Morphological and Transcriptome Analysis of Wheat Seedlings Response to Low Nitrogen Stress

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Low Nitrogen Stress Affects Wheat Seedlings Morphology
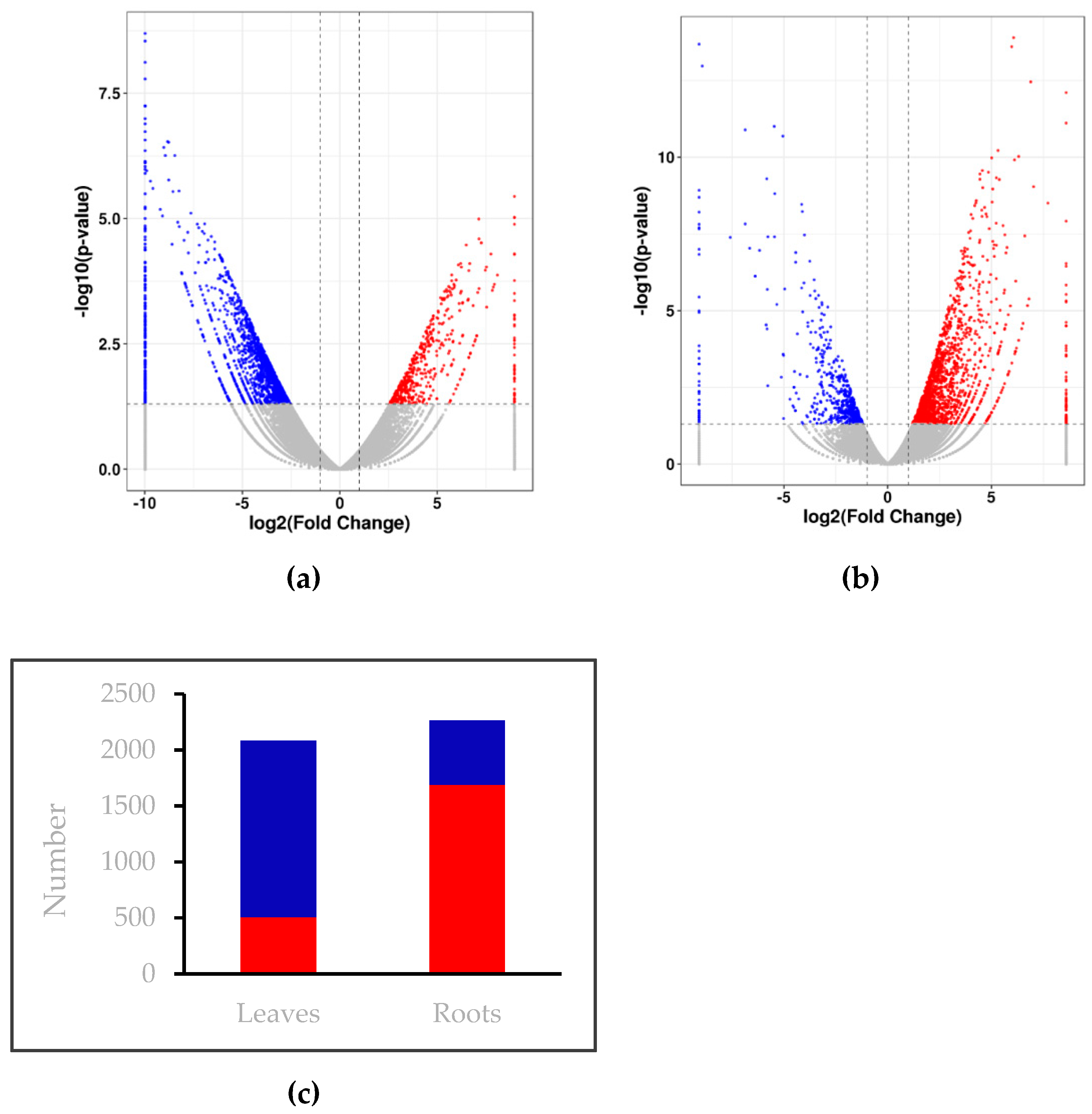
2.2. Overview of Transcriptome Sequencing Results
2.3. Low Nitrogen Stress Affects Genes Expression in Wheat Seedlings
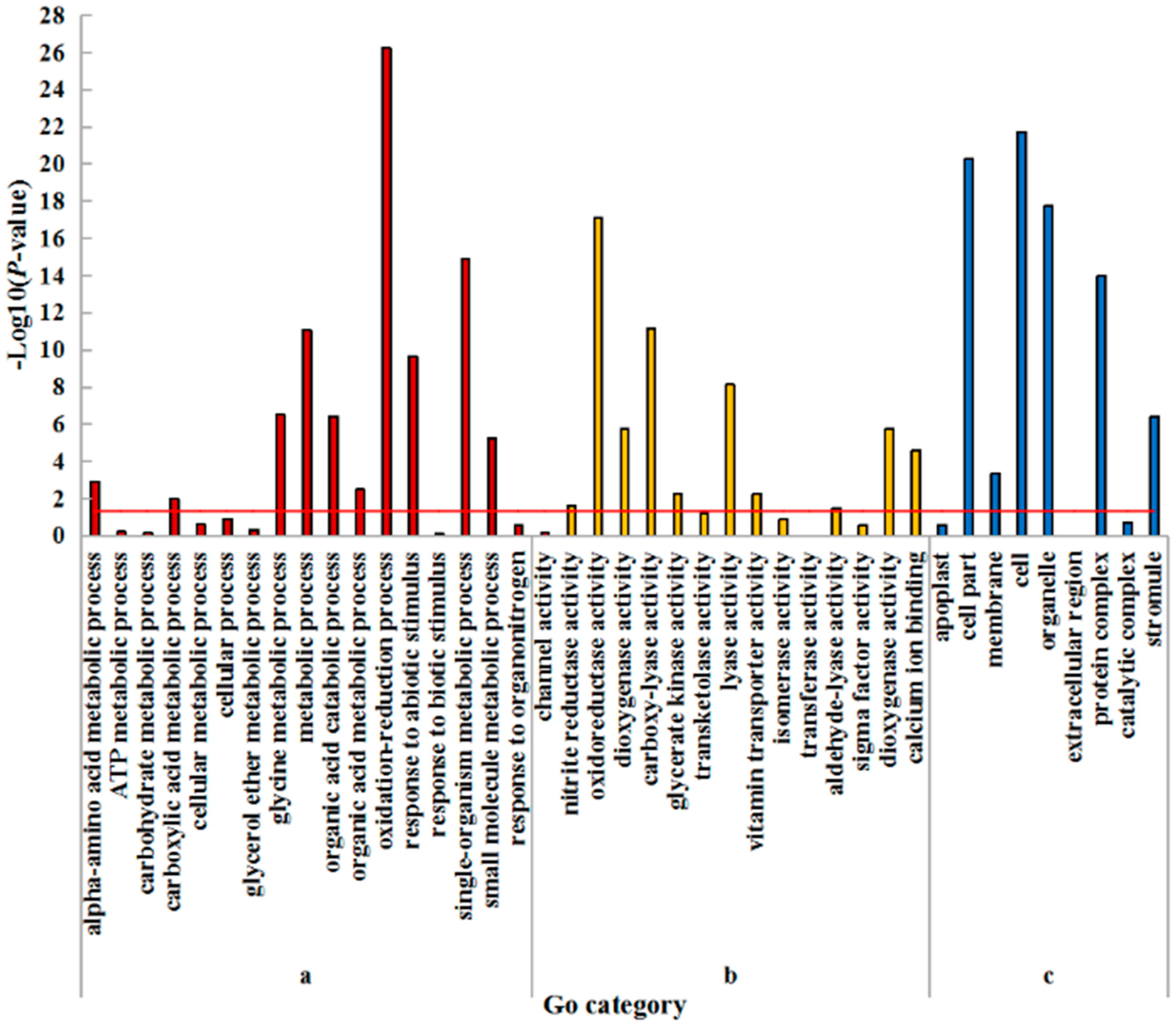
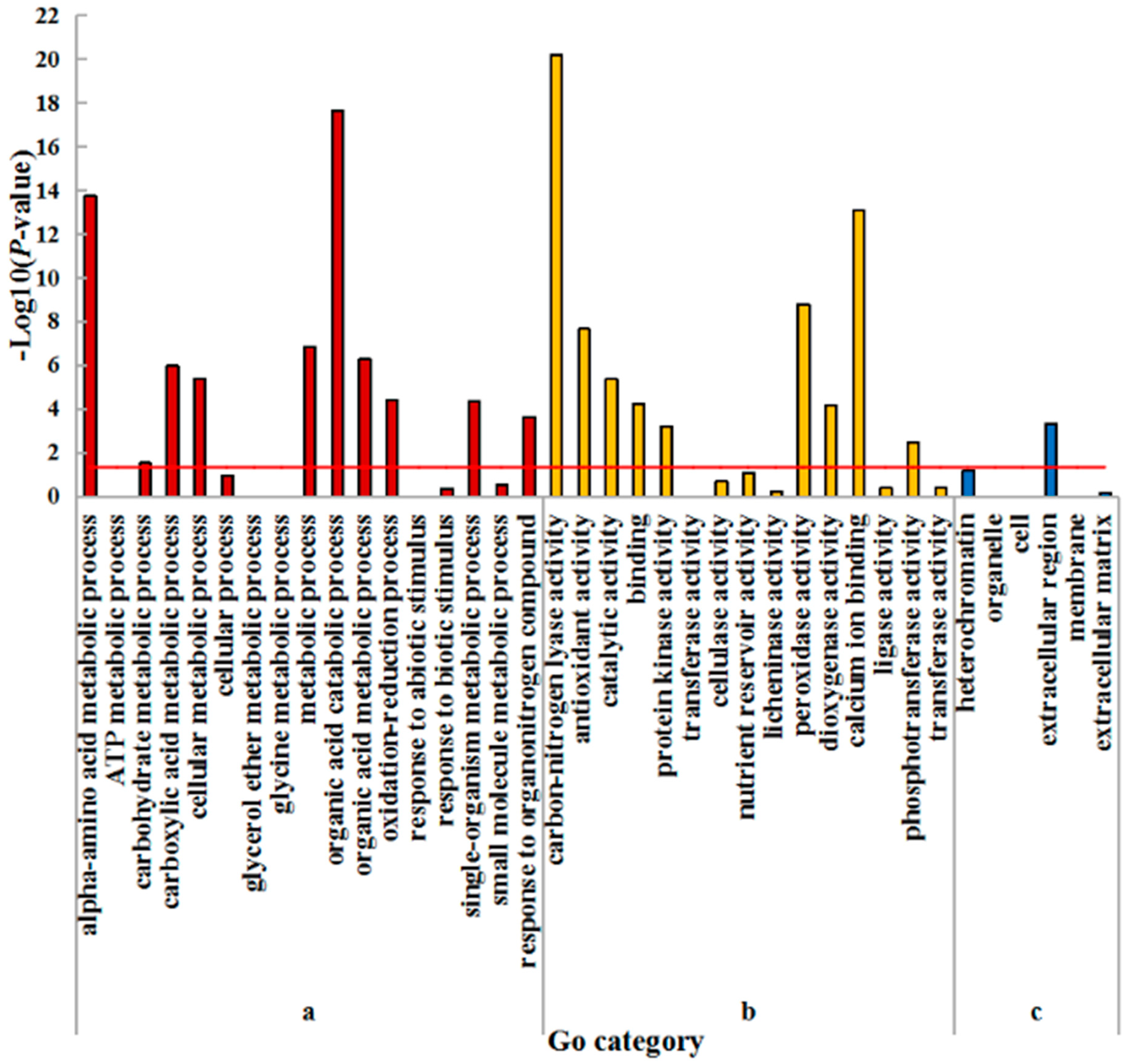
2.4. GO Enrichment Analysis of DEGs
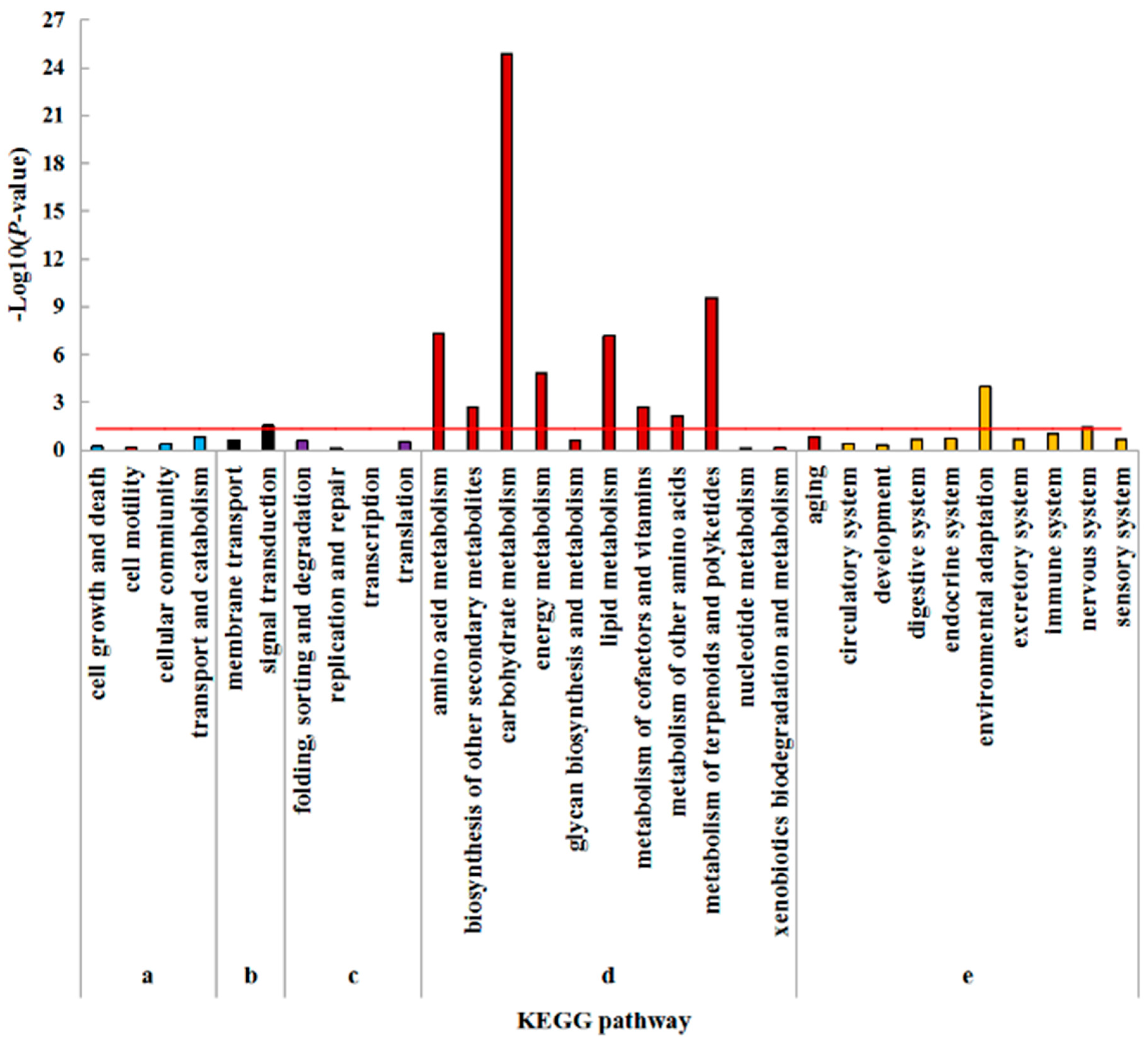
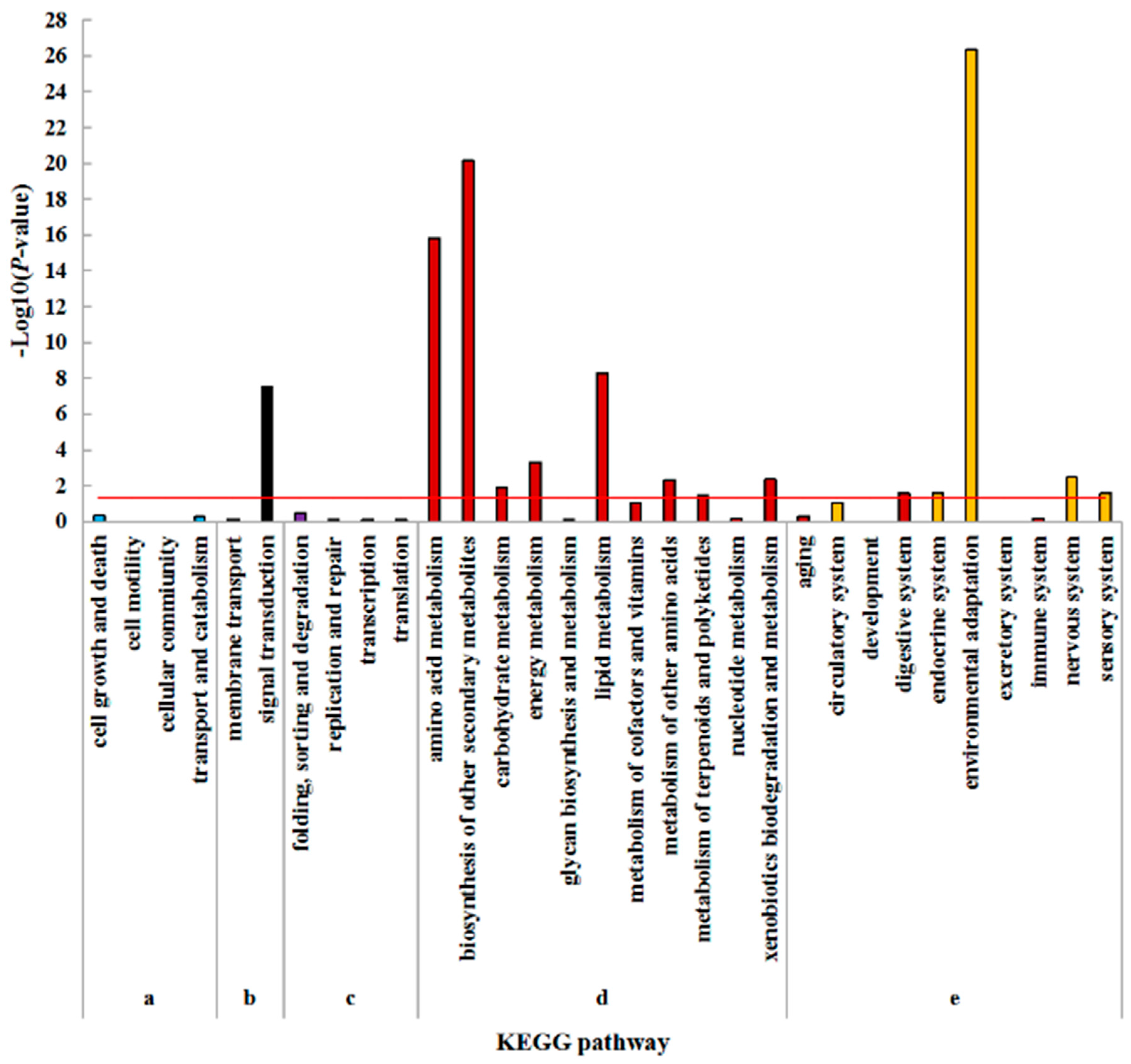
2.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of DEGs
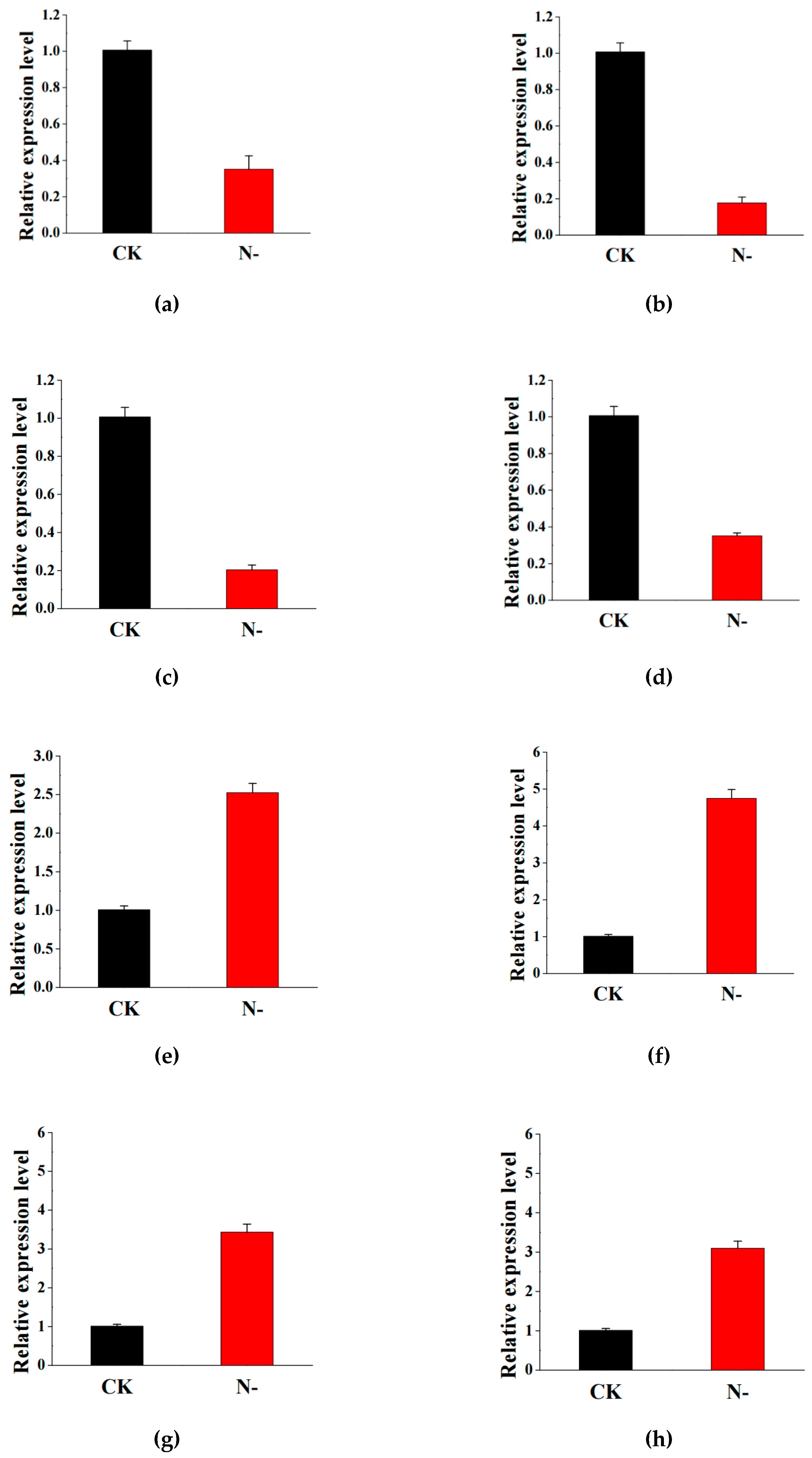
2.6. Validation of RNA Sequencing Data by Quantitative Real-Time PCR
3. Discussion
3.1. Response to Low Nitrogen Stress by Morphological Changes
3.2. Potential DEGs Play Important Roles in Low Nitrogen Tolerance in Wheat Seedlings
3.3. Amino Acid Metabolism, Lipid Metabolism, Energy Metabolism, and Signal Transduction Pathway Play Important Roles Under Low Nitrogen Stress
3.4. Some Candidate Genes for Plant Low Nitrogen Stress Tolerance Breeding
4. Material and Methods
4.1. Plant Material and Growth Conditions
4.2. Experimental Design
4.3. Determination of Morphological Parameters
4.4. Isolation of Total RNA, cDNA Library Construction, and Illumina Sequencing
4.5. Data Filtering and Mapping of Illumina Reads
4.6. Enrichment Analysis of DEGs
4.7. Quantitative Real-Time PCR Validation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Curci, P.L.; Bergès, H.; Marande, W.; Maccaferri, M.; Tuberosa, R.; Sonnante, G. Asparagine synthetase genes (AsnS1, and AsnS2) in durum wheat: Structural analysis and expression under nitrogen stress. Euphytica 2018, 214, 36. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, Y.; Yang, W.; Miao, G. Biological response of roots in different spring wheat genotypes to low nitrogen stress. Acta Agron. Sin. 2006, 32, 1349–1354. [Google Scholar]
- Frink, C.R.; Waggoner, P.E.; Ausubel, J.H. Nitrogen fertilizer: Retrospect and prospect. Proc. Natl. Acad. Sci. USA 1999, 96, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.O.; Simpson, R.J.; Moore, A.D. Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 2006, 286, 7–19. [Google Scholar] [CrossRef]
- Gruber, B.D.; Giehl, R.F.; Friedel, S.; Von, W.N. Plasticity of the arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Plant morphology and root hydraulics are altered by nutrient deficiency in Pistacia lentiscus (L.). Trees 2006, 20, 334–339. [Google Scholar] [CrossRef]
- Jeuffroy, M.H.; Bouchard, C. Intensity and duration of nitrogen deficiency on wheat grain number. Crop Sci. 1999, 39, 1385–1393. [Google Scholar] [CrossRef]
- Xue, Y.F.; Zhang, W.; Liu, D.Y.; Yue, S.C.; Zou, C.Q. Effects of nitrogen management on root morphology and zinc translocation from root to shoot of winter wheat in the field. Field Crops Res. 2014, 161, 38–45. [Google Scholar] [CrossRef]
- Rose, T.J.; Impa, S.M.; Rose, M.T.; Pariasca-Tanaka, J.; Mori, A.; Heuer, S.; JohnsonBeebout, S.E.; Wissuwa, M. Enhancing phosphorus and zinc acquisition efficiency in rice: A critical review of root traits and their potential utility in rice breeding. Ann. Bot. 2013, 112, 331–345. [Google Scholar] [CrossRef]
- Diao, J.; Liu, H.; Hu, F.; Li, L.; Wang, X.; Gai, C. Transcriptome analysis of immune response in fat greenling (Hexagrammos otakii) against Vibrio harveyi infection. Fish Shellfish Immunol. 2018, 84, 937–947. [Google Scholar] [CrossRef]
- Wan, Y.; King, R.; Mitchell, R.A.C.; Hassani-Pak, K.; Hawkesford, M.J. Spatiotemporal expression patterns of wheat amino acid transporters reveal their putative roles in nitrogen transport and responses to abiotic stress. Sci. Rep. 2017, 7, 5461–5474. [Google Scholar] [CrossRef]
- Dai, Z.W.; Plessis, A.; Vincent, J.; Duchateau, N.; Besson, A.; Dardevet, M.; Prodhomme, D.; Gibon, Y.; Hilbert, G.; Pailloux, M.; et al. Transcriptional and metabolic alternations rebalance wheat grain storage protein accumulation under variable nitrogen and sulfur supply. Plant J. 2015, 83, 326–343. [Google Scholar] [CrossRef]
- Kirkman, M.A.; Miflin, B.J. The nitrate content and amino acid composition of the xylem fluid of spring wheat throughout the growing season. J. Sci. Food Agric. 1979, 30, 653–660. [Google Scholar] [CrossRef]
- Lam, H.M.; Hsieh, M.-H.; Coruzzi, G. Reciprocal regulation of distinct asparagine synthetase genes by light and metabolites in Arabidopsis thaliana. Plant J. 1998, 16, 345–353. [Google Scholar] [CrossRef]
- Todd, J.; Screen, S.; Crowley, J. Identification and characterization of four distinct asparagine synthetase (AsnS) genes in maize (Zea mays L.). Plant Sci. 2008, 175, 799–808. [Google Scholar] [CrossRef]
- Yang, S.Y.; Hao, D.L.; Song, Z.Z.; Yang, G.Z.; Wang, L.; Su, Y.H. RNA-Seq analysis of differentially expressed genes in rice under varied nitrogen supplies. Gene 2015, 555, 305–317. [Google Scholar] [CrossRef]
- Curci, P.L.; Cigliano, R.A.; Zuluaga, D.L.; Janni, M.; Sanseverino, W.; Sonnante, G. Transcriptomic response of durum wheat to nitrogen starvation. Sci. Rep. 2017, 7, 1176–1190. [Google Scholar] [CrossRef]
- Forde, B.; Lorenzo, H.; Powlson, D.S. The nutritional control of root development. Plant Soil 2001, 232, 51–68. [Google Scholar] [CrossRef]
- Werf, A.V.D.; Nuenen, M.V.; Visser, A.J.; Lambers, H. Contribution of physiological and morphological plant traits to a species’ competitive ability at high and low nitrogen supply. Oecologia 1993, 94, 434–440. [Google Scholar] [CrossRef]
- Rga, B.; Mensink, M. The Influence of Nitrogen Availability on Growth Parameters of Fast- and Slow-Growing Perennial Grasses; Blackwell Science: Hoboken, NJ, USA, 1991; pp. 161–168. [Google Scholar]
- Boot, R.G.A.; Mensink, M. Size and morphology of root systems of perennial grasses from contrasting habitats as affected by nitrogen supply. Plant Soil 1990, 129, 291–299. [Google Scholar] [CrossRef]
- Schippers, P.; Olff, H. Biomass partitioning, architecture and turnover of six herbaceous species from habitats with different nutrient supply. Plant Ecol. 2000, 149, 219–231. [Google Scholar] [CrossRef]
- Li, X.J.; Guo, C.J.; Lu, W.J.; Duan, W.W.; Zhao, M.; Ma, C.Y. Expression pattern analysis of zinc finger protein genes in wheat (Triticum aestivum L.) under phosphorus deprivation. J. Int. Agric. 2014, 13, 1621–1633. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Bai, L.; Chaoxing, H.E.; Xianchang, Y.U. Dynamic expression of miRNAs and their targets in the response to drought stress of grafted cucumber seedlings. Hortic. Plant J. 2016, 2, 41–49. [Google Scholar] [CrossRef]
- Jun, H.E.; Ren, Y.; Wang, Y. Root morphological and physiological responses of rice seedlings with different tolerance to cadmium stress. Acta Ecol. Sin. 2011, 31, 522–528. [Google Scholar]
- Li, H.; Hu, T.; Amombo, E.; Fu, J. Transcriptome profilings of two tall fescue (Festuca arundinacea) cultivars in response to lead (Pb) stress. BMC Genom. 2017, 18, 145. [Google Scholar] [CrossRef]
- Bi, Y.M.; Wang, R.L.; Tong, Z.; Rothstein, S.J. Global transcription profiling reveals differential responses to chronic nitrogen stress and putative nitrogen regulatory components in Arabidopsis. BMC Genom. 2007, 8, 281. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Stitt, M. Nitrate regulation of metabolism and growth. Curr. Opin. Plant Biol. 1999, 2, 178–186. [Google Scholar] [CrossRef]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Scheible, W.R.; GonzalezFontes, A.; Lauerer, M.; MullerRober, B.; Caboche, M.; Stitt, M. Nitrate acts as a signal to induce organic acid metabolism and repress starch metabolism in tobacco. Plant Cell 1997, 9, 783–798. [Google Scholar] [CrossRef]
- Gelli, M.; Duo, Y.; Konda, A.; Zhang, C.; Holding, D.; Dweikat, I. Identification of differentially expressed genes between sorghum genotypes with contrasting nitrogen stress tolerance by genome-wide transcriptional profiling. BMC Genom. 2014, 15, 179. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Kieselbach, T.; Sauer, U.H.; Schröder, W.P. Purification, crystallization and preliminary X-ray analysis of PPD6, a PsbP-domain protein from Arabidopsis thaliana. Acta Crystallogr. 2012, 68, 278–280. [Google Scholar]
- Bricker, T.M.; Roose, J.L.; Zhang, P.; Frankel, L.K. The PsbP family of proteins. Photosynth. Res. 2013, 116, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, H.; Lu, Q.; Wen, X.; Chen, F.; Peng, L. PsbP-domain protein1, a nuclear-encoded thylakoid lumenal protein, is essential for photosystem I assembly in Arabidopsis. Plant Cell 2012, 24, 4992–5006. [Google Scholar] [CrossRef] [PubMed]
- Fey, V.; Wagner, R.; Braütigam, K.; Wirtz, M.; Hell, R.; Dietzmann, A. Retrograde plastid redox signals in the expression of nuclear genes for chloroplast proteins of Arabidopsis thaliana. J. Biol. Chem. 2005, 280, 5318–5328. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Wu, W.S.; Chen, B.S. Identifying stress transcription factors using gene expression and TF-gene association data. Bioinform. Biol. Insights 2007, 1, 137–145. [Google Scholar] [CrossRef]
- Qu, B.; He, X.; Wang, J.; Zhao, Y.; Teng, W.; Shao, A. A wheat ccaat box-binding transcription factor increases the grain yield of wheat with less fertilizer input. Plant Physiol. 2015, 167, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, G.; Jia, J.; Liu, X.; Kong, X. Molecular characterization of 60 isolated wheat MYB genes and analysis of their expression during abiotic stress. J. Exp. Bot. 2012, 63, 203–214. [Google Scholar] [CrossRef]
- Gaufichon, L.; Reisdorfcren, M.; Rothstein, S.J. Biological functions of asparagine synthetase in plants. Plant Sci. 2015, 179, 141–153. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, M.; Ishiyama, K.; Kojima, S.; Konishi, N.; Nakano, K.; Kanno, K. Asparagine synthetase1, but not asparagine synthetase2, is responsible for the biosynthesis of asparagine following the supply of ammonium to rice roots. Plant Cell Physiol. 2015, 56, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive Oxygen Species and Root Hairs in Arabidopsis Root Response to Nitrogen, Phosphorus and Potassium Deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.M.; Wang, S.P.; Zhang, J.W.; Feng, Q.; Zhang, L.D.; Fan, A.L. Expression profiles of 10,422 genes at early stage of low nitrogen stress in rice assayed using a cDNA microarray. Plant Mol. Biol. 2006, 60, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.S.; Li, J.; Chen, H.H.; Wang, L.L.; Peng, Z.S.; Hu, X.R. Preliminary identification of the biologic characteristics of Chuanmai 42 under different N, P, K concentration. Southwest China J. Agric. Sci. 2007, 20, 281–285. [Google Scholar]
- Boris, L.; Tomáš, L.; Manschadi, A.M. Arbuscular mycorrhizae modify winter wheat root morphology and alleviate phosphorus deficit stress. Plant Soil Environ. 2018, 64, 47–52. [Google Scholar] [CrossRef]
- Qiu, Z.; Yuan, M.; He, Y.; Li, Y.; Zhang, L. Physiological and transcriptome analysis of He-Ne laser pretreated wheat seedlings in response to drought stress. Sci. Rep. 2017, 7, 6108. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Kenneth, J.; Livak, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Method 2001, 25, 402–408. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Leaf Area (cm2/Plant) | Shoot Dry Weigh (mg/Plant) | Root Dry Weigh (mg/Plant) | Total Root Length (cm/Plant) | Total Root Number |
|---|---|---|---|---|---|---|
| CK | 26.58 ± 0.60 a | 14.70 ± 0.17 a | 30.77 ± 0.27 a | 23.17 ± 0.57 a | 57.83 ± 1.31 a | 8.50 ± 0.26 a |
| N- | 19.67 ± 0.56 b | 10.57 ± 0.24 b | 23.29 ± 0.32 b | 14.37 ± 0.76 b | 33.90 ± 1.25 b | 6.70 ± 0.03 b |
| Sample | Raw Reads | Clean Reads | Q20 (%) | Q30 (%) | GC (%) | Total Mapped | Multiple Mapped | Uniquely Mapped | |
|---|---|---|---|---|---|---|---|---|---|
| Leaf | CK | 45,633,102 | 45,096,752 | 95.24 | 88.76 | 55.38 | 38,073,404 (84.43%) | 3,068,994 (8.06%) | 35,004,410 (91.94%) |
| N- | 41,236,032 | 40,827,914 | 95.53 | 89.35 | 54.62 | 34,306,913 (84.03%) | 2,229,665 (6.50%) | 32,077,248 (93.50%) | |
| Root | CK | 41,250,032 | 40,726,872 | 95.24 | 88.94 | 53.08 | 28,921,761 (71.01%) | 1,953,811 (6.76%) | 26,967,950 (93.24%) |
| N- | 44,308,466 | 43,721,986 | 95.00 | 88.39 | 54.92 | 33,846,419 (77.41%) | 1,908,786 (5.64%) | 31,937,633 (94.36%) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Song, K.; Sun, L.; Qin, Q.; Sun, Y.; Pan, J.; Xue, Y. Morphological and Transcriptome Analysis of Wheat Seedlings Response to Low Nitrogen Stress. Plants 2019, 8, 98. https://doi.org/10.3390/plants8040098
Wang J, Song K, Sun L, Qin Q, Sun Y, Pan J, Xue Y. Morphological and Transcriptome Analysis of Wheat Seedlings Response to Low Nitrogen Stress. Plants. 2019; 8(4):98. https://doi.org/10.3390/plants8040098
Chicago/Turabian StyleWang, Jun, Ke Song, Lijuan Sun, Qin Qin, Yafei Sun, Jianjun Pan, and Yong Xue. 2019. "Morphological and Transcriptome Analysis of Wheat Seedlings Response to Low Nitrogen Stress" Plants 8, no. 4: 98. https://doi.org/10.3390/plants8040098