Transcriptomic Analysis of Dark-Induced Senescence in Bermudagrass (Cynodon dactylon)


Abstract
:1. Introduction
2. Results
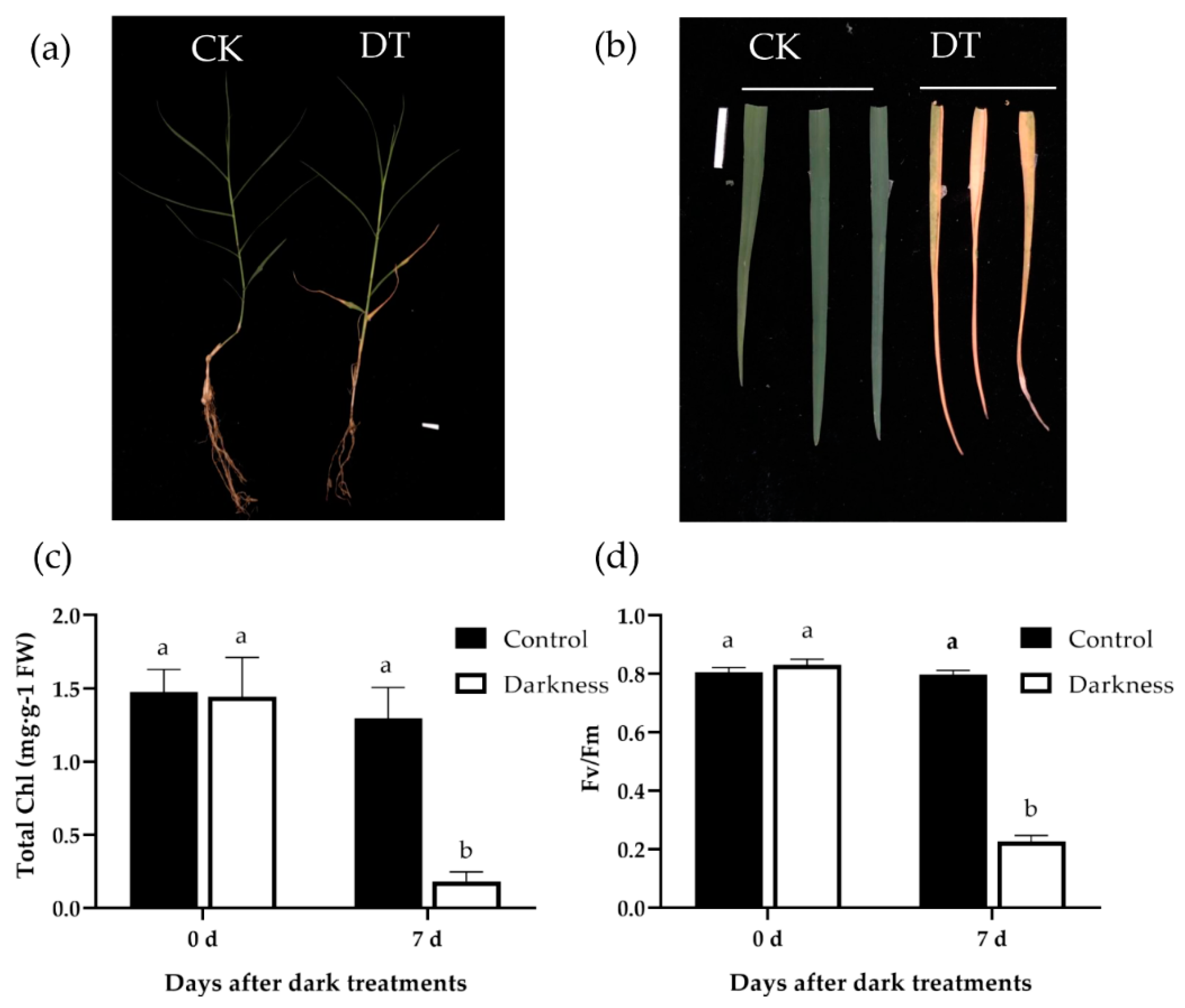
2.1. Dark Induced Leaf Senescence in Bermudagrass
2.2. Characteristics of Genes in Response to Dark-Induced Leaf Senescence in Bermudagrass
2.3. Gene Annotation of the Bermudagrass Transcriptome
2.4. Differentially Expressed Genes (DEGs) in Response to Dark-Induced Leaf Senescence in Bermudagrass
2.5. GO Analysis of Differentially Expressed Genes between CK and DT
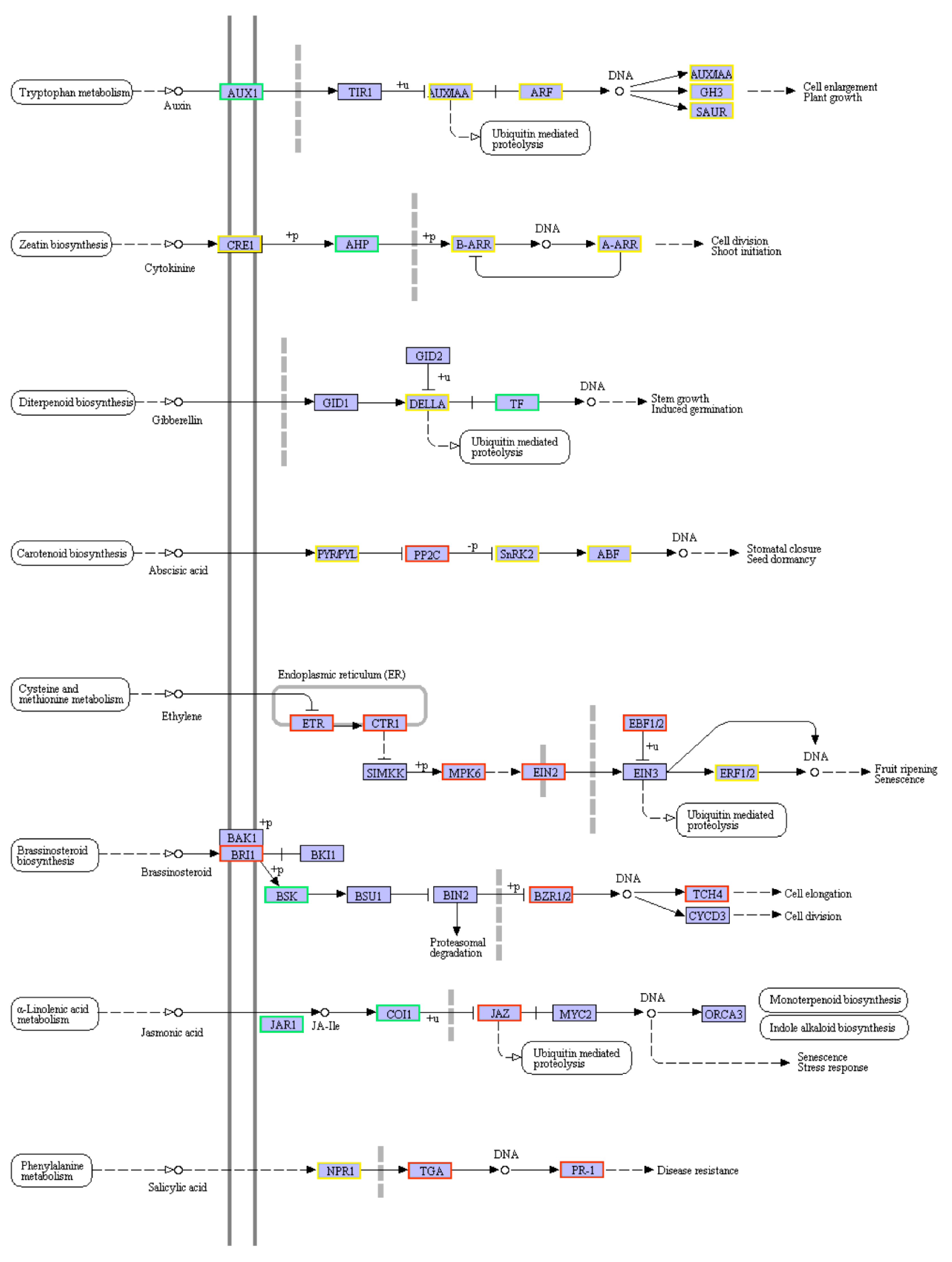
2.6. KEGG Pathway Enrichment Analysis of DEGs between CK and DT
2.7. Genes Responsive to Dark-Induced Leaf Senescence in Bermudagrass
2.8. Regulation of Transcription Factor (TF) Families during Dark-Induced Leaf Senescence
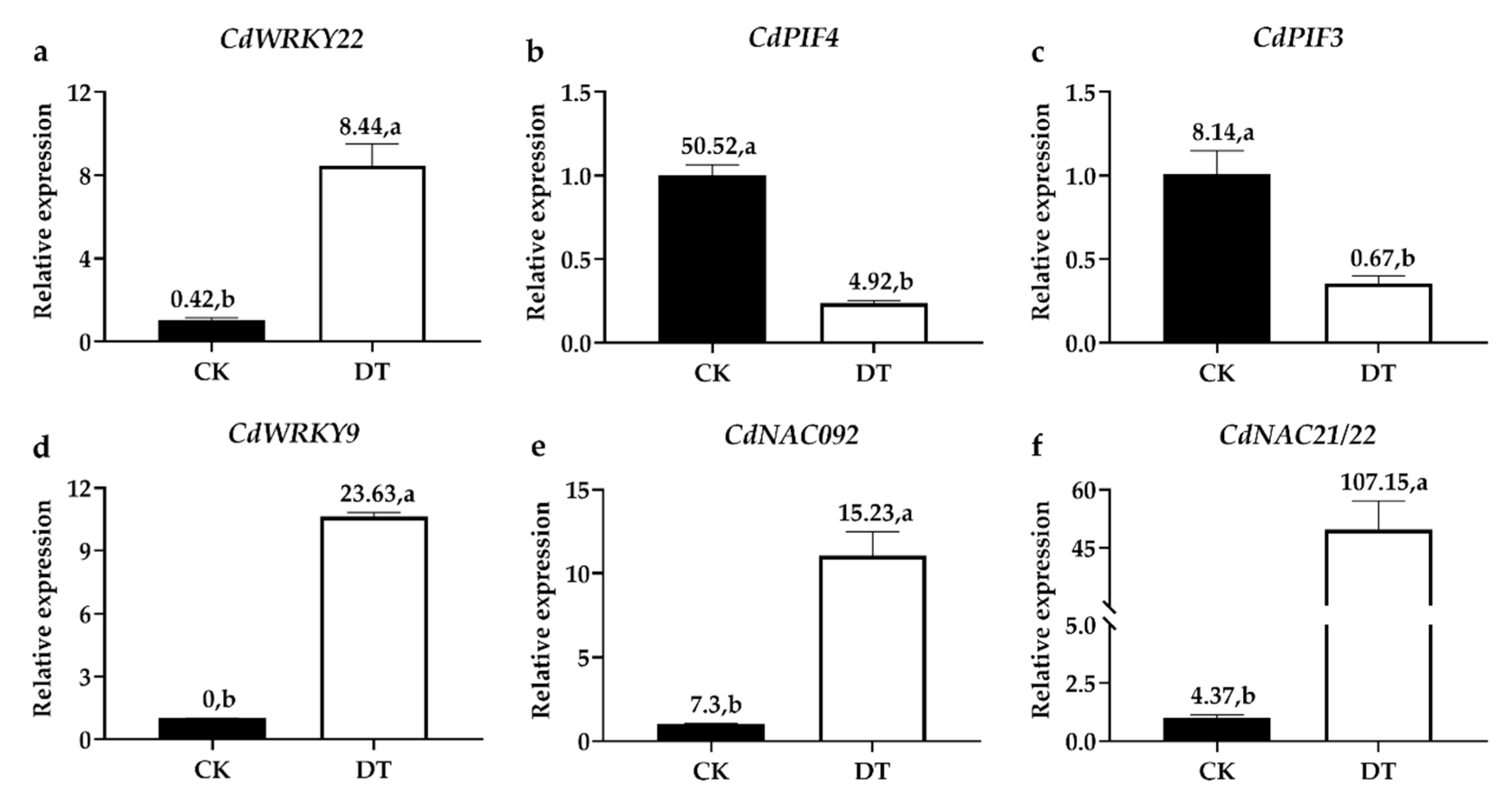
2.9. Validating the DEGs by qRT-PCR Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Treatments
5.2. RNA Preparation
5.3. Library Preparation for Transcription Sequencing
5.4. Data Analysis
5.4.1. Quality Control
5.4.2. Transcriptome Assembly
5.4.3. Gene Functional Annotation
5.4.4. Differential Expression Analysis
5.4.5. GO Enrichment Analysis
5.4.6. KEGG Pathway Enrichment Analysis
5.5. Validation of RNA-Sequence Data by Quantitative Real-Time PCR (qRT-PCR)
5.6. Measurements of Chlorophyll Content and Fv/Fm
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Peterson, K.W.; Fry, J.D.; Bremer, D.J. Growth Responses of Zoysia spp. under Tree Shade in the Midwestern United States. HortScience 2014, 49, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Tegg, R.S.; Lane, P.A. A comparison of the performance and growth of a range of turfgrass species under shade. Aust. J. Exp. Agric. 2004, 44, 353–358. [Google Scholar] [CrossRef]
- Chin, S.-W. Comparative Performance of Three Tropical Turfgrasses Digitaria longiflora, Axonopus compressus and St. Augustinegrass under Simulated Shade Conditions. Weed Turfgrass Sci. 2017, 6, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Lim, P.O.; Woo, H.R.; Gil Nam, H. Molecular genetics of leaf senescence in Arabidopsis. Trends Plant Sci. 2003, 8, 272–278. [Google Scholar] [CrossRef]
- Brouwer, B.; Ziolkowska, A.; Bagard, M.; Keech, O.; Gardeström, P. The impact of light intensity on shade-induced leaf senescence. Plant Cell Environ. 2012, 35, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Wollaston, V.; Page, T.; Harrison, E.; Breeze, E.; Lim, P.O.; Gil Nam, H.; Lin, J.-F.; Wu, S.-H.; Swidzinski, J.; Ishizaki, K.; et al. Comparative transcriptome analysis reveals significant differences in gene expression and signalling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J. 2005, 42, 567–585. [Google Scholar] [CrossRef]
- Lee, R.; Wang, C.; Huang, L.; Chen, S.G. Leaf senescence in rice plants: Cloning and characterization of senescence up-regulated genes. J. Exp. Bot. 2001, 52, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Worley, E.; Udvardi, M. A NAP-AAO3 regulatory module promotes chlorophyll degradation via ABA biosynthesis in Arabidopsis leaves. Plant Cell 2014, 26, 4862–4874. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Gan, S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.J.; Vu, Q.T.; Jung, S.; McClung, C.R.; Hong, S.; Gil Nam, H. Circadian control of ORE1 by PRR9 positively regulates leaf senescence in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 8448–8453. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, M.; Sakuraba, Y.; Yanagisawa, S. A Jasmonate-activated MYC2-Dof2.1-MYC2 Transcriptional Loop Promotes Leaf Senescence in Arabidopsis. Plant Cell 2019. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Tanaka, A. Rice Non-Yellow Coloring1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an α/β hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J. 2009, 59, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Han, S.H.; Yang, H.J.; Piao, W.; Paek, N.C. Mutation of Rice Early Flowering3. 1 (OsELF3. 1) delays leaf senescence in rice. Plant Mol. Biol. 2016, 92, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.-S.; Penfold, C.A.; Jenkins, D.; et al. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, S.; Sano, S.; Koshiba, T.; Ushimaru, T. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence: Comparison with those during dark-induced senescence. Physiol. Plant. 2000, 109, 211–216. [Google Scholar] [CrossRef]
- Liebsch, D.; Keech, O. DarK-induced leaf senescence: New insights into a complex light-dependent regulatory pathway. New Phytol. 2016, 212, 563–570. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Jeong, J.; Kang, M.-Y.; Kim, J.; Paek, N.-C.; Choi, G. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis. Nat. Commun. 2014, 5, 4636. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yang, C.; Gao, S.; Zhang, W.; Li, L.; Kuai, B. Age-triggered and dark-induced leaf senescence require the bHLH transcription factors PIF3, 4, and 5. Mol. Plant 2014, 7, 1776–1787. [Google Scholar] [CrossRef] [Green Version]
- Qiu, K.; Li, Z.; Yang, Z.; Chen, J.; Wu, S.; Zhu, X.; Gao, S.; Gao, J.; Ren, G.; Kuai, B.; et al. EIN3 and ORE1 Accelerate Degreening during Ethylene-Mediated Leaf Senescence by Directly Activating Chlorophyll Catabolic Genes in Arabidopsis. PLoS Genet. 2015, 11, e1005399. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.; Weltmeier, F.; Ehlert, A.; Weiste, C.; Stahl, M.; Harter, K.; Dröge-Laser, W. Heterodimers of the Arabidopsis Transcription Factors bZIP1 and bZIP53 Reprogram Amino Acid Metabolism during Low Energy Stress. Plant Cell 2011, 23, 381–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mair, A.; Pedrotti, L.; Wurzinger, B.; Anrather, R.; Simeunovic, A.; Weiste, C.; Valerio, C.; Dietrich, K.; Kirchler, T.; Nagele, T.; et al. Decision letter: SnRK1-triggered switch of bZIP63 dimerization mediates the low-energy response in plants. Decis. Lett. 2015, 4, 05828. [Google Scholar]
- Huang, X.; Shi, H.; Hu, Z.; Liu, A.; Amombo, E.; Chen, L.; Fu, J. ABA Is Involved in Regulation of Cold Stress Response in Bermudagrass. Front. Plant Sci. 2017, 8, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hieno, A.; Naznin, H.A.; Inaba-Hasegawa, K.; Yokogawa, T.; Hayami, N.; Nomoto, M.; Tada, Y.; Yokogawa, T.; Higuchi-Takeuchi, M.; Hanada, K.; et al. Transcriptome Analysis and Identification of a Transcriptional Regulatory Network in the Response to H2O2. Plant Physiol. 2019, 180, 1629–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Kim, I.; Kim, Y.-K.; Heo, J.B.; Suh, M.C.; Kim, H.U. Strigolactone Signaling Genes Showing Differential Expression Patterns in Arabidopsis max Mutants. Plants 2019, 8, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Gho, Y.-S.; Jung, K.-H.; Kim, S.-R. Genome-Wide Identification and Analysis of Genes, Conserved between japonica and indica Rice Cultivars, that Respond to Low-Temperature Stress at the Vegetative Growth Stage. Front. Plant Sci. 2017, 8, 1120. [Google Scholar] [CrossRef] [Green Version]
- Murshid, A.; Eguchi, T.; Calderwood, S.K. Stress proteins in aging and life span. Int. J. Hyperth. 2013, 29, 442–447. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Jiang, Z.; Yu, D. WRKY22 Transcription Factor Mediates Dark-Induced Leaf Senescence in Arabidopsis. Mol. Cells 2011, 31, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Li, Z.; Huang, P.; Li, B.; Fang, S.; Chu, J.; Guo, H. A Tripartite Amplification Loop Involving the Transcription Factor WRKY75, Salicylic Acid, and Reactive Oxygen Species Accelerates Leaf Senescence. Plant Cell 2017, 29, 2854–2870. [Google Scholar] [CrossRef]
- Miao, Y.; Laun, T.; Zimmermann, P.; Zentgraf, U. Targets of the WRKY53 transcription factor and its role during leaf senescence in Arabidopsis. Plant Mol. Boil. 2004, 55, 853–867. [Google Scholar] [CrossRef]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.-P.; Mueller-Roeber, B. ORS1, an H2O2—Responsive NAC Transcription Factor, Controls Senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balazadeh, S.; Siddiqui, H.; Allu, A.D.; Matallana-Ramirez, L.P.; Caldana, C.; Mehrnia, M.; Köhler, B.; Mueller-Roeber, B.; Zanor, M.-I. A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant J. 2010, 62, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Rosado, D.; Gramegna, G.; Cruz, A.; Lira, B.S.; Freschi, L.; De Setta, N.; Rossi, M. Phytochrome Interacting Factors (PIFs) in Solanum lycopersicum: Diversity, Evolutionary History and Expression Profiling during Different Developmental Processes. PLoS ONE 2016, 11, e0165929. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Yoo, S.-D. Novel connections and gaps in ethylene signaling from the ER membrane to the nucleus. Front. Plant Sci. 2015, 5, 733. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Z.; Chen, Y.; He, J.-X.; Bi, Y. Phytochrome-Interacting Factor 5 (PIF5) positively regulates dark-induced senescence and chlorophyll degradation in Arabidopsis. Plant Sci. 2015, 237, 57–68. [Google Scholar] [CrossRef]
- Li, Z.; Peng, J.; Wen, X.; Guo, H. Ethylene-Insensitive3 Is a Senescence-Associated Gene That Accelerates Age-Dependent Leaf Senescence by Directly Repressing miR164 Transcription in Arabidopsis. Plant Cell 2013, 25, 3311–3328. [Google Scholar] [CrossRef] [Green Version]
- Khanna, R.; Shen, Y.; Marion, C.M.; Tsuchisaka, A.; Theologis, A.; Schäfer, E.; Quail, P.H. The Basic Helix-Loop-Helix Transcription Factor PIF5 Acts on Ethylene Biosynthesis and Phytochrome Signaling by Distinct Mechanisms. Plant Cell 2007, 19, 3915–3929. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.-H.; Guo, F.-Q. Nitric Oxide Regulates Dark-Induced Leaf Senescence Through EIN2 in ArabidopsisF. J. Integr. Plant Boil. 2012, 54, 516–525. [Google Scholar] [CrossRef]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, I.H.; Jun, J.H.; Phee, B.-K.; Rupak, T.; Jeong, H.; Lee, Y.; Hong, B.S.; et al. Gene regulatory cascade of senescence-associated NAC transcription factors activated by Ethylene-Insensitive2-mediated leaf senescence signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.A.; Park, J.; Lee, G.I.; Paek, K.H.; Park, S.K.; Gil Nam, H.; Miklashevichs, E.; Czaja, I.; Cordeiro, A.; Prinsen, E.; et al. Identification of three genetic loci controlling leaf senescence in Arabidopsis thaliana. Plant J. 1997, 12, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Van Der Graaff, E.; Schwacke, R.; Schneider, A.; DeSimone, M.; Flügge, U.-I.; Kunze, R. Transcription Analysis of Arabidopsis Membrane Transporters and Hormone Pathways during Developmental and Induced Leaf Senescence1. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiba, T.; Naitou, T.; Koizumi, N.; Yamashino, T.; Sakakibara, H.; Mizuno, T. Combinatorial Microarray Analysis Revealing Arabidopsis Genes Implicated in Cytokinin Responses through the His→Asp Phosphorelay Circuitry. Plant Cell Physiol. 2005, 46, 339–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhang, Y.; Di, C.; Zhang, Q.; Zhang, K.; Wang, C.; You, Q.; Yan, H.; Dai, S.Y.; Yuan, J.S.; et al. JAZ7 negatively regulates dark-induced leaf senescence in Arabidopsis. J. Exp. Bot. 2015, 67, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.F.; Wu, S.H. Molecular events in senescing Arabidopsis leaves. Plant J. 2004, 39, 612–628. [Google Scholar] [CrossRef]
- Ding, N.; Nie, H.; Sun, X.; Sun, W.; Qu, Y.; Liu, X.; Yao, Y.; Liang, X.; Chen, C.C.; Li, Y. Human serum N-glycan profiles are age and sex dependent. Age Ageing 2011, 40, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Meli, V.S.; Ghosh, S.; Prabha, T.N.; Chakraborty, N.; Chakraborty, S.; Datta, A. Enhancement of fruit shelf life by suppressing N-glycan processing enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 2413–2418. [Google Scholar] [CrossRef] [Green Version]
- Almoguera, C.; Personat, J.-M.; Prieto-Dapena, P.; Jordano, J. Heat shock transcription factors involved in seed desiccation tolerance and longevity retard vegetative senescence in transgenic tobacco. Planta 2015, 242, 461–475. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Boil. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Yu, N.; Li, J.; Qi, Z.; Wang, D.; Chen, L. Heritability and Reversibility of DNA Methylation Induced by in vitro Grafting between Brassica juncea and B. oleracea. Sci. Rep. 2016, 6, 27233. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Bases | Error (%) | Q20(%) 1 | Q30(%) 2 | GC(%) 3 |
|---|---|---|---|---|---|---|---|
| CK_1 | 45,224,598 | 42,557,344 | 6.38 | 0.03 | 97.90 | 94.07 | 51.53 |
| CK_2 | 55,052,156 | 53,558,496 | 8.03 | 0.02 | 98.21 | 94.71 | 53.28 |
| CK_3 | 59,896,392 | 58,132,180 | 8.72 | 0.02 | 98.35 | 95.04 | 53.44 |
| DT_1 | 67,686,056 | 64,587,208 | 9.69 | 0.02 | 98.03 | 94.40 | 52.53 |
| DT_2 | 54,946,418 | 52,195,444 | 7.83 | 0.02 | 98.10 | 94.51 | 52.67 |
| DT_3 | 65,845,158 | 60,166,630 | 9.02 | 0.02 | 98.00 | 94.31 | 52.34 |
| Category | Transcripts | Unigenes |
|---|---|---|
| 200–500 bp | 128,446 | 60,507 |
| 500–1 kbp | 111,291 | 53,622 |
| 1–2 kbp | 97,001 | 30,181 |
| >2 kbp | 50,483 | 14,897 |
| Min length (bp) 1 | 301 | 301 |
| Mean length (bp) | 1090 | 948 |
| Max length (bp) 2 | 53,494 | 53,494 |
| N50 length (bp) 3 | 1559 | 1284 |
| N90 length (bp) 4 | 465 | 426 |
| Total | 387,221 | 159,207 |
| Categories | Number of Unigenes | Percentage (%) |
|---|---|---|
| Annotated in NR | 39,336 | 24.7 |
| Annotated in NT | 76,881 | 48.28 |
| Annotated in KEGG | 36,014 | 22.62 |
| Annotated in Swiss-Prot | 83,705 | 52.57 |
| Annotated in PFAM | 84,295 | 52.94 |
| Annotated in GO | 84,295 | 52.94 |
| Annotated in KOG | 41,079 | 25.8 |
| Annotated in all Databases | 6961 | 4.37 |
| Annotated in at least one Database | 119,581 | 75.11 |
| Total Unigenes | 159,207 | 100 |
| GeneID | Log2(DT/CK) | padj-value | Gene Description |
|---|---|---|---|
| Cluster-109720.12769-2R | −2.8599 | 1.71 × 10−16 | Transcription factor PIF4 OS = Arabidopsis thaliana GN = PIF4 PE = 1 SV = 1 |
| Cluster-107104.0-1F | −7.2349 | 1.02 × 10−19 | Transcription factor PIF4 OS = Arabidopsis thaliana GN = PIF1 PE = 1 SV = 1 |
| Cluster-113186.0-2F | −5.49 | 1.41 × 10−10 | Transcription factor PIF3 OS = Oryza sativa subsp. japonica GN = PIF3 PE = 1 SV = 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Lou, Y.; Shi, H.; Chen, L.; Cao, L. Transcriptomic Analysis of Dark-Induced Senescence in Bermudagrass (Cynodon dactylon). Plants 2019, 8, 614. https://doi.org/10.3390/plants8120614
Fan J, Lou Y, Shi H, Chen L, Cao L. Transcriptomic Analysis of Dark-Induced Senescence in Bermudagrass (Cynodon dactylon). Plants. 2019; 8(12):614. https://doi.org/10.3390/plants8120614
Chicago/Turabian StyleFan, Jibiao, Yanhong Lou, Haiyan Shi, Liang Chen, and Liwen Cao. 2019. "Transcriptomic Analysis of Dark-Induced Senescence in Bermudagrass (Cynodon dactylon)" Plants 8, no. 12: 614. https://doi.org/10.3390/plants8120614