Effect of Water Stress during Grain Filling on Yield, Quality and Physiological Traits of Illpa and Rainbow Quinoa (Chenopodium quinoa Willd.) Cultivars

 ,
, 
Abstract
:1. Introduction
2. Results
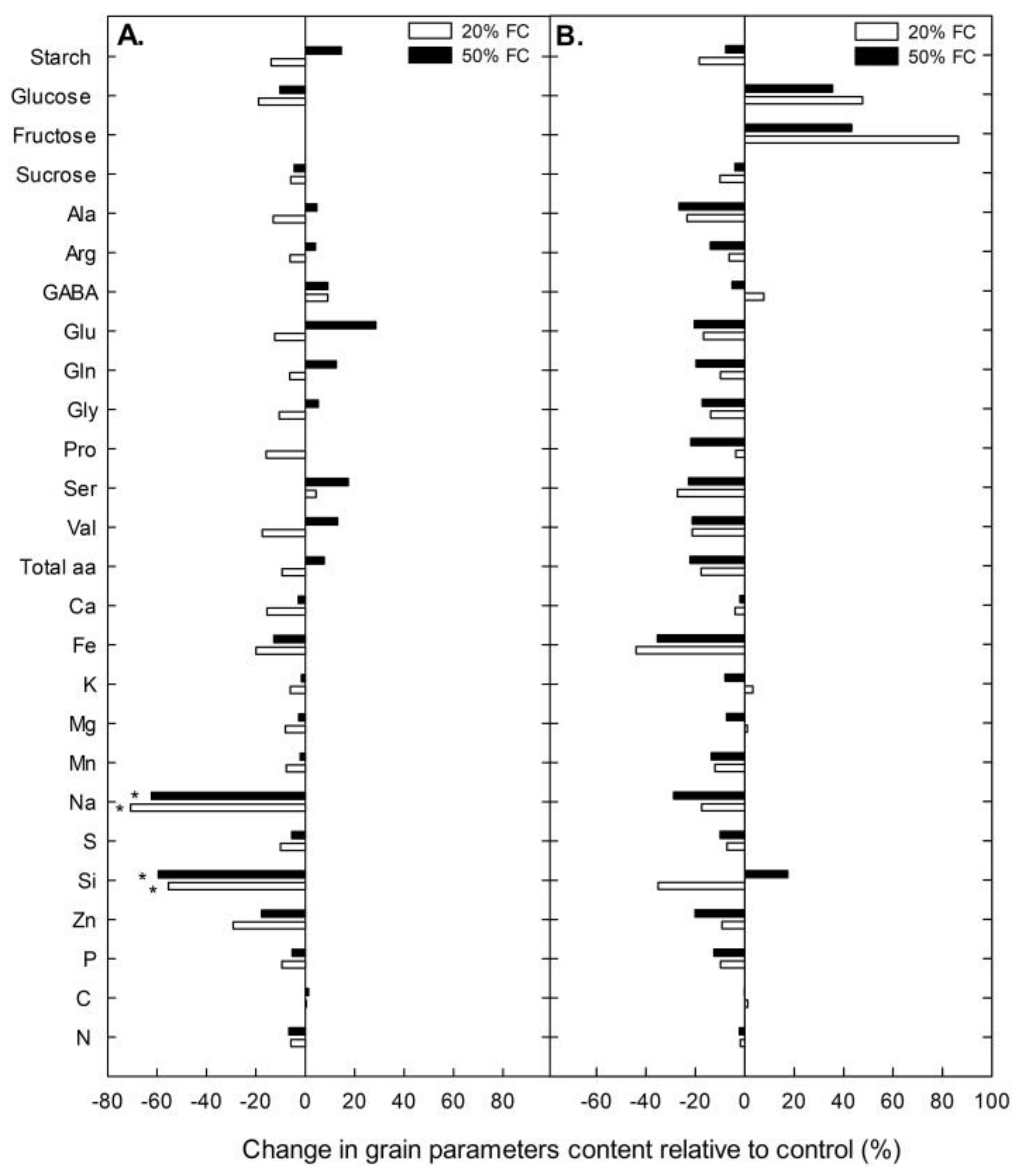
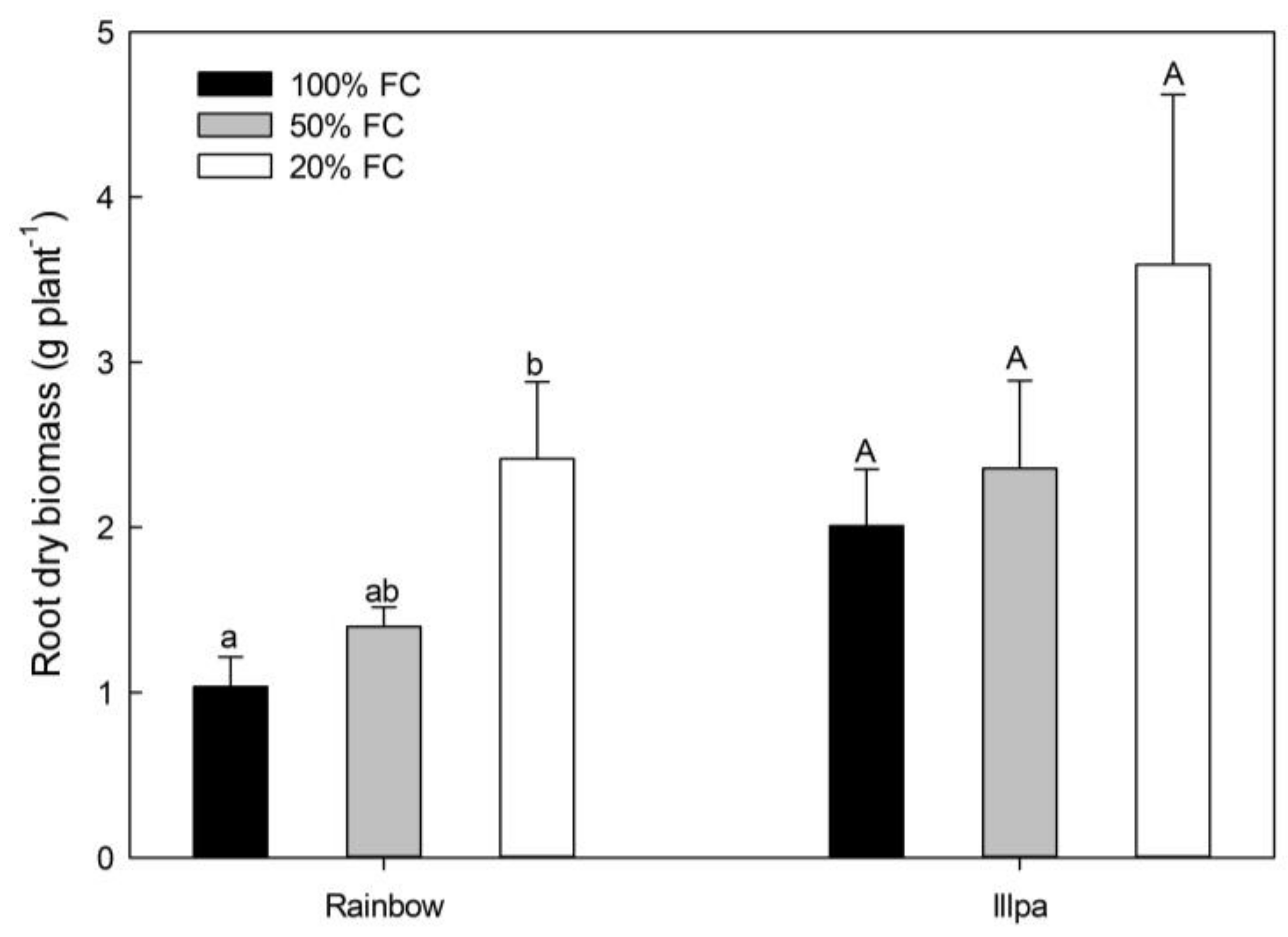
2.1. Crop Yield, Grain Quality and Root Biomass under Water Stress
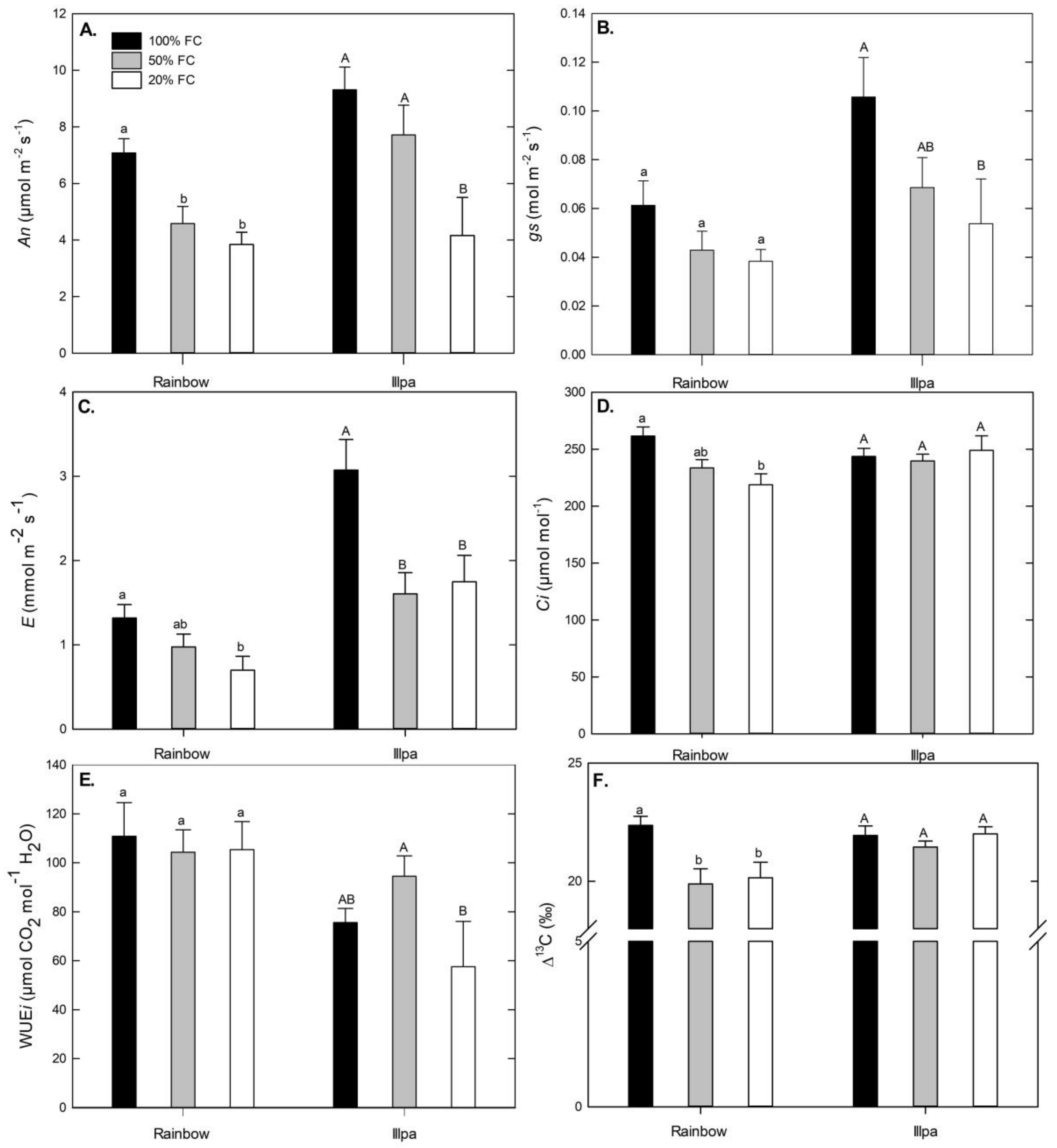
2.2. Physiological Response to Water Stress
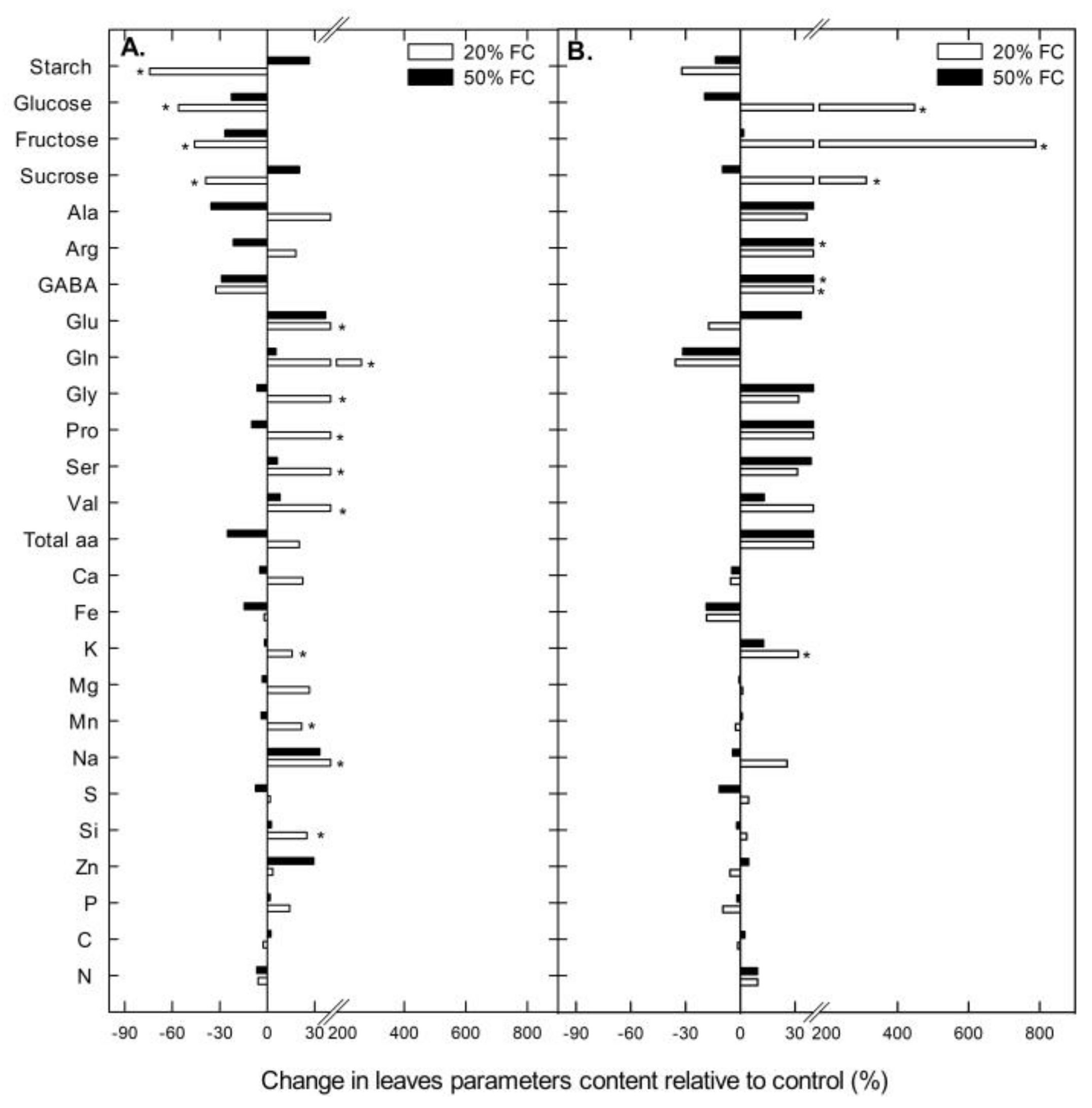
2.3. Leaf Carbohydrates, Amino Acids and Mineral Concentration under Water Stress
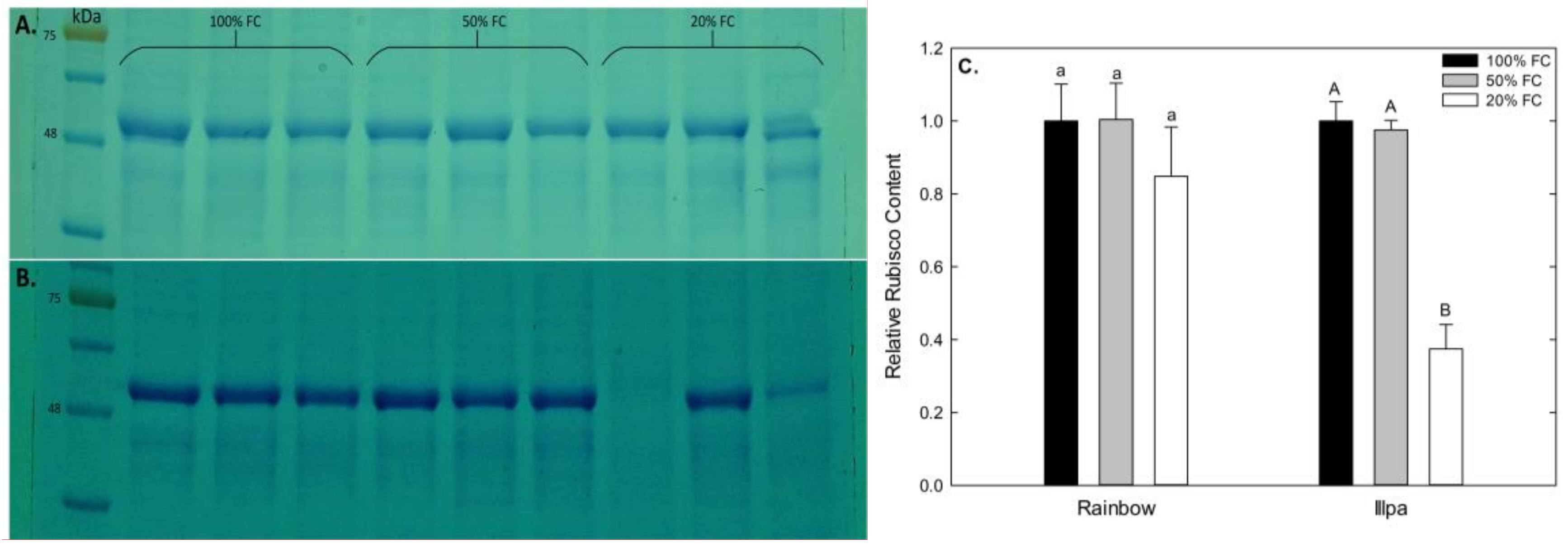
2.4. Relative Rubisco Concentration under Water Stress
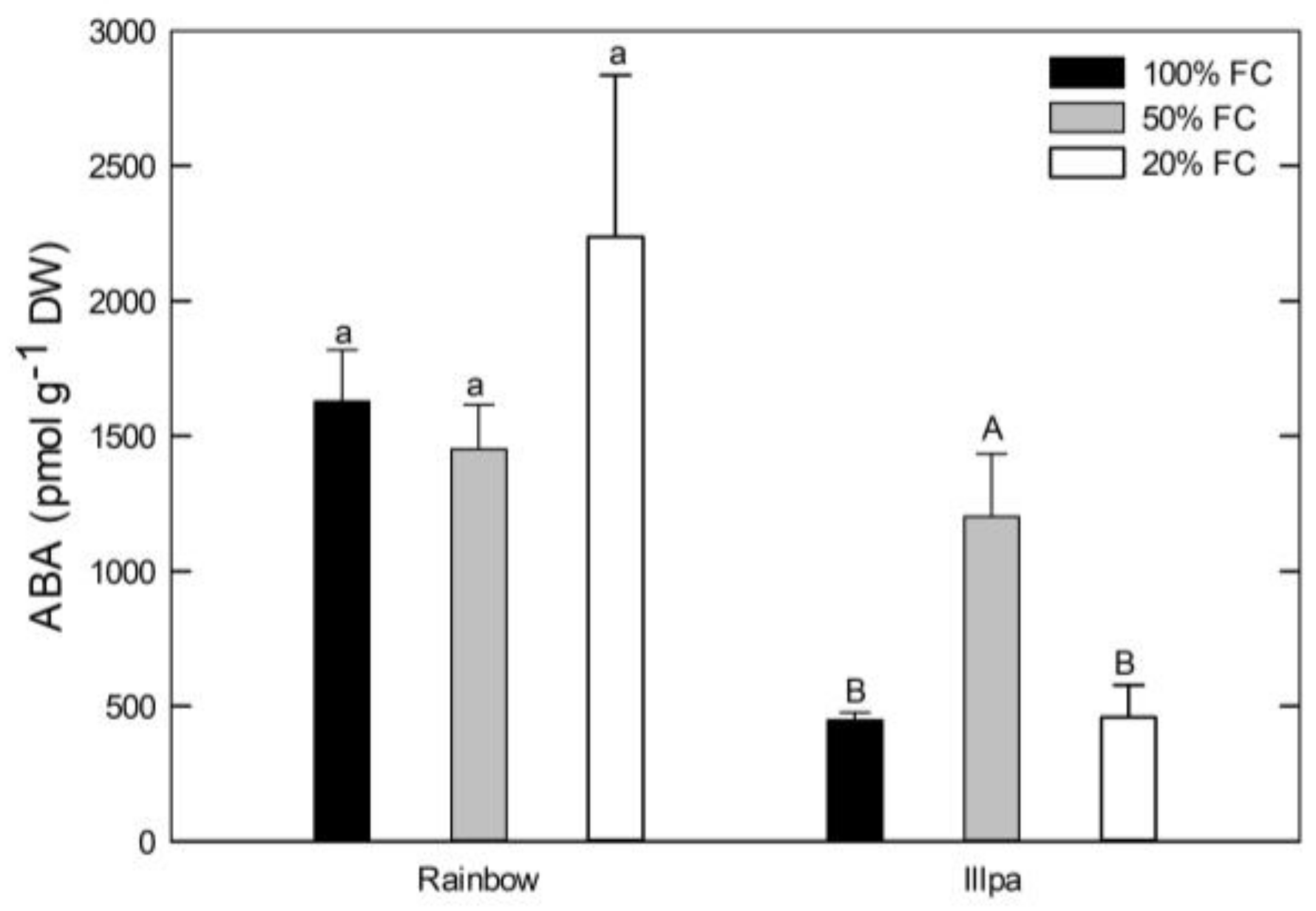
2.5. Leaf ABA Concentration under Water Stress
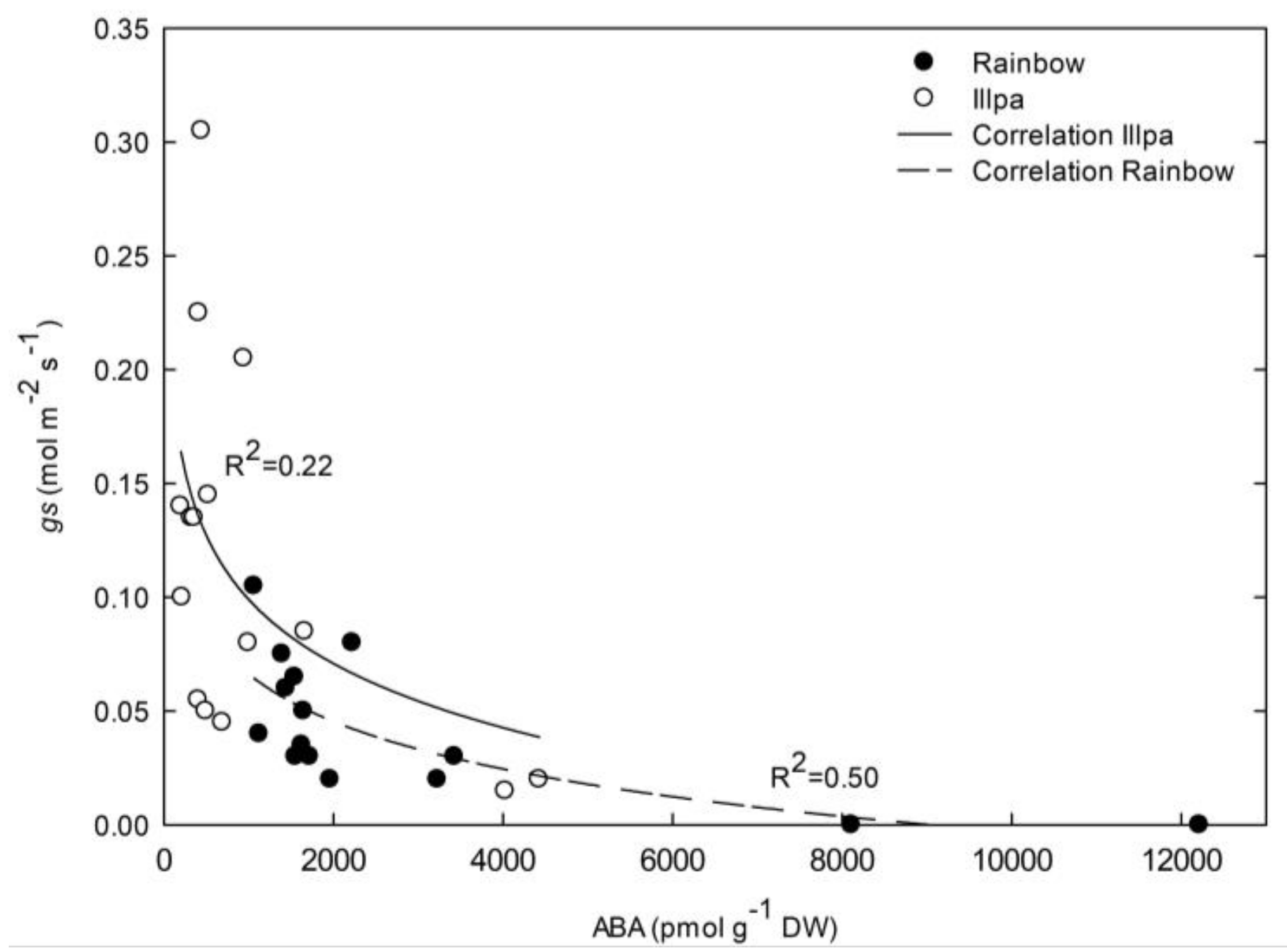
2.6. Relationship between Leaf ABA Concentration and Stomatal Closure under Water Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Plant Growth
4.3. Mineral Composition Analyses
4.4. Physiological Measurements
4.5. Carbohydrates and Amino Acids at Grain Filling
4.6. Relative Rubisco Content
4.7. Leaf Abscisic Acid Concentration
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Repo-Carrasco, R.; Espinoza, C.; Jacobsen, S.E. Nutritional value and use of the andean crops quinoa (Chenopodium quinoa) and kañiwa (Chenopodium pallidicaule). Food Rev. Int. 2003, 19, 179–189. [Google Scholar] [CrossRef]
- Abugoch, L.E. Quinoa (Chenopodium quinoa Willd.): Composition, chemistry, nutritional, and functional properties. In Advances in Food and Nutrition Research, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 58, pp. 1–31. [Google Scholar]
- Gordillo-Bastidas, E.; Díaz-Rizzolo, D.; Roura, E.; Massanés, T.; Gomis, R. Quinoa (Chenopodium quinoa Willd), from Nutritional Value to Potential Health Benefits: An Integrative Review. J. Nutr. Food Sci. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Bates, B.; Kundzewicz, Z.; Wu, S.; Palutikof, J. El Cambio Climático y el Agua; Intergovernmental Panel on Climate Change: Ginebra, Switzerland, 2008. [Google Scholar]
- Georgakopoulos, P.; Travlos, I.S.; Kakabouki, I.; Kontopoulou, C.K.; Pantelia, A.; Bilalis, D.J. Climate Change and Chances for the Cultivation of New Crops. Not. Bot. Horti Agrobo 2016, 44, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Jellen, E.; Maughan, P.; Bertero, D.; Munir, H. Prospects for Quinoa (Chenopodium Quinoa Willd.) improvement through biotechnology. In Biotechnology of Neglected and Underutilized Crops; Mohan, S., Gupta, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 173–203. [Google Scholar]
- Jacobsen, S.E.; Liu, F.; Jensen, C.R. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd.). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef]
- Jensen, C.; Jacobsen, S.-E.; Andersen, M.; Núñez, N.; Andersen, S.; Rasmussen, L.; Mogensen, V. Leaf gas exchange and water relation characteristics of field quinoa (Chenopodium quinoa Willd.) during soil drying. Eur. J. Agron. 2000, 13, 11–25. [Google Scholar] [CrossRef]
- Razzaghi, F.; Jacobsen, S.; Jensen, C.R.; Neumann, M. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought—mechanisms of tolerance. Funct. Plant Biol. 2015, 42, 136–148. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, F.; Bendevis, M.; Shabala, S.; Jacobsen, S. Sensitivity of two quinoa (Chenopodium quinoa Willd.) varieties to progressive drought stress. J. Agron. Crop. Sci. 2014, 200, 12–23. [Google Scholar] [CrossRef]
- Telahigue, D.C.; Ben Laila, Y.; Aljane, F.; Belhouchett, K.; Toumi, L. Grain yield, biomass productivity and water use efficiency in quinoa (Chenopodium quinoa Willd.) under drought stress. J. Sci. Agric 2017, 1, 222–232. [Google Scholar] [CrossRef]
- Alvarez-Flores, R.; Winkel, T.; Degueldre, D.; Del Castillo, C.; Joffre, R. Plant growth dynamics and root morphology of little-known species of Chenopodium from contrasted Andean habitats. Botany 2014, 92, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, E.F. Studies on Some Micro-Macromorphological and Anatomical Characters of Quinoa. J. Agric. Biol. Sci. 2014, 10, 24–36. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Review article Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agric. Water Manag. 2017, 179, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Campuzano, G.; Miralles, D.J.; Slafer, G.A. Genotypic variability and response to water stress of pre-and post-anthesis phases in triticale. Eur. J. Agron. 2008, 28, 171–177. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Wang, W. Remobilization of carbon reserves in response to water deficit during grain filling of rice. Field Crop. Res. 2001, 71, 47–55. [Google Scholar] [CrossRef]
- Flagella, Z.; Giuliani, M.M.; Giuzio, L.; Volpi, C.; Masci, S. Influence of water deficit on durum wheat storage protein composition and technological quality. Eur. J. Agron. 2010, 33, 197–207. [Google Scholar] [CrossRef]
- Fischer, S.; Wilckens, R.; Jara, J.; Aranda, M.; Valdivia, W.; Bustamente, L.; Graf, F.; Obal, I. Protein and antioxidant composition of quinoa (Chenopodium quinoa Willd.) sprout from seeds submitted to water stress, salinity and light conditions. Ind. Crop. Prod. 2017, 107, 558–564. [Google Scholar] [CrossRef]
- Morales, A.; Zurita-Silva, A.; Maldonado, J.; Silva, H. Transcriptional Responses of Chilean Quinoa (Chenopodium quinoa Willd.) Under Water Deficit Conditions Uncovers ABA-Independent Expression Patterns. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Alandia, G.; Jacobsen, S.; Kyvsgaard, N.C.; Condori, B.; Liu, F. Nitrogen Sustains Seed Yield of Quinoa Under Intermediate Drought. J. Agron. Crop. Sci. 2016, 202, 281–291. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Nam, N.H.; Chauhan, Y.S.; Johansen, C. Osmotic adjustment, water relations and carbohydrate remobilization in pigeonpea under water deficits. J. Plant Physiol. 2000, 157, 651–659. [Google Scholar] [CrossRef]
- Samarah, N. Effects of drought stress on growth and yield of barley. Agron. Sustain. Dev. 2005, 25, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Vacher, J.J. Responses of two main Andean crops, quinoa (Chenopodium quinoa Willd) and papa amarga (Solanum juzepczukii Buk.) to drought on the Bolivian Altiplano: Significance of local adaptation. Agric. Ecosyst. Environ. 1998, 68, 99–108. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Jensen, C.R.; Liu, F. Improving Crop Production in the Arid Mediterranean Climate. Field Crop. Res. 2013, 128, 187–209. [Google Scholar] [CrossRef]
- Kavar, T.; Maras, M.; Kidrič, M.; Šuštar-Vozlič, J.; Meglič, V. Identification of genes involved in the response of leaves of Phaseolus vulgaris to drought stress. Mol. Breed. 2008, 21, 159–172. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Bruno, M.; Valoy, M.; Prado, F.E. Genotypic Variation of Gas Exchange Parameters and Leaf Stable Carbon and Nitrogen Isotopes in Ten Quinoa Cultivars Grown under Drought. J. Agron. Crop. Sci. 2011, 197, 81–93. [Google Scholar] [CrossRef]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable Isotopes in Plant Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Tambussi, E.A.; Bort, J.; Araus, J.L. Water use efficiency in C3 cereals under Mediterranean conditions: A review of physiological aspects. Ann. Appl. Biol. 2007, 150, 307–321. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Discrimination and Photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 50–537. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, J.; Zhao, B.; Zhang, H.; Huang, P. Stable Isotope Studies of Crop Carbon and Water Relations: A Review. Agric. Sci. China 2009, 8, 578–590. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Amjad, M.; Iqbal, S.; Jacobsen, S.E. Growth and Physiological Responses of Quinoa to Drought and Temperature Stress. J. Agron. Crop. Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Medrano, H.; Ribas-Carbo, M.; Tomás, M.; et al. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cao, X.; Bai, Z.; Zhang, J.; Zhu, L.; Huang, J.; Jin, Q. Nitrogen metabolism correlates with the acclimation of photosynthesis to short-term water stress in rice (Oryza sativa L.). Plant Physiol. Biochem. 2018, 125, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Al-Naggar, A.; El-Salam, R.; Badran, A.; El-Moghazi, M. Drought tolerance of Five Quinoa (Chenopodium quinoa Willd.) Genotypes and Its Association with Other Traits under Moderate and Severe Drought Stress. Asian J. Adv. Agric. Res. 2017, 3, 1–13. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Laloum, T.; Martín, G.; Duque, P. Alternative Splicing Control of Abiotic Stress Responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S. The situation for Quinoa and its production in southern Bolivia: From economic success to environmental disaster. J. Agron. Crop. Sci. 2011, 197, 390–399. [Google Scholar] [CrossRef]
- Gomez, L.; Aguilar, E. Guía de Cultivo de la Quinua; Universdad Nacional Agraria La Molina, Ed.; FAO: Lima, Perú, 2016. [Google Scholar]
- Torres, N.; Goicoechea, N.; Zamarreño, A.M.; Antolín, M.C. Mycorrhizal symbiosis affects ABA metabolism during berry ripening in Vitis vinifera L. cv. Tempranillo grown under climate change scenarios. Plant Sci. 2018, 274, 383–393. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation | Grain Yield (g Plant−1) | Thousand-Grain Weight (g) | Grain Number | |||
|---|---|---|---|---|---|---|
| Rainbow | Illpa | Rainbow | Illpa | Rainbow | Illpa | |
| 100% FC | 15.66 ± 2.09 | 3.79 ± 0.92 | 2.07 ± 0.04 | 3.20 ± 0.17a | 7480 ± 1278 | 1244 ± 360 |
| 50% FC | 13.44 ± 0.83 | 3.30 ± 1.34 | 1.96 ± 0.06 | 2.81 ± 0.26ab | 6871 ± 631 | 1319 ± 622 |
| 20% FC | 16.78 ± 1.50 | 2.79 ± 1.09 | 2.09 ± 0.05 | 2.38 ± 0.17b | 8142 ± 1233 | 1181 ± 506 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gámez, A.L.; Soba, D.; Zamarreño, Á.M.; García-Mina, J.M.; Aranjuelo, I.; Morales, F. Effect of Water Stress during Grain Filling on Yield, Quality and Physiological Traits of Illpa and Rainbow Quinoa (Chenopodium quinoa Willd.) Cultivars. Plants 2019, 8, 173. https://doi.org/10.3390/plants8060173
Gámez AL, Soba D, Zamarreño ÁM, García-Mina JM, Aranjuelo I, Morales F. Effect of Water Stress during Grain Filling on Yield, Quality and Physiological Traits of Illpa and Rainbow Quinoa (Chenopodium quinoa Willd.) Cultivars. Plants. 2019; 8(6):173. https://doi.org/10.3390/plants8060173
Chicago/Turabian StyleGámez, Angie L., David Soba, Ángel M. Zamarreño, José M. García-Mina, Iker Aranjuelo, and Fermín Morales. 2019. "Effect of Water Stress during Grain Filling on Yield, Quality and Physiological Traits of Illpa and Rainbow Quinoa (Chenopodium quinoa Willd.) Cultivars" Plants 8, no. 6: 173. https://doi.org/10.3390/plants8060173