Evolutionary Analysis of GH3 Genes in Six Oryza Species/Subspecies and Their Expression under Salinity Stress in Oryza sativa ssp. japonica

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of the GH3 Genes
2.3. Phylogenetic Analysis
2.4. Analysis of Gene Structure and Conserved Motifs
2.5. Analysis of Chromosome Locations, Gene Duplication Events, and Ka/Ks Values
2.6. Microsynteny Analysis, Cis-Acting Element Analysis, and Functional Annotation Analysis
2.7. Expression Analysis and Co-Expression Network Analysis of OsGH3 Genes Based on the RNA-seq Datasets from Different Tissues
2.8. Expression Analysis of OsGH3 Genes under Salinity Stress by qRT-PCR
3. Results
3.1. Identification and Classification of the GH3 Gene Family
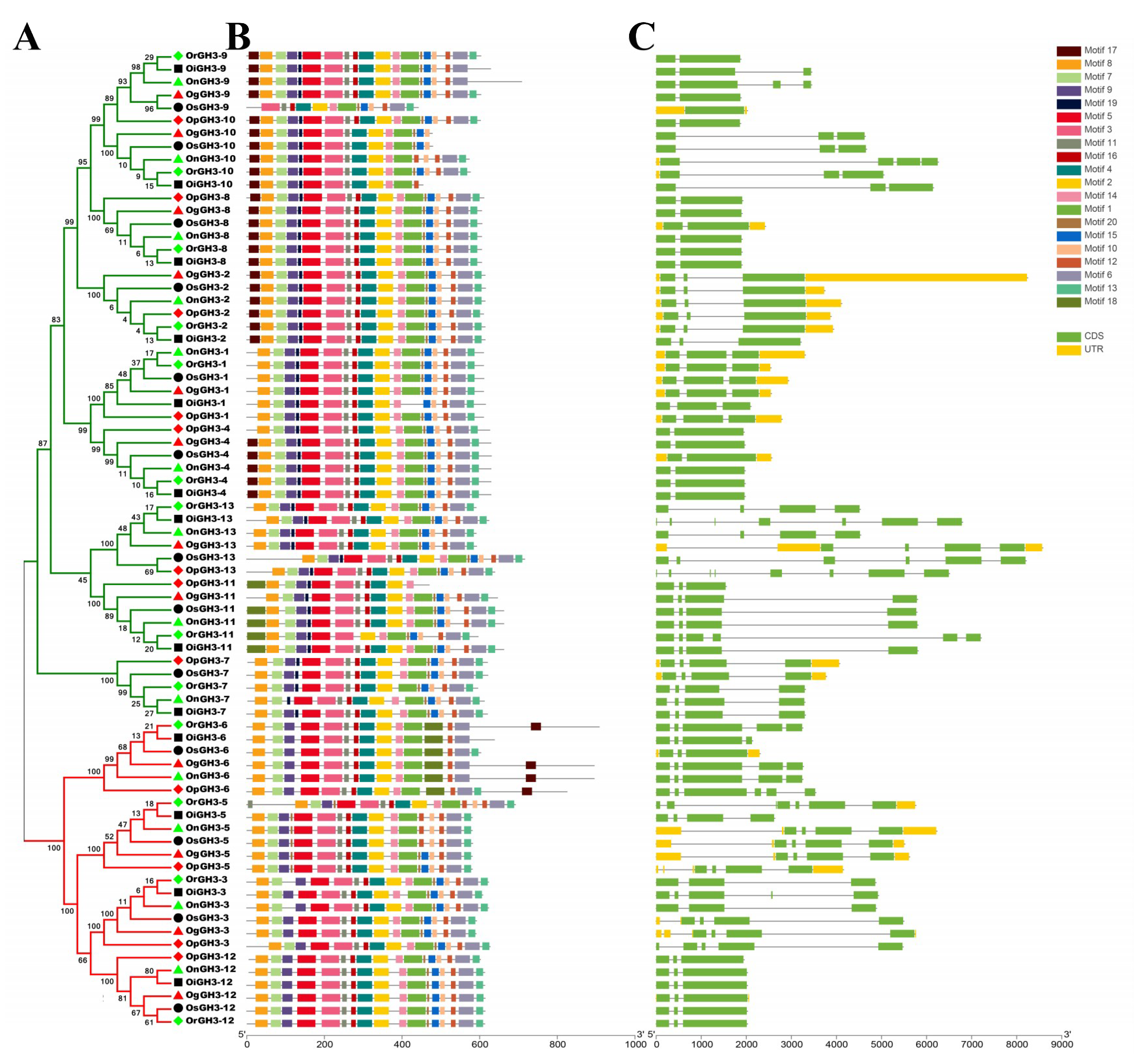
3.2. Gene Structure and Conserved Motif Analysis
3.3. Chromosome Locations, Duplication Events, Selection Pressure, and Microsynteny Analysis
3.4. Analysis of Cis-Elements in OsGH3 Genes
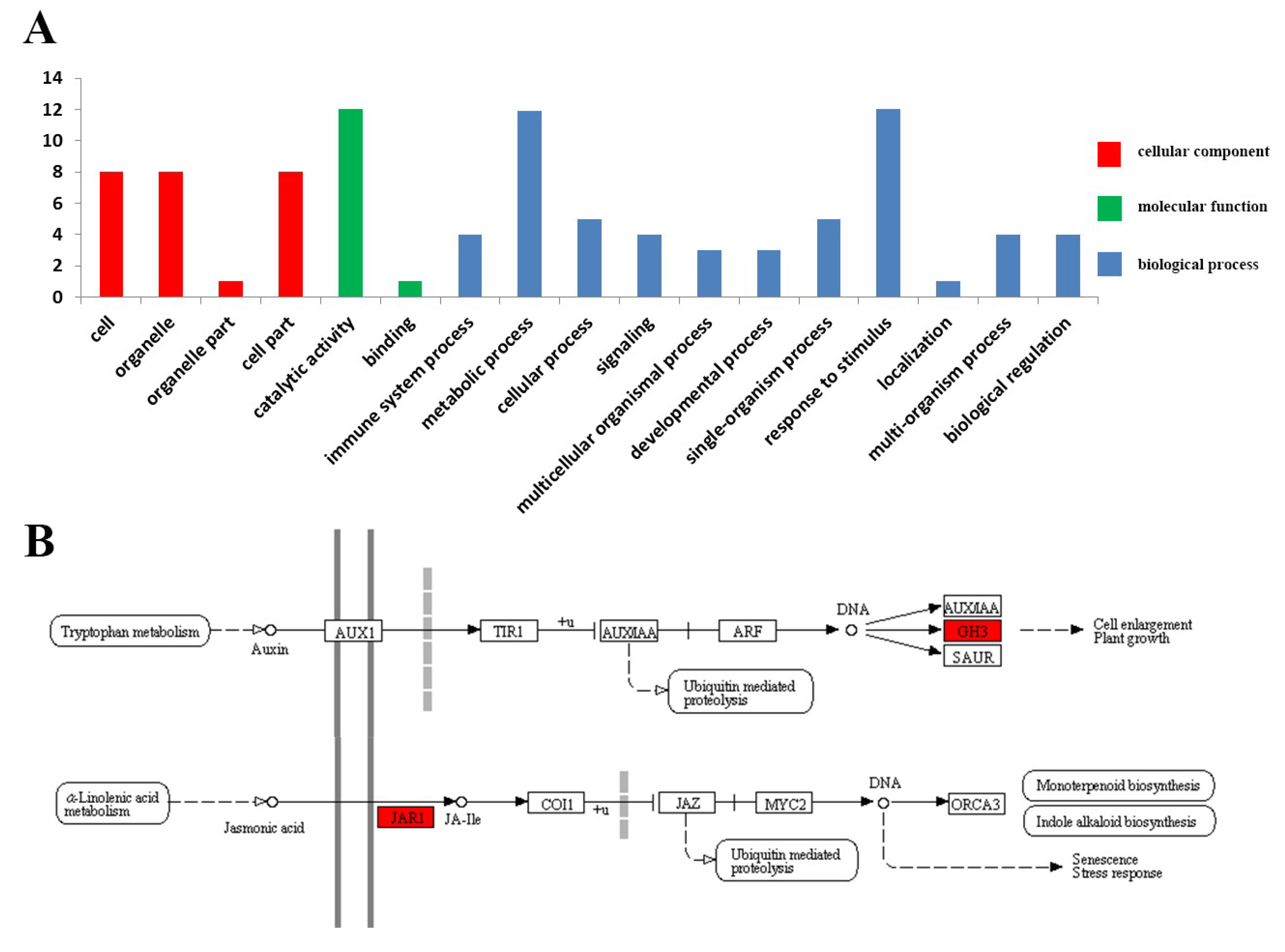
3.5. Functional Annotations Analysis of the OsGH3 Proteins
3.6. Expression Analysis of OsGH3 Genes in Different Tissues and under Salinity Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethics Approval and Consent to Participate
References
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Baranwal, V.; Negi, N.; Khurana, P. Auxin response factor genes repertoire in mulberry: Identification, and structural, functional and evolutionary analyses. Genes 2017, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xi, H.; Cao, J. Small auxin upregulated RNA (SAUR) gene family in maize: Identification, evolution, and its phylogenetic comparison with Arabidopsis, rice, and sorghum. J. Integr. Plant Biol. 2014, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Hagen, G.; Kleinschmidt, A.; Guilfoyle, T. Auxin-regulated gene expression in intact soybean hypocotyl and excised hypocotyl sections. Planta 1984, 162, 147–153. [Google Scholar] [CrossRef]
- Staswick, P.E.; Tiryaki, I.; Rowe, M.L. Jasmonate response locus JAR1 and several related Arabidopsis genes encode enzymes of the firefly luciferase superfamily that show activity on Jasmonic, Salicylic, and Indole-3-acetic acids in an assay for adenylation. Plant Cell 2002, 14, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Agarwal, P.; Tyagi, A.K.; Sharma, A.K. Genome-wide investigation and expression analysis suggest diverse roles of Auxin-responsive GH3 genes during development and response to different stimuli in tomato (Solanum lycopersicum). Mol. Genet. Genom. 2012, 287, 221–235. [Google Scholar] [CrossRef]
- Singh, V.K.; Jain, M.; Garg, R. Genome-wide analysis and expression profiling suggest diverse roles of GH3 genes during development and abiotic stress responses in legumes. Front. Plant. Sci. 2015, 5, 789. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, L.; Wang, D.; Ma, H.; Liu, B.; Shi, Z.; Ma, X.; Chen, Y.; Chen, Q. Evolutionary history of the glycoside hydrolase 3 (GH3) family based on the sequenced genomes of 48 plants and identification of Jasmonic acid-related GH3 proteins in Solanum tuberosum. Int. J. Mol. Sci. 2018, 19, 1850. [Google Scholar] [CrossRef]
- Okrent, R.A.; Brooks, M.D.; Wildermuth, M.C. Arabidopsis GH3.12 (PBS3) conjugates amino acids to 4-substituted benzoates and is inhibited by Salicylate. J. Biol. Chem. 2009, 284, 9742–9754. [Google Scholar] [CrossRef]
- Feng, S.; Yue, R.; Tao, Y.; Yang, Y.; Zhang, L.; Xu, M.; Wang, H.; Shen, C. Genome-wide identification, expression analysis of Auxin-responsive GH3 family genes in maize (Zea mays L.) under abiotic stresses. Acta Bot. Sin. 2015, 57, 783–795. [Google Scholar]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The Auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Terol, J.; Domingo, C.; Talón, M. The GH3 family in plants: Genome wide analysis in rice and evolutionary history based on EST analysis. Gene 2006, 371, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Yu, H.; Li, X.; Xiao, J.; Wang, S. Rice GH3 gene family: Regulators of growth and development. Plant Signal. Behav. 2011, 6, 570–574. [Google Scholar] [CrossRef]
- Fu, J.; Liu, H.; Li, Y.; Yu, H.; Li, X.; Xiao, J.; Wang, S. Manipulating broad-spectrum disease resistance by suppressing pathogen-induced Auxin accumulation in rice. Plant Signal. Behav. 2011, 155, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Cao, Y.; Huang, L.; Zhao, J.; Xu, C.; Li, X.; Wang, S. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes Salicylate- and Jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Andrés, F.; Tharreau, D.; Iglesias, D.J.; Talón, M. Constitutive expression of OsGH3.1 reduces Auxin content and enhances defense response and resistance to a fungal pathogen in rice. Mol. Plant Microbe Interact. 2009, 22, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, C.; Cao, J.; Zhang, Y.; Zhang, S.; Xia, Y.; Sun, D.; Sun, Y. Altered architecture and enhanced drought tolerance in rice via the down-regulation of Indole-3-acetic acid by TLD1/OsGH3.13 activation. Plant Physiol. 2009, 151, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 family member, OsGH3-2, modulates Auxin and Abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Han, S.; Yoon, E.; Lee, W. Evidence of an Auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous Auxin in cultured rice cells. Nucleic Acids Res. 2006, 34, 1892–1899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nallamilli, B.R.; Mujahid, H.; Peng, Z.H. OsMADS6 plays an essential role in endosperm nutrient accumulation and is subject to epigenetic regulation in rice (Oryza sativa). Plant J. 2010, 64, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.R.; Khanday, I.; Majhi, B.B.; Veluthambi, K.; Vijayraghavan, U. Auxin-responsive OsMGH3, a common downstream target of OsMADS1 and OsMADS6, controls rice floret fertility. Plant Cell Physiol. 2011, 52, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wang, J.; Yang, X.; Lu, H.; Miao, X.; Shi, Z. Modulation of plant architecture by the miR156f–OsSPL7–OsGH3. 8 pathway in rice. J. Exp. Bot. 2018, 69, 5117–5130. [Google Scholar] [CrossRef]
- Rafii, M.Y.; Kalhori, N.; Hanafi, M.M.; Sahebi, M.; Mahmud, T.M.M.; Abiri, R.; Azizi, P.; Taheri, S.; Atabaki, N.; Shabanimofrad, M. Improvement of drought tolerance in rice (Oryza sativa L.): Genetics, genomic tools, and the WRKY gene family. Biomed. Res. Int. 2018, 4, 1–20. [Google Scholar]
- Lavania, D.; Dhingra, A.; Grover, A. Analysis of transactivation potential of rice (Oryza sativa L.) heat shock factors. Planta 2018, 247, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brozynska, M.; Copetti, D.; Furtado, A.; Wing, R.A.; Crayn, D.; Fox, G.; Ishikawa, R.; Henry, R.J. Sequencing of Australian wild rice genomes reveals ancestral relationships with domesticated rice. Plant Biotechnol. J. 2017, 15, 765–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, D.A.; Morishima, H.; Kadowaki, K. Diversity in the Oryza genus. Curr. Opin. Plant Biol. 2003, 6, 139–146. [Google Scholar] [CrossRef]
- Kong, W.; Yang, S.; Wang, Y.; Bendahmane, M.; Fu, X. Genome-wide identification and characterization of aquaporin gene family in Beta vulgaris. PeerJ 2017, 5, e3747. [Google Scholar] [CrossRef]
- Cao, Y.; Meng, D.; Chen, Y.; Abdullah, M.; Jin, Q.; Lin, Y.; Cai, Y. Comparative and expression analysis of ubiquitin conjugating domain-containing genes in two Pyrus species. Cells 2018, 7, 77. [Google Scholar] [CrossRef]
- Kong, W.; Bendahmane, M.; Fu, X. Genome-wide identification and characterization of aquaporins and their role in the flower opening processes in carnation (Dianthus caryophyllus). Molecules 2018, 23, 1895. [Google Scholar] [CrossRef]
- Verma, J.; Wardhan, V.; Singh, D.; Chakraborty, S.; Chakraborty, N. Genome-wide identification of the Alba gene family in plants and stress-responsive expression of the rice Alba genes. Genes 2018, 9, 183. [Google Scholar] [CrossRef]
- Mohanta, T.; Mohanta, N.; Bae, H. Identification and expression analysis of PIN-like (PILS) gene family of rice treated with Auxin and Cytokinin. Genes 2015, 6, 622–640. [Google Scholar] [CrossRef]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Hu, W.; Yue, W.; Liu, B.; Yan, H.; Yan, X. Genome-wide identification, classification, and expression of phytocyanins in Populus trichocarpa. Planta 2018, 247, 1133–1148. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopaedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Osakabe, Y.; Qin, F.; Simpson, S.D.; Maruyama, K.; Fujita, Y.; Shinozaki, K.; Yamaguchishinozaki, K. Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis. Plant J. 2010, 49, 46–63. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, e11. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Cao, Y.; Cheng, X.; Meng, D.; Chen, Y.; Shakoor, A.; Gao, J.; Cai, Y. The sucrose synthase gene family in chinese pear (Pyrus bretschneideri Rehd.): Structure, expression, and evolution. Molecules 2018, 23, 1144. [Google Scholar] [CrossRef] [PubMed]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; Berger, F. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 2017, 171, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Bierfreund, N.M.; Tintelnot, S.; Reski, R.; Decker, E.L. Loss of GH3 function does not affect phytochrome-mediated development in a moss, Physcomitrella patens. J. Plant Physiol. 2004, 161, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zou, X.H.; Achoundong, G.; Potgieter, C.; Second, G.; Zhang, D.Y.; Ge, S. Phylogeny and biogeography of the rice tribe (Oryzeae): Evidence from combined analysis of 20 chloroplast fragments. Mol. Phylogenet. Evol. 2010, 54, 266–277. [Google Scholar] [CrossRef]
- Stein, J.C.; Yu, Y.; Copetti, D.; Zwickl, D.J.; Zhang, L.; Zhang, C.; Chougule, K.; Gao, D.; Iwata, A.; Goicoechea, J.L. Genomes of 13 domesticated and wild rice relatives highlight genetic conservation, turnover and innovation across the genus Oryza. Nat. Genet. 2018, 7, 285–296. [Google Scholar] [CrossRef]
- Choi, J.Y.; Platts, A.E.; Fuller, D.Q.; Hsing, Y.I.; Wing, R.A.; Purugganan, M.D. The rice paradox: Multiple origins but single domestication in Asian rice. Mol. Biol. Evol. 2017, 34, 969–979. [Google Scholar] [CrossRef]
- Kim, K.; Lee, S.-C.; Lee, J.; Yu, Y.; Yang, K.; Choi, B.-S.; Koh, H.-J.; Waminal, N.E.; Choi, H.-I.; Kim, N.-H. Complete chloroplast and ribosomal sequences for 30 accessions elucidate evolution of Oryza AA genome species. Sci. Rep. 2015, 5, 15655. [Google Scholar] [CrossRef]
- Kovach, M.J.; Sweeney, M.T.; Mccouch, S.R. New insights into the history of rice domestication. Trends Genet. 2007, 23, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Londo, J.P.; Yu-Chung, C.; Kuo-Hsiang, H.; Tzen-Yuh, C.; Schaal, B.A. Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Natl. Acad. Sci. USA 2006, 103, 9578–9583. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Qiao, W.H.; Chen, Y.T.; Wang, R.S.; Cao, L.R.; Zhang, W.X.; Yuan, N.N.; Li, Z.C.; Zeng, H.L.; Yang, Q.W. Domestication and geographic origin of Oryza sativa in China: Insights from multilocus analysis of nucleotide variation of O. sativa and O. rufipogon. Mol. Ecol. 2012, 21, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yue, R.; Sun, T.; Zhang, L.; Chen, W.; Zeng, H.; Wang, H.; Shen, C. Genome-wide identification, expression analysis of GH3 family genes in Medicago truncatula under stress-related hormones and Sinorhizobium meliloti infection. Appl. Microbiol. Biotechnol. 2015, 99, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bai, Y.; Shen, C.; Wu, Y.; Zhang, S.; Jiang, D.; Guilfoyle, T.J.; Chen, M.; Qi, Y. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Name | Gene Identifier | CHR | CHR.start | CHR.end | Subfamily |
|---|---|---|---|---|---|---|
| Oryza sativa ssp. japonica | OsGH3-3 | LOC_Os01g12160.1 | Oschr1 | 6,624,963 | 6,630,462 | Group1 |
| OsGH3-5 | LOC_Os05g50890.1 | Oschr5 | 29,200,198 | 29,205,717 | Group1 | |
| OsGH3-6 | LOC_Os05g05180.2 | Oschr5 | 2,522,903 | 2,525,214 | Group1 | |
| OsGH3-12 | LOC_Os11g08340.1 | Oschr11 | 4,401,492 | 4,403,522 | Group1 | |
| OsGH3-1 | LOC_Os01g57610.1 | Oschr1 | 33,308,448 | 33,311,391 | Group2 | |
| OsGH3-2 | LOC_Os01g55940.1 | Oschr1 | 32,221,376 | 32,225,123 | Group2 | |
| OsGH3-4 | LOC_Os05g42150.1 | Oschr5 | 24,643,516 | 24,646,086 | Group2 | |
| OsGH3-7 | LOC_Os06g30440.1 | Oschr6 | 17,586,899 | 17,590,682 | Group2 | |
| OsGH3-8 | LOC_Os07g40290.1 | Oschr7 | 24,149,649 | 24,152,079 | Group2 | |
| OsGH3-9 | LOC_Os07g38890.1 | Oschr7 | 23,325,197 | 23,327,227 | Group2 | |
| OsGH3-10 | LOC_Os07g38860.1 | Oschr7 | 23,314,482 | 23,319,154 | Group2 | |
| OsGH3-11 | LOC_Os07g47490.1 | Oschr7 | 28,391,725 | 28,400,307 | Group2 | |
| OsGH3-13 | LOC_Os11g32520.1 | Oschr11 | 19,188,565 | 19,190,125 | Group2 | |
| Oryza sativa ssp. indica | OiGH3-3 | BGIOSGA002109-PA | Oichr1 | 7,084,891 | 7,089,830 | Group1 |
| OiGH3-5 | BGIOSGA020457-PA | Oichr5 | 30,569,921 | 30,572,557 | Group1 | |
| OiGH3-6 | BGIOSGA018825-PA | Oichr5 | 2,768,250 | 2,770,394 | Group1 | |
| OiGH3-12 | BGIOSGA034955-PA | Oichr11 | 4,242,522 | 4,244,552 | Group1 | |
| OiGH3-1 | BGIOSGA004585-PA | Oichr1 | 36,754,839 | 36,756,946 | Group2 | |
| OiGH3-2 | BGIOSGA004510-PA | Oichr1 | 35,535,345 | 35,538,565 | Group2 | |
| OiGH3-4 | BGIOSGA017778-PA | Oichr5 | 26,098,760 | 26,100,739 | Group2 | |
| OiGH3-7 | BGIOSGA021194-PA | Oichr6 | 18,602,739 | 18,606,054 | Group2 | |
| OiGH3-8 | BGIOSGA023979-PA | Oichr7 | 22,260,570 | 22,262,481 | Group2 | |
| OiGH3-9 | BGIOSGA024029-PA | Oichr7 | 21,410,892 | 21,414,347 | Group2 | |
| OiGH3-10 | BGIOSGA025998-PA | Oichr7 | 21,402,461 | 21,402,912 | Group2 | |
| OiGH3-11 | BGIOSGA023736-PA | Oichr7 | 26,392,482 | 26,397,014 | Group2 | |
| OiGH3-13 | BGIOSGA03388PA | Oichr11 | 15,962,369 | 15,968,186 | Group2 | |
| Oryza rufipogon | OrGH3-3 | ORUFI01G08270.1 | Orchr1 | 6,148,966 | 6,153,847 | Group1 |
| OrGH3-5 | ORUFI05G29440.1 | Orchr5 | 25,858,361 | 25,864,126 | Group1 | |
| OrGH3-6 | ORUFI05G03070.1 | Orchr5 | 2,229,725 | 2,232,983 | Group1 | |
| OrGH3-12 | ORUFI11G05180.1 | Orchr11 | 4,022,556 | 4,024,586 | Group1 | |
| OrGH3-1 | ORUFI01G36550.1 | Orchr1 | 30,644,226 | 30,646,778 | Group2 | |
| OrGH3-2 | ORUFI01G35260.1 | Orchr1 | 29,632,340 | 29,636,282 | Group2 | |
| OrGH3-4 | ORUFI05G23110.1 | Orchr5 | 21,592,626 | 21,594,605 | Group2 | |
| OrGH3-7 | ORUFI06G16960.1 | Orchr6 | 16,046,446 | 16,049,761 | Group2 | |
| OrGH3-8 | ORUFI07G21680.1 | Orchr7 | 21,004,765 | 21,006,676 | Group2 | |
| OrGH3-9 | ORUFI07G20520.1 | Orchr7 | 20,243,069 | 20,244,953 | Group2 | |
| OrGH3-10 | ORUFI07G20500.1 | Orchr7 | 20,233,907 | 20,238,965 | Group2 | |
| OrGH3-11 | ORUFI07G26860.1 | Orchr7 | 24,893,286 | 24,900,088 | Group2 | |
| OrGH3-13 | ORUFI11G16590.1 | Orchr11 | 19,413,084 | 19,420,302 | Group2 | |
| Oryza nivara | OnGH3-3 | ONIVA01G09800.1 | Onchr1 | 7,585,019 | 7,589,913 | Group1 |
| OnGH3-5 | ONIVA05G29530.1 | Onchr5 | 27,396,671 | 27,402,904 | Group1 | |
| OnGH3-6 | ONIVA05G02860.1 | Onchr5 | 2,172,465 | 2,175,724 | Group1 | |
| OnGH3-12 | ONIVA11G05730.1 | Onchr11 | 4,716,885 | 4,718,915 | Group1 | |
| OnGH3-1 | ONIVA01G38150.1 | Onchr1 | 32,674,099 | 32,677,412 | Group2 | |
| OnGH3-2 | ONIVA01G36390.1 | Onchr1 | 31,322,609 | 31,326,729 | Group2 | |
| OnGH3-4 | ONIVA05G22520.1 | Onchr5 | 22,188,295 | 22,190,274 | Group2 | |
| OnGH3-7 | ONIVA06G18950.1 | Onchr6 | 17,364,605 | 17,367,911 | Group2 | |
| OnGH3-8 | ONIVA07G19200.1 | Onchr7 | 18,453,792 | 18,455,703 | Group2 | |
| OnGH3-9 | ONIVA07G18070.1 | Onchr7 | 17,588,411 | 17,591,865 | Group2 | |
| OnGH3-10 | ONIVA07G18060.1 | Onchr7 | 17,579,620 | 17,585,886 | Group2 | |
| OnGH3-11 | ONIVA07G25530.1 | Onchr7 | 23,284,878 | 23,289,419 | Group2 | |
| OnGH3-13 | ONIVA11G14940.1 | Onchr11 | 16,487,974 | 16,493,787 | Group2 | |
| Oryza punctata | OpGH3-3 | OPUNC01G07310.1 | Opchr1 | 6,025,916 | 6,031,401 | Group1 |
| OpGH3-5 | OPUNC05G25060.1 | Opchr5 | 30,476,518 | 30,480,678 | Group1 | |
| OpGH3-6 | OPUNC05G02820.1 | Opchr5 | 2,336,689 | 2,340,237 | Group1 | |
| OpGH3-12 | OPUNC11G05160.1 | Opchr11 | 4,742,463 | 4,744,418 | Group1 | |
| OpGH3-1 | OPUNC01G32450.1 | Opchr1 | 35,758,425 | 35,761,223 | Group2 | |
| OpGH3-2 | OPUNC01G31080.1 | Opchr1 | 34,433,992 | 34,437,877 | Group2 | |
| OpGH3-4 | OPUNC05G19440.1 | Opchr5 | 26,021,393 | 26,023,350 | Group2 | |
| OpGH3-7 | OPUNC06G12520.1 | Opchr6 | 13,614,325 | 13,618,401 | Group2 | |
| OpGH3-8 | OPUNC07G19610.1 | Opchr7 | 26,181,745 | 26,183,673 | Group2 | |
| OpGH3-9 | OPUNC07G18650.1 | Opchr7 | 25,197,946 | 25,199,823 | Group2 | |
| OpGH3-11 | OPUNC07G24330.1 | Opchr7 | 29,891,760 | 29,898,273 | Group2 | |
| OpGH3-13 | OPUNC11G12860.1 | Opchr11 | 20,394,638 | 20,400,442 | Group2 | |
| Oryza glumaepatula | OgGH3-3 | OGLUM01G08700.1 | Ogchr1 | 7,724,249 | 7,730,000 | Group1 |
| OgGH3-5 | OGLUM05G28990.1 | Ogchr5 | 29,839,316 | 29,844,943 | Group1 | |
| OgGH3-6 | OGLUM05G02960.1 | Ogchr5 | 2,520,030 | 2,523,301 | Group1 | |
| OgGH3-12 | OGLUM11G05200.1 | Ogchr11 | 4,202,889 | 4,204,936 | Group1 | |
| OgGH3-1 | OGLUM01G37610.1 | Ogchr1 | 36,364,851 | 36,367,411 | Group2 | |
| OgGH3-2 | OGLUM01G36180.1 | Ogchr1 | 35,099,335 | 35,107,582 | Group2 | |
| OgGH3-4 | OGLUM05G23040.1 | Ogchr5 | 25,041,853 | 25,043,832 | Group2 | |
| OgGH3-8 | OGLUM07G20630.1 | Ogchr7 | 23,059,911 | 23,061,822 | Group2 | |
| OgGH3-9 | OGLUM07G19490.1 | Ogchr7 | 22,131,580 | 22,133,468 | Group2 | |
| OgGH3-10 | OGLUM07G19470.1 | Ogchr7 | 22,122,128 | 22,126,771 | Group2 | |
| OgGH3-11 | OGLUM07G25930.1 | Ogchr7 | 27,048,553 | 27,056,767 | Group2 | |
| OgGH3-13 | OGLUM11G14980.1 | Ogchr11 | 18,442,620 | 18,448,411 | Group2 |
| Seq1 | Seq2 | Ks | Ka | Ka/Ks Ratio | Date (MY) | Duplication Type |
|---|---|---|---|---|---|---|
| OiGH3-1 | OiGH3-4 | 0.5644 | 0.207 | 0.367 | 31.010989 | WGD or segmental duplication |
| OsGH3-1 | OsGH3-4 | 0.4274 | 0.1251 | 0.293 | 23.483516 | WGD or segmental duplication |
| OgGH3-1 | OgGH3-4 | 0.4223 | 0.1273 | 0.301 | 23.203297 | WGD or segmental duplication |
| OnGH3-1 | OnGH3-4 | 0.4237 | 0.1269 | 0.300 | 23.28022 | WGD or segmental duplication |
| OpGH3-1 | OpGH3-4 | 0.4754 | 0.1245 | 0.262 | 26.120879 | WGD or segmental duplication |
| OrGH3-1 | OrGH3-4 | 0.4198 | 0.1252 | 0.298 | 23.065934 | WGD or segmental duplication |
| OiGH3-10 | OiGH3-9 | 0.2407 | 0.1365 | 0.567 | 13.225275 | Tandem duplication |
| OnGH3-10 | OnGH3-9 | 0.25 | 0.1274 | 0.510 | 13.736264 | Tandem duplication |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, W.; Zhong, H.; Deng, X.; Gautam, M.; Gong, Z.; Zhang, Y.; Zhao, G.; Liu, C.; Li, Y. Evolutionary Analysis of GH3 Genes in Six Oryza Species/Subspecies and Their Expression under Salinity Stress in Oryza sativa ssp. japonica. Plants 2019, 8, 30. https://doi.org/10.3390/plants8020030
Kong W, Zhong H, Deng X, Gautam M, Gong Z, Zhang Y, Zhao G, Liu C, Li Y. Evolutionary Analysis of GH3 Genes in Six Oryza Species/Subspecies and Their Expression under Salinity Stress in Oryza sativa ssp. japonica. Plants. 2019; 8(2):30. https://doi.org/10.3390/plants8020030
Chicago/Turabian StyleKong, Weilong, Hua Zhong, Xiaoxiao Deng, Mayank Gautam, Ziyun Gong, Yue Zhang, Gangqing Zhao, Chang Liu, and Yangsheng Li. 2019. "Evolutionary Analysis of GH3 Genes in Six Oryza Species/Subspecies and Their Expression under Salinity Stress in Oryza sativa ssp. japonica" Plants 8, no. 2: 30. https://doi.org/10.3390/plants8020030