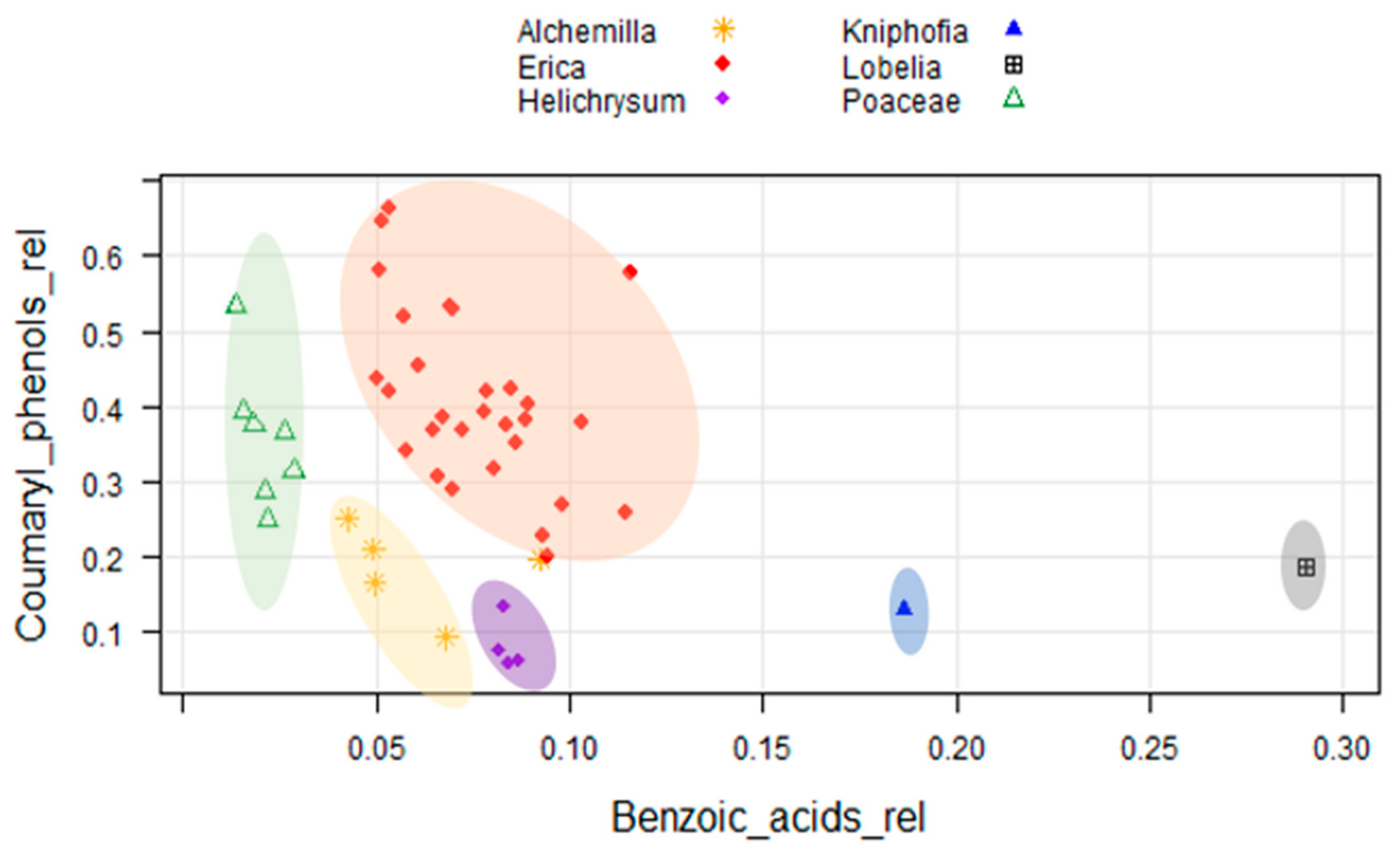
2.2. Cluster Analysis of Phenolic Compounds
2.3. Biomarker Identification
3. Material and Methods
3.1. Study Area
3.1.1. Geomorphological Setting
3.1.2. Climate and Biota
3.2. Sample Collection
3.3. Analysis of Phenolic Compounds Released after Alkaline CuO Oxidation
3.4. Data Analysis
3.4.1. Hierarchical Clustering and PCA
3.4.2. Machine Learning
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heiri, C.; Bugmann, H.; Tinner, W.; Heiri, O.; Lischke, H. A model-based reconstruction of Holocene treeline dynamics in the Central Swiss Alps. J. Ecol. 2006, 94, 206–216. [Google Scholar] [CrossRef]
- Umer, M.; Lamb, H.F.; Bonnefille, R.; Lézine, A.M.; Tiercelin, J.J.; Gibert, E.; Cazet, J.P.; Watrin, J. Late Pleistocene and Holocene vegetation history of the Bale Mountains, Ethiopia. Quat. Sci. Rev. 2007, 26, 2229–2246. [Google Scholar] [CrossRef]
- Jansen, B.; van Loon, E.E.; Hooghiemstra, H.; Verstraten, J.M. Improved reconstruction of palaeo-environments through unravelling of preserved vegetation biomarker patterns. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 285, 119–130. [Google Scholar] [CrossRef]
- Terwilliger, V.J.; Eshetu, Z.; Colman, A.; Bekele, T.; Gezahgne, A.; Fogel, M.L. Reconstructing palaeoenvironment from δ13C and δ15N values of soil organic matter: A calibration from arid and wetter elevation transects in Ethiopia. Geoderma 2008, 147, 197–210. [Google Scholar] [CrossRef]
- Tiercelin, J.J.; Gibert, E.; Umer, M.; Bonnefille, R.; Disnar, J.R.; Lézine, A.M.; Hureau-Mazaudier, D.; Travi, Y.; Keravis, D.; Lamb, H.F. High-resolution sedimentary record of the last deglaciation from a high-altitude lake in Ethiopia. Quat. Sci. Rev. 2008, 27, 449–467. [Google Scholar] [CrossRef] [Green Version]
- Hillman, J.C. Conservation in Bale Mountains National Park, Ethiopia. Oryx 1986, 20, 89–94. [Google Scholar] [CrossRef]
- Hillman, J.C. The Bale Mountains National Park area, Southeast Ethiopia, and its management. Mt. Res. Dev. 1988, 8, 253–258. [Google Scholar] [CrossRef]
- Friis, I. Zonation of Forest Vegetation on the South Slope of Bale Mountains, South Ethiopia. SINET Ethiop. J. Sci. 1986, 9, 29–44. [Google Scholar]
- Miehe, G.; Miehe, S. Ericaceous Forests and Heathlands in the Bale Mountains of South Ethiopia: Ecology and Man’s Impact; T. Warnke Verlag: Hamburg, Germany, 1994; p. 206. [Google Scholar]
- Hedberg, O. Features of Afroalpine Plant Ecology; Swedish Science Press: Uppsala, Sweden, 1964; p. 148. [Google Scholar]
- Kidane, Y.; Stahlmann, R.; Beierkuhnlein, C. Vegetation dynamics, and land use and land cover change in the Bale Mountains, Ethiopia. Environ. Monit. Assess. 2012, 184, 7473–7489. [Google Scholar] [CrossRef]
- Bonnefille, R.; Hamilton, A. Quaternary and Late Tertiary history of Ethiopian vegetation. Symb. Bot. Ups. 1986, 26, 48–63. [Google Scholar]
- Bonnefille, R.; Mohammed, U. Pollen-inferred climatic fluctuations in Ethiopia during the last 3000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 109, 331–343. [Google Scholar] [CrossRef]
- Hamilton, A.C. Environmental History of East Africa: A Study of the Quaternary; Academic Press: London, UK, 1982; p. 328. [Google Scholar]
- Hicks, S. When no pollen does not mean no trees. Veg. Hist. Archaeobotany 2006, 15, 253–261. [Google Scholar] [CrossRef]
- Ortu, E.; Brewer, S.; Peyron, O. Pollen-inferred palaeoclimate reconstructions in mountain areas: Problems and perspectives. J. Quat. Sci. 2006, 21, 615–627. [Google Scholar] [CrossRef]
- Glaser, B.; Zech, W. Reconstruction of climate and landscape changes in a high mountain lake catchment in the Gorkha Himal, Nepal during the Late Glacial and Holocene as deduced from radiocarbon and compound-specific stable isotope analysis of terrestrial, aquatic and microbial biomarkers. Org. Geochem. 2005, 36, 1086–1098. [Google Scholar] [CrossRef]
- Ram Singh, C. Chemotaxonomy: A Tool for Plant Classification. J. Med. Plants Stud. JMPS 2016, 90, 90–93. [Google Scholar]
- Ertel, J.R.; Hedges, J.I. The lignin component of humic substances: Distribution among soil and sedimentary humic, fulvic, and base-insoluble fractions. Geochim. Cosmochim. Acta 1984, 48, 2065–2074. [Google Scholar] [CrossRef]
- Míka, V.; Kubáň, V.; Klejdus, B.; Odstrčilová, V.; Nerušil, P. Phenolic compounds as chemical markers of low taxonomic levels in the family Poaceae. Plant Soil Environ. 2005, 51, 506–512. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar]
- Witzell, J.; Gref, R.; Na, T. Plant-part specific and temporal variation in phenolic compounds of boreal bilberry (Vaccinium myrtillus) plants. Biochem. Syst. Ecol. 2003, 31, 115–127. [Google Scholar] [CrossRef]
- Nichols-Orians, C.M.; Fritz, R.S.; Clausen, T.P. The genetic basis for variation in the concentration of phenolic glycosides in Salix sericea: Clonal variation and sex-based differences. Biochem. Syst. Ecol. 1993, 21, 535–542. [Google Scholar] [CrossRef]
- Hakulinen, J.; Julkunen-Tiitto, R.; Tahvanainen, J. Does nitrogen fertilization have an impact on the trade-off between willow growth and defensive secondary metabolism? Trees 1995, 9, 235–240. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Iwasaki, M.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the Rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and quantification of phenolic compounds in berries of Fragaria and Rubus species (family Rosaceae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar] [CrossRef]
- Fernandez-Lorenzo, J.L.; Rigueiro, A.; Ballester, A. Polyphenols as potential markers to differentiate juvenile and mature chestnut shoot cultures. Tree Physiol. 1999, 19, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.-M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. A chemotaxonomic survey of flavonoids and simple phenols in leaves of the Ericaceae. Bot. J. Linn. Soc. 1973, 66, 37–54. [Google Scholar] [CrossRef]
- Jay, M. Chemotaxonomic researches on vascular plants. XIX. Flavonoid distribution in the Pittosporaceae. Bot. J. Linn. Soc. 1969, 62, 423–429. [Google Scholar] [CrossRef]
- Giannasi, D.E. Systematic aspects of flavonoid biosynthesis and evolution. Bot. Rev. 1978, 44, 399–429. [Google Scholar] [CrossRef]
- Umadevi, I.; Daniel, M.; Sabnis, S.D. Chemotaxonomic studies on some members of Anacardiaceae. Proc. Plant Sci. 1988, 98, 205–208. [Google Scholar]
- Guendouze-Bouchefa, N.; Madani, K.; Chibane, M.; Boulekbache-Makhlouf, L.; Hauchard, D.; Kiendrebeogo, M.; Stévigny, C.; Okusa, P.N.; Duez, P. Phenolic compounds, antioxidant and antibacterial activities of three Ericaceae from Algeria. Ind. Crop. Prod. 2015, 70, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Márquez-García, B.; Fernández, M.Á.; Córdoba, F. Phenolics composition in Erica sp. differentially exposed to metal pollution in the Iberian Southwestern Pyritic Belt. Bioresour. Technol. 2009, 100, 446–451. [Google Scholar] [CrossRef]
- Márquez-García, B.; Fernández-Recamales, M.; Córdoba, F. Effects of cadmium on phenolic composition and antioxidant activities of Erica andevalensis. J. Bot. 2012, 2012, 936950. [Google Scholar] [CrossRef]
- Djurdjević, L.; Mitrović, M.; Pavlović, P.; Perišić, S. Total phenolics and phenolic acids content in low (Chrysopogon gryllus) and mediocre quality (Festuca vallesiaca) forage grasses of Deliblato Sands meadow-pasture communities in Serbia. Czech J. Anim. Sci. 2005, 50, 54–59. [Google Scholar] [CrossRef]
- Pljevljakušić, D.; Bigović, D.; Janković, T.; Jelačić, S.; Šavikin, K. Sandy everlasting (Helichrysum arenarium (L.) Moench): Botanical, chemical and biological properties. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Babotă, M.; Mocan, A.; Vlase, L.; Crișan, O.; Ielciu, I.; Gheldiu, A.-M.; Vodnar, D.; Crișan, G.; Păltinean, R. Phytochemical analysis, antioxidant and antimicrobial activities of Helichrysum arenarium (L.) Moench. and Antennaria dioica (L.) Gaertn. flowers. Molecules 2018, 23, 409. [Google Scholar] [CrossRef]
- Albayrak, S.; Aksoy, A.; SaĞdiÇ, O.; Budak, Ü. Phenolic compounds and antioxidant and antimicrobial properties of Helichrysum species collected from eastern Anatolia, Turkey. Turk. J. Biol. 2010, 34, 463–473. [Google Scholar]
- Sroka, Z.; Kuta, I.; Cisowski, W.; Dryś, A. Antiradical activity of hydrolyzed and non-hydrolyzed extracts from Helichrysi inflorescentia and its phenolic contents. Z. Naturforschung C 2004, 59, 363–367. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Liu, C.; Song, Z.; Li, Q.; Zha, Q.; Lu, C.; Wang, C.; Ning, Z.; Zhang, Y.; et al. The chemotaxonomic classification of Rhodiola plants and its correlation with morphological characteristics and genetic taxonomy. Chem. Cent. J. 2013, 7, 118. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, J.; Zhao, Y.-L.; Wang, Y.-Z.; Jin, H. Chemotaxonomic studies of nine Gentianaceae species from western China based on liquid chromatography tandem mass spectrometry and Fourier transform infrared spectroscopy. Phytochem. Anal. 2016, 27, 158–167. [Google Scholar] [CrossRef]
- Cruz, A.V.M.; Ferreira, M.J.; Scotti, M.T.; Kaplan, M.A.; Emerenciano, V.P. Chemotaxonomic relationships in Celastraceae inferred from principal component analysis (PCA) and partial least squares (PLS). Nat. Prod. Commun. 2008, 3, 911–917. [Google Scholar] [CrossRef]
- Chesson, A.; Stewart, C.S.; Wallace, R.J. Influence of Plant Phenolic Acids on Growth and Cellulolytic Activity of Rumen Bacteria. Appl. Environ. Microbiol. 1982, 44, 597–603. [Google Scholar] [Green Version]
- Hedges, J.I.; Mann, D.C. The characterization of plant tissues by their lignin oxidation products. Geochim. Cosmochim. Acta 1979, 43, 1803–1807. [Google Scholar] [CrossRef]
- Li, H.-B.; Wong, C.-C.; Cheng, K.-W.; Chen, F. Antioxidant properties in vitro and total phenolic contents in methanol extracts from medicinal plants. LWT Food Sci. Technol. 2008, 41, 385–390. [Google Scholar] [CrossRef]
- Nazemiyeh, H.; Bahadori, F.; Delazar, A.; Ay, M.; Topçu, G.; Nahar, L.; Majinda, R.R.; Sarker, S.D. Antioxidant phenolic compounds from the leaves of Erica arborea (Ericaceae). Nat. Prod. Res. 2008, 22, 1385–1392. [Google Scholar] [CrossRef]
- Stanković, M.; Ćurčić, S.; Zlatić, N.; Bojović, B. Ecological variability of the phenolic compounds of Olea europaea L. leaves from natural habitats and cultivated conditions. Biotechnol. Biotechnol. Equip. 2017, 31, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Thevenot, M.; Dignac, M.F.; Rumpel, C. Fate of lignins in soils: A review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar] [CrossRef]
- Dümig, A.; Knicker, H.; Schad, P.; Rumpel, C.; Dignac, M.F.; Kögel-Knabner, I. Changes in soil organic matter composition are associated with forest encroachment into grassland with long-term fire history. Eur. J. Soil Sci. 2009, 60, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Osmaston, H.A.; Mitchell, W.A.; Osmaston, J.A.N. Quaternary glaciation of the Bale Mountains, Ethiopia. J. Quat. Sci. 2005, 20, 593–606. [Google Scholar] [CrossRef]
- Billi, P. Geomorphological landscapes of Ethiopia. In Landscapes and Landforms of Ethiopia; Springer: Dordrecht, The Netherlands, 2015; pp. 3–32. [Google Scholar]
- Woldu, Z.; Feoli, E.; Nigatu, L. Partitioning an Elevation Gradient of Vegetation from Southeastern Ethiopia by Probabilistic Methods. Plateau 1989, 81, 189–198. [Google Scholar]
- Bonnefille, R. Evidence for a cooler and drier climate in the Ethiopian uplands towards 2.5 Myr ago. Nature 1983, 303, 487–491. [Google Scholar] [CrossRef]
- Admasu, E.; Thirgood, S.J.; Bekele, A.; Karen Laurenson, M. Spatial ecology of golden jackal in farmland in the Ethiopian Highlands. Afr. J. Ecol. 2004, 42, 144–152. [Google Scholar] [CrossRef]
- Kebede, S.; Travi, Y. Origin of the δ18O and δ2H composition of meteoric waters in Ethiopia. Quat. Int. 2012, 257, 4–12. [Google Scholar] [CrossRef]
- Segele, Z.T.; Lamb, P.J.; Leslie, L.M. Large-scale atmospheric circulation and global sea surface temperature associations with horn of Africa June-September rainfall. Int. J. Climatol. 2009, 29, 1075–1100. [Google Scholar] [CrossRef]
- Dullo, B.W.; Grootjans, A.P.; Roelofs, J.G.M.; Senbeta, A.F.; Fritz, C. Fen mires with cushion plants in Bale Mountains, Ethiopia. Mires Peat 2015, 15, 1–10. [Google Scholar]
- Hedges, J.I.; Ertel, J.R. Characterization of Lignin. Anal. Chem. 1982, 54, 174–178. [Google Scholar] [CrossRef]
- Goñi, M.A.; Hedges, J.I. Lignin dimers: Structures, distribution, and potential geochemical applications. Geochim. Cosmochim. Acta 1992, 56, 4025–4043. [Google Scholar] [CrossRef]
- Amelung, W.; Martius, C.; Bandeira, A.G.; Garcia, M.V.B.; Zech, W. Lignin characteristics and density fractions of termite nests in an Amazonian rain forest—Indicators of termite feeding guilds? Soil Biol. Biochem. 2002, 34, 367–372. [Google Scholar] [CrossRef]
- Möller, A.; Kaiser, K.; Zech, W. Lignin, carbohydrate, and amino sugar distribution and transformation in the tropical highland soils of northern Thailand under cabbage cultivation, Pinus reforestation, secondary forest, and primary forest. Aust. J. Soil Res. 2002, 40, 977–998. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2019. Available online: http://www.R-project.org/ (accessed on 26 April 2019).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Contribution | Alchemilla | Erica | Helichrysum | Kniphofia | Lobelia | Poaceae |
|---|---|---|---|---|---|---|
| Vanillyl phenols | ↑ | ↓ | ||||
| Vanillin | ||||||
| Vanillic_acid | ||||||
| _4_hydroxy_3_methoxyacetophenone | ||||||
| Syringyl phenols | ↑ | ↓ | ↑ | |||
| Syringaldehyde | ||||||
| Syringic_acid | ||||||
| _3_5_dimethoxy_4_acetophenone | ||||||
| Coumaryl phenols | ↑ | ↓ | ↑ | |||
| p_coumaric_acid | ||||||
| Ferulic_acid | ||||||
| p_hydroxy_phenols | ↓ | ↑ | ||||
| _4_hydroxybenzaldehyde | ||||||
| _4_hydroxybenzacetophenone | ||||||
| Benzoic_acids | ↑ | ↑ | ↓ | |||
| Benzoic_acid | ||||||
| Salicylic_acid | ||||||
| Phthalic_acid | ||||||
| Hydroxy_benzoic_acids | ↑ | ↓ | ↑ | |||
| _3_hydroxybenzoic_acid | ||||||
| _4_hydroxybenzoic_acid | ||||||
| _3_5_Dihydroxy_benzoic_acid |
| Species | Diversity Index (H) |
|---|---|
| Alchemilla haumannii | 2.28 |
| Erica spp. | 2.19 |
| Helichrysum splendidum | 1.63 |
| Kniphofia foliosa | 2.47 |
| Lobelia rhynchopetalum | 2.09 |
| Festuca abyssinica | 2.17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemma, B.; Grehl, C.; Zech, M.; Mekonnen, B.; Zech, W.; Nemomissa, S.; Bekele, T.; Glaser, B. Phenolic Compounds as Unambiguous Chemical Markers for the Identification of Keystone Plant Species in the Bale Mountains, Ethiopia. Plants 2019, 8, 228. https://doi.org/10.3390/plants8070228
Lemma B, Grehl C, Zech M, Mekonnen B, Zech W, Nemomissa S, Bekele T, Glaser B. Phenolic Compounds as Unambiguous Chemical Markers for the Identification of Keystone Plant Species in the Bale Mountains, Ethiopia. Plants. 2019; 8(7):228. https://doi.org/10.3390/plants8070228
Chicago/Turabian StyleLemma, Bruk, Claudius Grehl, Michael Zech, Betelhem Mekonnen, Wolfgang Zech, Sileshi Nemomissa, Tamrat Bekele, and Bruno Glaser. 2019. "Phenolic Compounds as Unambiguous Chemical Markers for the Identification of Keystone Plant Species in the Bale Mountains, Ethiopia" Plants 8, no. 7: 228. https://doi.org/10.3390/plants8070228