Research Progress on the Functions of Gasotransmitters in Plant Responses to Abiotic Stresses



Abstract
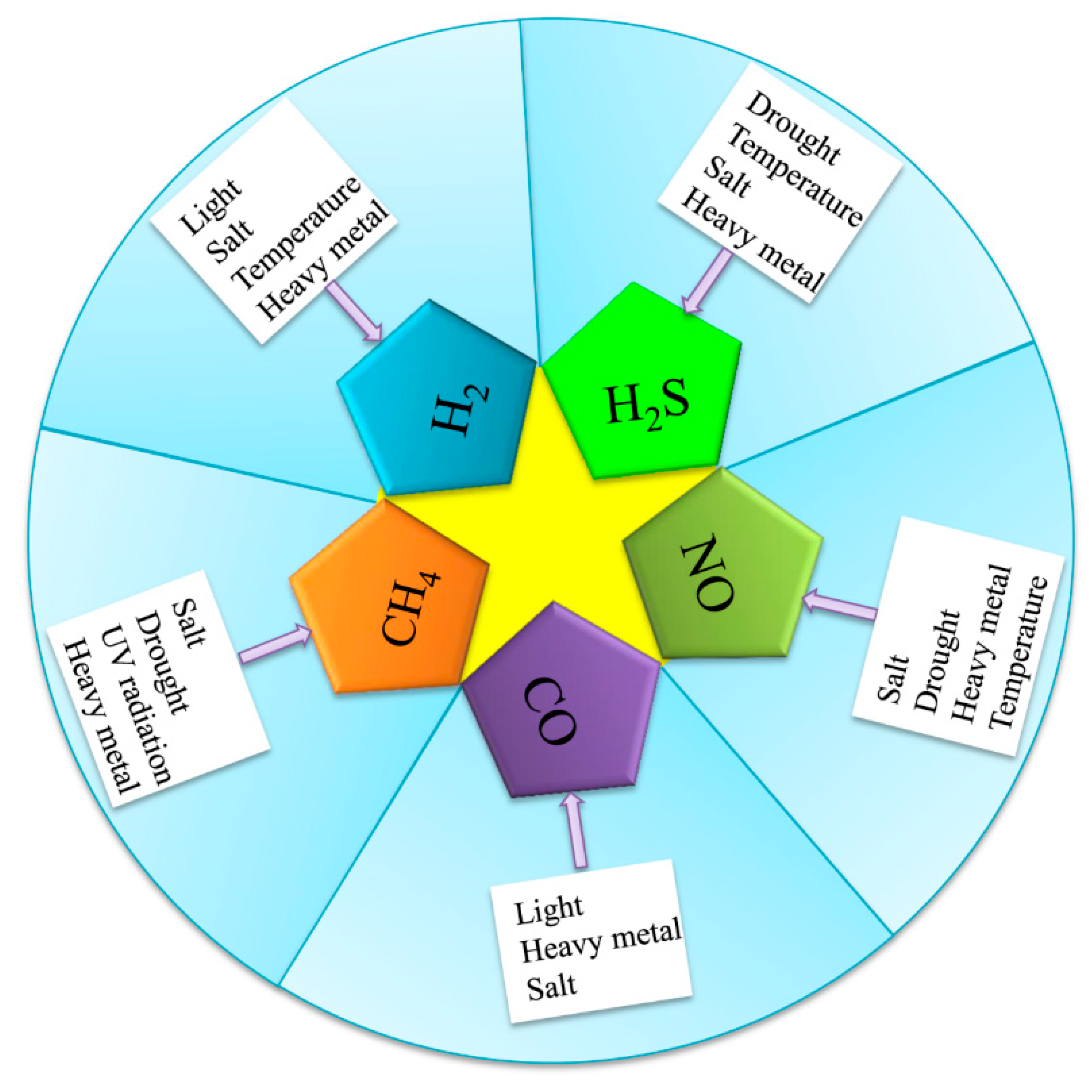
:1. Introduction
2. Production of Gasotransmitters under Adverse Conditions
2.1. Hydrogen Gas (H2)
2.2. Hydrogen Sulfide (H2S)
2.3. Nitric Oxide (NO)
2.4. Carbon Monoxide (CO)
2.5. Methane (CH4)
3. The Role of Gasotransmitters under Adverse Conditions
3.1. Hydrogen Gas (H2)
3.1.1. Heavy Metal Stress
3.1.2. Salt and Temperature Stresses
3.1.3. Ultraviolet Radiation
3.1.4. Drought and Paraquat Stresses
3.2. Hydrogen Sulfide (H2S)
3.2.1. Heavy Metal Stress
3.2.2. Salt Stress
3.2.3. Temperature Stress
3.2.4. Drought Stress
3.3. Nitric Oxide (NO)
3.3.1. Heavy Metal Stress
3.3.2. Salt Stress
3.3.3. Temperature and Drought Stress
3.4. Carbon Monoxide (CO)
3.4.1. Heavy Metal Stress
3.4.2. Salt Stress
3.4.3. Drought and Temperature Stress
3.5. Methane (CH4)
4. Gasotransmitter Interactions under Adverse Conditions
4.1. Interaction between H2 and NO
4.2. Interaction between H2 and CO
4.3. Crosstalk between H2S and NO
4.4. Crosstalk between NO and CO
4.5. Interaction between CH4 and Other Signallings
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| H2 | hydrogen gas |
| H2S | hydrogen Sulfide |
| NO | nitric Oxide |
| CO | carbon Monoxide |
| CH4 | methane |
| EC | electrolyte leakage |
| H2O2 | hydrogen peroxide |
| MDA | malondialdehyde |
| ROS | reactive oxygen species |
| ABA | abscisic acid |
| GSH | glutathione |
| AsA-GSH | ascorbate–glutathione |
| AsA | ascorbic acid |
| HO-1 | heme oxygenase-1 |
| HO | heme oxygenase |
| HRW | hydrogen-rich water |
| Cu | copper |
| Cd | cadmium |
| Al | aluminum |
| Hg | mercury |
| Zn | zinc |
| Ni | nickel |
| Pb | lead |
| As | arsenate |
| UV | ultraviolet radiation |
| NPT | nonprotein thiols |
| Pro | proline |
| pH | potential of hydrogen |
| SOD | superoxide dismutase |
| MAPK | mitogen-activated protein kinase |
| TPC | total phenolic contents |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| TBARS | thiobarbituric acid reactive substances |
| CAT | catalase |
| APX | ascorbate peroxidase |
| POD | peroxidase |
| LCD | L-cysteine desulfhydrase |
| DCD | D-cysteine desulfhydrase |
| GR | glutathione reductase |
| DHAR | dehydroascorbate reductase |
| PCD | programmed cell death |
| O2⋅− | superoxide anion |
| ·OH | hydroxy radicals |
References
- Ali, Q.; Daud, M.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L.) by enhancing physiological and biochemical parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Wang, Z.; Ma, Q.; Sun, L.; Zhang, L.; Liu, Z.; Liu, D.; Hao, X.; Pei, Y. Hydrogen sulfide mediates ion fluxes inducing stomatal closure in response to drought stress in Arabidopsis thaliana. Plant Soil 2017, 419, 141–152. [Google Scholar] [CrossRef]
- Shen, Q.; Jiang, M.; Li, H.; Che, L.L.; Yang, Z.M. Expression of a Brassica napus heme oxygenase confers plant tolerance to mercury toxicity. Plant Cell Environ. 2011, 34, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, M.; Jiang, M. Hydrogen sulfide alleviates mercury toxicity by sequestering it in roots or regulating reactive oxygen species productions in rice seedlings. Plant Physiol. Biochem. 2017, 111, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, L.; Lu, L.; Jin, C.; Lin, X. Nitric oxide acts downstream of hydrogen peroxide in regulating aluminum-induced antioxidant defense that enhances aluminum resistance in wheat seedlings. Environ. Exp. Bot. 2018, 145, 95–103. [Google Scholar] [CrossRef]
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric oxide alleviates salt stress inhibited photosynthetic performance by interacting with sulfur assimilation in mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Su, N.; Cai, J.; Shen, Z.; Cui, J. Hydrogen-rich water enhances cadmium tolerance in Chinese cabbage by reducing cadmium uptake and increasing antioxidant capacities. J. Plant Physiol. 2015, 175, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Alleman, R.J.; Katunga, L.A.; Nelson, M.A.; Brown, D.A.; Anderson, E.J. The “Goldilocks Zone” from a redox perspective—Adaptive vs. deleterious responses to oxidative stress in striated muscle. Front. Physiol. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.; Cui, W.; Dai, C.; Zhu, K.; Zhang, J.; Wang, R.; La, H.; Li, X.; Shen, W. Involvement of hydrogen peroxide and heme oxygenase-1 in hydrogen gas-induced osmotic stress tolerance in alfalfa. Plant Growth Regul. 2016, 80, 215–223. [Google Scholar] [CrossRef]
- Wang, R. Overview of Gasotransmitters and the Related Signaling Network. In Gasotransmitters; Wang, R., Ed.; Royal Society of Chemistry: Cambridge, UK, 2018; pp. 1–28. [Google Scholar]
- Abdulmajeed, A.M.; Derby, S.R.; Strickland, S.K.; Qaderi, M.M. Interactive effects of temperature and UVB radiation on methane emissions from different organs of pea plants grown in hydroponic system. J. Photochem. Photobiol. B Biol. 2017, 166, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Cao, H.; Yao, P.; Pan, J.; Gu, Q.; Xu, S.; Wang, R.; Ouyang, Z.; Wang, Q.; Shen, W. Methane enhances aluminum resistance in alfalfa seedlings by reducing aluminum accumulation and reestablishing redox homeostasis. Biometals 2017, 30, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, R.; Yang, W.; Chen, Z.; Hu, X. Carbon monoxide signal regulates light-initiated seed germination by suppressing SOM expression. Plant Sci. 2018, 272, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Maryan, K.E.; Lahiji, H.S.; Farrokhi, N.; Komeleh, H.H. Analysis of Brassica napus dehydrins and their Co-Expression regulatory networks in relation to cold stress. Gene Expr. Patterns 2019, 31, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Cao, H.; Fang, W.; Pan, J.; Chen, J.; Zhang, J.; Shen, W. Linking hydrogen-enhanced rice aluminum tolerance with the reestablishment of GA/ABA balance and miRNA-modulated gene expression: A case study on germination. Ecotoxicol. Environ. Saf. 2017, 145, 303. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, M.; Stickland, L.H. Hydrogenase: A bacterial enzyme activating molecular hydrogen: The properties of the enzyme. Biochem. J. 1931, 25, 205. [Google Scholar]
- Sanadze, G. Absorption of molecular hydrogen by green leaves in light. Fiziol Rast 1961, 8, 555–559. [Google Scholar]
- Renwick, G.; Giumarro, C.; Siegel, S. Hydrogen metabolism in higher plants. Plant Physiol. 1964, 39, 303. [Google Scholar] [CrossRef]
- Xu, S.; Zhu, S.; Jiang, Y.; Wang, N.; Wang, R.; Shen, W.; Yang, J. Hydrogen-rich water alleviates salt stress in rice during seed germination. Plant Soil 2013, 370, 47–57. [Google Scholar] [CrossRef]
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Shen, W. H2 Enhances Arabidopsis Salt Tolerance by Manipulating ZAT10/12-Mediated Antioxidant Defence and Controlling Sodium Exclusion. PLoS ONE 2012, 7, e49800. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Jiang, Y.; Cui, W.; Jin, Q.; Zhang, Y.; Bu, D.; Fu, J.; Wang, R.; Zhou, F.; Shen, W. Hydrogen enhances adaptation of rice seedlings to cold stress via the reestablishment of redox homeostasis mediated by miRNA expression. Plant Soil 2017, 414, 53–67. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zhang, M.; Sun, X. Molecular Hydrogen Is Involved in Phytohormone Signaling and Stress Responses in Plants. PLoS ONE 2013, 8, e71038. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.T.; Whiteman, M. Hydrogen sulfide and cell signaling: Team player or referee? Plant Physiol. Biochem. 2014, 78, 37–42. [Google Scholar] [CrossRef]
- Hancock, J.T. Hydrogen sulfide and environmental stresses. Environ. Exp. Bot. 2019, 161, 50–56. [Google Scholar] [CrossRef]
- Jin, Z.; Shen, J.; Qiao, Z.; Yang, G.; Wang, R.; Pei, Y. Hydrogen sulfide improves drought resistance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2011, 414, 481–486. [Google Scholar] [CrossRef]
- Ma, D.; Ding, H.; Wang, C.; Qin, H.; Han, Q.; Hou, J.; Lu, H.; Xie, Y.; Guo, T. Alleviation of drought stress by hydrogen sulfide is partially related to the abscisic acid signaling pathway in wheat. PLoS ONE 2016, 11, e0163082. [Google Scholar] [CrossRef]
- Fu, P.; Wang, W.; Hou, L.; Liu, X. Hydrogen sulfide is involved in the chilling stress response in Vitis vinifera L. Acta Soc. Bot. Pol. 2013, 82, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, Y.; Cao, C.; Liang, S.; Ma, Y.; Liu, X.; Pei, Y. The role of H2S in low temperature-induced cucurbitacin C increases in cucumber. Plant Mol. Biol. 2019, 99, 535–544. [Google Scholar] [CrossRef]
- Cheng, T.; Shi, J.; Dong, Y.; Ma, Y.; Peng, Y.; Hu, X.; Chen, J. Hydrogen sulfide enhances poplar tolerance to high-temperature stress by increasing S-nitrosoglutathione reductase (GSNOR) activity and reducing reactive oxygen/nitrogen damage. Plant Growth Regul. 2018, 84, 11–23. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Chan, Z. Nitric oxide-activated hydrogen sulfide is essential for cadmium stress response in bermudagrass (Cynodon dactylon (L). Pers.). Plant Physiol. Biochem. 2014, 74, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yang, B.; Hao, Z.; Zhu, J.; Zhang, Y.; Xu, T. Exogenous hydrogen sulfide ameliorates seed germination and seedling growth of cauliflower under lead stress and its antioxidant role. J. Plant Growth Regul. 2018, 37, 5–15. [Google Scholar] [CrossRef]
- Valivand, M.; Amooaghaie, R.; Ahadi, A. Interplay between hydrogen sulfide and calcium/calmodulin enhances systemic acquired acclimation and antioxidative defense against nickel toxicity in zucchini. Environ. Exp. Bot. 2019, 158, 40–50. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Siddiqui, M.H. Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide 2017, 68, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B.; González-Gordo, S.; Muñoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghdam, M.S.; Mahmoudi, R.; Razavi, F.; Rabiei, V.; Soleimani, A. Hydrogen sulfide treatment confers chilling tolerance in hawthorn fruit during cold storage by triggering endogenous H2S accumulation, enhancing antioxidant enzymes activity and promoting phenols accumulation. Sci. Hortic. 2018, 238, 264–271. [Google Scholar] [CrossRef]
- Jost, R.; Berkowitz, O.; Wirtz, M.; Hopkins, L.; Hawkesford, M.; Hell, R. Genomic and functional characterization of the oas gene family encoding O-acetylserine (thiol) lyases, enzymes catalyzing the final step in cysteine biosynthesis in Arabidopsis thaliana. Gene 2000, 253, 237–247. [Google Scholar] [CrossRef]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2017, 69, 3401–3411. [Google Scholar] [CrossRef]
- Kolbert, Z.; Feigl, G.; Freschi, L.; Poór, P. Gasotransmitters in action: Nitric oxide-ethylene crosstalk during plant growth and abiotic stress responses. Antioxidants 2019, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Klepper, L. Nitric oxide (NO) and nitrogen dioxide (NO2) emissions from herbicide-treated soybean plants. Atmos. Environ. (1967) 1979, 13, 537–542. [Google Scholar] [CrossRef]
- Da Silva, C.J.; Fontes, E.P.B.; Modolo, L.V. Salinity-induced accumulation of endogenous H2S and NO is associated with modulation of the antioxidant and redox defense systems in Nicotiana tabacum L. cv. Havana. Plant Sci. 2017, 256, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, R.-J.; Han, T.-T.; Cai, W.; Fu, Z.-W.; Lu, Y.-T. Salt stress reduces root meristem size by nitric oxide-mediated modulation of auxin accumulation and signaling in Arabidopsis. Plant Physiol. 2015, 168, 343–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous nitric oxide donor and arginine provide protection against short-term drought stress in wheat seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.-B.; Huang, G.-B.; Yu, J.-H.; Zhang, M.-L. Nitric oxide and hydrogen peroxide alleviate drought stress in marigold explants and promote its adventitious root development. Plant Physiol. Biochem. 2012, 58, 6–15. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, J.; Cheng, D.; Wang, R.; Mei, Y.; Hu, H.; Shen, W.; Zhang, Y. Nitric oxide contributes to methane-induced osmotic stress tolerance in mung bean. BMC Plant Biol. 2018, 18, 207. [Google Scholar] [CrossRef]
- Faria-Lopes, R.O.; Muniz, D.R.; Chaves, I.S.; França, M.G.; Modolo, L.V. Nitric oxide precursors prevent Al-triggered auxin flow inhibition in Triticum aestivum roots. J. Adv. Res. 2019, 15, 27–36. [Google Scholar] [CrossRef]
- He, H.; Huang, W.; Oo, T.L.; Gu, M.; Zhan, J.; Wang, A.; He, L.-F. Nitric oxide suppresses aluminum-induced programmed cell death in peanut (Arachis hypoganea L.) root tips by improving mitochondrial physiological properties. Nitric Oxide 2018, 74, 47–55. [Google Scholar] [CrossRef]
- Han, B.; Yang, Z.; Xie, Y.; Nie, L.; Cui, J.; Shen, W. Arabidopsis HY1 confers cadmium tolerance by decreasing nitric oxide production and improving iron homeostasis. Mol. Plant 2014, 7, 388–403. [Google Scholar] [CrossRef] [Green Version]
- Kováčik, J.; Dresler, S.; Micalizzi, G.; Babula, P.; Hladký, J.; Mondello, L. Nitric oxide affects cadmium-induced changes in the lichen Ramalina farinacea. Nitric Oxide 2019, 83, 11–18. [Google Scholar] [CrossRef]
- Liao, W.; Huang, G.; Yu, J.; Zhang, M.; Shi, X. Nitric oxide and hydrogen peroxide are involved in indole-3-butyric acid-induced adventitious root development in marigold. J. Hortic. Sci. Biotechnol. 2011, 86, 159–165. [Google Scholar] [CrossRef]
- Liao, W.-B.; Zhang, M.-L.; Yu, J.-H. Role of nitric oxide in delaying senescence of cut rose flowers and its interaction with ethylene. Sci. Hortic. 2013, 155, 30–38. [Google Scholar] [CrossRef]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Morris, P.; Ribeiro, D.; Wilson, I. Nitric oxide, stomatal closure, and abiotic stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liao, W.; Wang, M.; Niu, L.; Xu, Q.; Jin, X. Nitric oxide is required for hydrogen gas-induced adventitious root formation in cucumber. J. Plant Physiol. 2016, 195, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Wilks, S.S. Carbon monoxide in green plants. Science 1959, 129, 964–966. [Google Scholar] [CrossRef]
- Tarr, M.A.; Miller, W.L.; Zepp, R.G. Direct carbon monoxide photoproduction from plant matter. J. Geophys. Res. Atmos. 1995, 100, 11403–11413. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, J.; Chen, X.; Gao, Z.; Xuan, W.; Xu, S.; Ding, X.; Shen, W. Carbon monoxide alleviates cadmium-induced oxidative damage by modulating glutathione metabolism in the roots of Medicago sativa. New Phytol. 2008, 177, 155–166. [Google Scholar] [CrossRef]
- Zilli, C.G.; Santa-Cruz, D.M.; Balestrasse, K.B. Heme oxygenase-independent endogenous production of Carbon Monoxide by soybean plants subjected to salt stress. Environ. Exp. Bot. 2014, 102, 11–16. [Google Scholar] [CrossRef]
- Nouchi, I.; Mariko, S.; Aoki, K. Mechanism of methane transport from the rhizosphere to the atmosphere through rice plants. Plant Physiol. 1990, 94, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Keppler, F.; Hamilton, J.T.; Braß, M.; Röckmann, T. Methane emissions from terrestrial plants under aerobic conditions. Nature 2006, 439, 187. [Google Scholar] [CrossRef]
- McLeod, A.R.; Fry, S.C.; Loake, G.J.; Messenger, D.J.; Reay, D.S.; Smith, K.A.; Yun, B.W. Ultraviolet radiation drives methane emissions from terrestrial plant pectins. New Phytol. 2008, 180, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, D.; Mikkelsen, T.N.; Øbro, J.; Willats, W.G.T.; Ambus, P. Effects of temperature, ultraviolet radiation and pectin methyl esterase on aerobic methane release from plant material. Plant Biol. 2009, 11, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, D.; Mikkelsen, T.N.; Rolsted, M.M.M.; Egsgaard, H.; Ambus, P. Leaf surface wax is a source of plant methane formation under UV radiation and in the presence of oxygen. Plant Biol. 2014, 16, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Brüggemann, N.; Meier, R.; Steigner, D.; Zimmer, I.; Louis, S.; Schnitzler, J.P. Nonmicrobial aerobic methane emission from poplar shoot cultures under low-light conditions. New Phytol. 2009, 182, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Cui, W.; Dai, C.; Wu, M.; Zhang, J.; Zhang, Y.; Xie, Y.; Shen, W. Methane-rich water alleviates NaCl toxicity during alfalfa seed germination. Environ. Exp. Bot. 2016, 129, 37–47. [Google Scholar] [CrossRef]
- Han, B.; Duan, X.; Wang, Y.; Zhu, K.; Zhang, J.; Wang, R.; Hu, H.; Qi, F.; Pan, J.; Yan, Y. Methane protects against polyethylene glycol-induced osmotic stress in maize by improving sugar and ascorbic acid metabolism. Sci. Rep. 2017, 7, 46185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samma, M.K.; Zhou, H.; Cui, W.; Zhu, K.; Zhang, J.; Shen, W. Methane alleviates copper-induced seed germination inhibition and oxidative stress in Medicago sativa. Biometals 2017, 30, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Z.; Cui, W.; Zhang, Y.; Hu, H.; Yu, X.; Wang, Q.; Shen, W. Methane alleviates alfalfa cadmium toxicity via decreasing cadmium accumulation and reestablishing glutathione homeostasis. Ecotoxicol. Environ. Saf. 2018, 147, 861–871. [Google Scholar] [CrossRef]
- Martel, A.B.; Qaderi, M.M. Unravelling the effects of blue light on aerobic methane emissions from canola. J. Plant Physiol. 2019, 233, 12–19. [Google Scholar] [CrossRef]
- Messenger, D.J.; McLeod, A.R.; Fry, S.C. The role of ultraviolet radiation, photosensitizers, reactive oxygen species and ester groups in mechanisms of methane formation from pectin. Plant Cell Environ. 2009, 32, 1–9. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Wu, Q.; Su, N.; Chen, Q.; Shen, W.; Shen, Z.; Xia, Y.; Cui, J. Cadmium-induced hydrogen accumulation is involved in cadmium tolerance in Brassica campestris by reestablishment of reduced glutathione homeostasis. PLoS ONE 2015, 10, e0139956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Cui, W.; Pan, J.; Xie, Y.; Wang, J.; Shen, W. Proteomic analysis provides insights into the molecular bases of hydrogen gas-induced cadmium resistance in Medicago sativa. J. Proteom. 2017, 152, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, Q.; Wang, Y.; Shen, Z.; Shen, W.; Xu, X. Hydrogen-rich water induces aluminum tolerance in maize seedlings by enhancing antioxidant capacities and nutrient homeostasis. Ecotoxicol. Environ. Saf. 2017, 144, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cui, W.; Zhu, K.; Xie, Y.; Zhang, C.; Shen, W. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Fang, P.; Zhu, K.; Mao, Y.; Gao, C.; Xie, Y.; Wang, J.; Shen, W. Hydrogen-rich water confers plant tolerance to mercury toxicity in alfalfa seedlings. Ecotoxicol. Environ. Saf. 2014, 105, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.T.; Hancock, T.H. Hydrogen Gas, ROS Metabolism, and Cell Signaling: Are Hydrogen Spin States Important? React. Oxyg. Species 2018, 6, 389–395. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, X.; Lei, D.; Hu, S.; Shen, Z.; Shen, W.; Xu, X. Hydrogen-rich water pretreatment alters photosynthetic gas exchange, chlorophyll fluorescence, and antioxidant activities in heat-stressed cucumber leaves. Plant Growth Regul. 2017, 83, 1–14. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Liu, Y.; Cai, J.; Shen, W.; Xia, K.; Cui, J. Hydrogen-rich water reestablishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation. J. Agric. Food Chem. 2014, 62, 6454–6462. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, Z.; Duan, X.; Chen, D.; Zhang, Y.; Cui, W.; Ren, W.; Shen, W. Hydrogen-rich water-alleviated ultraviolet-B-triggered oxidative damage is partially associated with the manipulation of the metabolism of (iso)flavonoids and antioxidant defence in Medicago sativa. Funct. Plant Biol. 2015, 42, 1–10. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, J.; Tian, J.; Li, N.; Jia, L.; Shen, W.; Cui, J. Enhanced anthocyanin accumulation of immature radish microgreens by hydrogen-rich water under short wavelength light. Sci. Hortic. 2019, 247, 75–85. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Li, L.; Shen, W. Hydrogen-modulated stomatal sensitivity to abscisic acid and drought tolerance via the regulation of apoplastic pH in Medicago sativa. J. Plant Growth Regul. 2016, 35, 565–573. [Google Scholar] [CrossRef]
- Singh, V.P.; Singh, S.; Kumar, J.; Prasad, S.M. Hydrogen sulfide alleviates toxic effects of arsenate in pea seedlings through up-regulation of the ascorbate–glutathione cycle: Possible involvement of nitric oxide. J. Plant Physiol. 2015, 181, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Qiao, Z.; Zhang, L.; Li, H.; Pei, Y. Hydrogen sulfide and proline cooperate to alleviate cadmium stress in foxtail millet seedlings. Plant Physiol. Biochem. 2016, 109, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Rahman, A.; Ansary, M.M.U.; Watanabe, A.; Fujita, M.; Tran, L.-S.P. Hydrogen sulfide modulates cadmium-induced physiological and biochemical responses to alleviate cadmium toxicity in rice. Sci. Rep. 2015, 5, 14078. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.-Q.; Bao, J.; Yuan, F.; Liang, X.; Feng, Z.-T.; Wang, B.-S. Exogenous hydrogen sulfide alleviates salt stress in wheat seedlings by decreasing Na+ content. Plant Growth Regul. 2016, 79, 391–399. [Google Scholar] [CrossRef]
- Jiang, J.-L.; Tian, Y.; Li, L.; Yu, M.; Hou, R.-P.; Ren, X.-M. H2S Alleviates Salinity Stress in Cucumber by Maintaining the Na+/K+ Balance and Regulating H2S Metabolism and Oxidative Stress Response. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, W.; Niu, J.; Ren, Y.; Zhang, F. Hydrogen sulfide may function downstream of hydrogen peroxide in salt stress-induced stomatal closure in Vicia faba. Funct. Plant Biol. 2019, 46, 136–145. [Google Scholar] [CrossRef]
- Qi, Q.; Guo, Z.; Liang, Y.; Li, K.; Xu, H. Hydrogen sulfide alleviates oxidative damage under excess nitrate stress through MAPK/NO signaling in cucumber. Plant Physiol. Biochem. 2019, 135, 1–8. [Google Scholar] [CrossRef]
- Luo, Z.; Li, D.; Du, R.; Mou, W. Hydrogen sulfide alleviates chilling injury of banana fruit by enhanced antioxidant system and proline content. Sci. Hortic. 2015, 183, 144–151. [Google Scholar] [CrossRef]
- Li, D.; Limwachiranon, J.; Li, L.; Du, R.; Luo, Z. Involvement of energy metabolism to chilling tolerance induced by hydrogen sulfide in cold-stored banana fruit. Food Chem. 2016, 208, 272–278. [Google Scholar] [CrossRef]
- Li, Z.-G.; Gong, M.; Xie, H.; Yang, L.; Li, J. Hydrogen sulfide donor sodium hydrosulfide-induced heat tolerance in tobacco (Nicotiana tabacum L.) suspension cultured cells and involvement of Ca2+ and calmodulin. Plant Sci. 2012, 185, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jiao, H.; Jiang, C.X.; Wang, S.H.; Wei, Z.J.; Luo, J.P.; Jones, R.L. Hydrogen sulfide protects soybean seedlings against drought-induced oxidative stress. Acta Physiol. Plant. 2010, 32, 849–857. [Google Scholar] [CrossRef]
- Jin, Z.; Xue, S.; Luo, Y.; Tian, B.; Fang, H.; Li, H.; Pei, Y. Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis. Plant Physiol. Biochem. 2013, 62, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Del-Val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xiao, Q.; Wang, C.; Wang, W.-H.; Wu, F.-H.; He, B.-Y.; Zhu, Z.; Ru, Q.-M.; Zhang, L.-L.; Zheng, H.-L. Nitric oxide alleviates oxidative stress caused by salt in leaves of a mangrove species, Aegiceras corniculatum. Aquat. Bot. 2014, 117, 41–47. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.-S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Srivastava, S.; Seth, C.S. 24-Epibrassinolide and sodium nitroprusside alleviate the salinity stress in Brassica juncea L. cv. Varuna through cross talk among proline, nitrogen metabolism and abscisic acid. Plant Soil 2017, 411, 483–498. [Google Scholar] [CrossRef]
- Yadu, S.; Dewangan, T.L.; Chandrakar, V.; Keshavkant, S. Imperative roles of salicylic acid and nitric oxide in improving salinity tolerance in Pisum sativum L. Physiol. Mol. Biol. Plants 2017, 23, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Gadelha, C.G.; de Souza Miranda, R.; Alencar, N.L.M.; Costa, J.H.; Prisco, J.T.; Gomes-Filho, E. Exogenous nitric oxide improves salt tolerance during establishment of Jatropha curcas seedlings by ameliorating oxidative damage and toxic ion accumulation. J. Plant Physiol. 2017, 212, 69–79. [Google Scholar] [CrossRef]
- Kumar, R.; Tasleem, M.; Jain, M.; Ahuja, S.; Goswami, S.; Bakshi, S.; Jambhulkar, S.; Singh, S.; Singh, G.; Pathak, H. Nitric oxide triggered defense network in wheat: Augmenting tolerance and grain-quality related traits under heat-induced oxidative damage. Environ. Exp. Bot. 2019, 158, 189–204. [Google Scholar] [CrossRef]
- Rabiei, V.; Kakavand, F.; Zaare-Nahandi, F.; Razavi, F.; Aghdam, M.S. Nitric oxide and γ-aminobutyric acid treatments delay senescence of cornelian cherry fruits during postharvest cold storage by enhancing antioxidant system activity. Sci. Hortic. 2019, 243, 268–273. [Google Scholar] [CrossRef]
- Li, Z.; Yong, B.; Cheng, B.; Wu, X.; Zhang, Y.; Zhang, X.; Peng, Y. Nitric oxide, γ-aminobutyric acid, and mannose pretreatment influence metabolic profiles in white clover under water stress. J. Integr. Plant Biol. 2019. [Google Scholar] [CrossRef]
- Philippe, F.; Verdu, I.; Morère-Le Paven, M.-C.; Limami, A.M.; Planchet, E. Involvement of Medicago truncatula glutamate receptor-like channels in nitric oxide production under short-term water deficit stress. J. Plant Physiol. 2019, 236, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.S.; Barroso, J.B.; Brouquisse, R.; Corpas, F.J.; Gupta, K.J.; Lindermayr, C.; Loake, G.J.; Palma, J.M.; Petřivalský, M.; Wendehenne, D.; et al. A forty year journey: The generation and roles of NO in plants. Nitric Oxide 2019, 93, 53–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.; Fu, G.; Wu, H.; Shen, W. Cadmium-induced heme oxygenase-1 gene expression is associated with the depletion of glutathione in the roots of Medicago sativa. Biometals 2011, 24, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.K.; Chen, J.; Yang, Z.M. Enhancement of tolerance of Indian mustard (Brassica juncea) to mercury by carbon monoxide. J. Hazard. Mater. 2011, 186, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gong, C.; Ju, X.; Zhu, Z.; Shen, W.; Shen, Z.; Cui, J. Hemin Through the Heme Oxygenase 1/Ferrous Iron, Carbon Monoxide System Involved in Zinc Tolerance in Oryza Sativa L. J. Plant Growth Regul. 2018, 37, 947–957. [Google Scholar] [CrossRef]
- Huang, B.K.; Xu, S.; Xuan, W.; Li, M.; Cao, Z.Y.; Liu, K.L.; Ling, T.F.; Shen, W.B. Carbon monoxide alleviates salt-induced oxidative damage in wheat seedling leaves. J. Integr. Plant Biol. 2006, 48, 249–254. [Google Scholar] [CrossRef]
- Xu, S.; Sa, Z.S.; Cao, Z.Y.; Xuan, W.; Huang, B.K.; Ling, T.F.; Hu, Q.Y.; Shen, W.B. Carbon monoxide alleviates wheat seed germination inhibition and counteracts lipid peroxidation mediated by salinity. J. Integr. Plant Biol. 2006, 48, 1168–1176. [Google Scholar] [CrossRef]
- Liu, K.; Xu, S.; Xuan, W.; Ling, T.; Cao, Z.; Huang, B.; Sun, Y.; Fang, L.; Liu, Z.; Zhao, N. Carbon monoxide counteracts the inhibition of seed germination and alleviates oxidative damage caused by salt stress in Oryza sativa. Plant Sci. 2007, 172, 544–555. [Google Scholar] [CrossRef]
- Ling, T.; Zhang, B.; Cui, W.; Wu, M.; Lin, J.; Zhou, W.; Huang, J.; Shen, W. Carbon monoxide mitigates salt-induced inhibition of root growth and suppresses programmed cell death in wheat primary roots by inhibiting superoxide anion overproduction. Plant Sci. 2009, 177, 331–340. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Yuan, F.; Hu, S.; He, P. Effects of hematin and carbon monoxide on the salinity stress responses of Cassia obtusifolia L. seeds and seedlings. Plant Soil 2012, 359, 85–105. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, S.; Ling, T.; Xu, L.; Shen, W. Heme oxygenase/carbon monoxide system participates in regulating wheat seed germination under osmotic stress involving the nitric oxide pathway. J. Plant Physiol. 2010, 167, 1371–1379. [Google Scholar] [CrossRef]
- Wu, H.; Zheng, Y.; Liu, J.; Zhang, H.; Chen, H. Heme oxygenase-1 delays gibberellin-induced programmed cell death of rice aleurone layers subjected to drought stress by interacting with nitric oxide. Front. Plant Sci. 2016, 6, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Zhang, Y.; Nie, Y.; Cheng, D.; Wang, R.; Hu, H.; Chen, J.; Zhang, J.; Du, Y.; Shen, W. Hydrogen-induced osmotic tolerance is associated with nitric oxide-mediated proline accumulation and reestablishment of redox balance in alfalfa seedlings. Environ. Exp. Bot. 2018, 147, 249–260. [Google Scholar] [CrossRef]
- Zhu, Y.; Liao, W.; Niu, L.; Wang, M.; Ma, Z. Nitric oxide is involved in hydrogen gas-induced cell cycle activation during adventitious root formation in cucumber. BMC Plant Biol. 2016, 16, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corpas, F.J. Nitric Oxide and Hydrogen Sulfide in Higher Plants under Physiological and Stress Conditions; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2019. [Google Scholar]
- Xie, Y.; Ling, T.; Han, Y.; Liu, K.; Zheng, Q.; Huang, L.; Yuan, X.; He, Z.; Hu, B.; Fang, L. Carbon monoxide enhances salt tolerance by nitric oxide-mediated maintenance of ion homeostasis and up-regulation of antioxidant defence in wheat seedling roots. Plant Cell Environ. 2008, 31, 1864–1881. [Google Scholar] [CrossRef] [PubMed]
- Santa-Cruz, D.M.; Pacienza, N.A.; Polizio, A.H.; Balestrasse, K.B.; Tomaro, M.L.; Yannarelli, G.G. Nitric oxide synthase-like dependent NO production enhances heme oxygenase up-regulation in ultraviolet-B-irradiated soybean plants. Phytochemistry 2010, 71, 1700–1707. [Google Scholar] [CrossRef]
- Cui, W.; Qi, F.; Zhang, Y.; Cao, H.; Zhang, J.; Wang, R.; Shen, W. Methane-rich water induces cucumber adventitious rooting through heme oxygenase1/carbon monoxide and Ca2+ pathways. Plant Cell Rep. 2015, 34, 435–445. [Google Scholar] [CrossRef]
- Astier, J.; Lindermayr, C. Nitric oxide-dependent posttranslational modification in plants: An update. Int. J. Mol. Sci. 2012, 13, 15193–15208. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Plant Species | Abiotic Stress and Its Effect | H2 Roles under Stress | Reference |
|---|---|---|---|
| Alfalfa | Cd stress inhibited root elongation | Improving root growth, re-establishing glutathione homeostasis | [6] |
| Cabbage | Cd stress reduced the activities of the antioxidant enzyme | Enhancing the activities of the antioxidant enzyme | [8] |
| Cole | Cd stress affected the balance of glutathione | Governing reduced glutathione homeostasis | [72] |
| Alfalfa | Cd stress obviously inhibited alfalfa seedling growth | Attenuating damage in alfalfa seedlings, reducing oxidative damage | [73] |
| Alfalfa | Al stress increased NO production, inhibited root elongation | Improving seedling growth, decreasing NO production | [75] |
| Maize | Al stress inhibited seed germination, broke the ion balance | Alleviating Al toxicity, decreasing lipid peroxidation | [74] |
| Rice | Al stress enhanced oxidative damage | Alleviating germination inhibition, re-establishing redox homeostasis | [16] |
| Alfalfa | Hg stress promoted ROS production | Decreasing ROS production and alleviating oxidative stress | [76] |
| Arabidopsis | Salt stress increased ion outflow | Maintaining ion homeostasis, controlling sodium exclusion | [21] |
| Rice | Salt inhibited seed germination | The alleviation of oxidative damage | [20] |
| Cucumber | Temperature stress affected photosynthetic parameters | Altering photosynthetic gas exchange | [78] |
| Rice | Temperature stress destroyed redox homeostasis | Re-establishing redox homeostasis | [22] |
| Radish | UV-A stress reduced anthocyanin content | Upregulating the anthocyanin biosynthesis-related genes | [79] |
| Alfalfa | UV-B stress destroyed the antioxidant defense system | Reducing lipid peroxidation, regulating the antioxidant defence system | [80] |
| Radish | Short wavelength light stress influenced anthocyanin biosynthesis | Enriching anthocyanin content | [81] |
| Alfalfa | Oxidative stress enhanced oxidative damage | Increasing levels of the MsHO-1 transcript, alleviating oxidative stress | [23] |
| Alfalfa | Drought stress destroyed the redox balance | Modulating stomatal sensitivity, reducing transpirational water loss | [82] |
| Alfalfa | Drought stress affected the enzyme activity | Elevating H2O2 levels, the inhibition of NADPH oxidase | [10] |
| Plant Species | Abiotic Stress and Its Effect | H2S Roles under Stress | Reference |
|---|---|---|---|
| Rice | Cd stress affected the stability of the membrane | Improving oxidative damage and maintaining ROS homeostasis | [85] |
| Foxtail millet | Cd stress broke the ion balance | Decreasing electrolytic leakage and enhancing photosynthesis | [84] |
| Pea | As stress damaged proteins and membranes | Increasing the level of NO, alleviating oxidative damage | [83] |
| Rice | Hg stress promoted ROS production | Improving the transcription of bZIP60, alleviating Hg toxicity | [4] |
| Cauliflower | Pb stress destroyed GSH levels | Elevating nonprotein thiols and total GSH levels | [33] |
| Zucchini | Ni stress reduced antioxidant enzyme activity | Enhancing antioxidant enzyme activity and reducing Pro contents | [34] |
| Wheat | Salt stress inhibited growth of wheat | Decreasing the Na+ concentration, alleviating the growth inhibition of wheat | [86] |
| Cucumber | Salt stress induced oxidative stress | Maintaining Na+ and K+ homeostasis | [87] |
| Broad bean | Salt stress affected stomatal sensitivity | Inducing stomatal closure, promoting H2O2 production | [88] |
| Cucumber | Salt stress broke the redox balance | Alleviating oxidative damage, upregulating the CsNMAPK transcript level | [89] |
| Grape | Low temperature stress affected the plasma membrane stability | Improving SOD activity and the plasma membrane stability of grape | [29] |
| Banana | Low temperature disrupted ion stability | Maintaining a higher peel firmness, reducing accumulation of MDA | [90] |
| Banana | Low temperature stress broke the redox balance | Inhibiting electrolyte leakage and reducing ethylene production | [91] |
| Hawthorn | Low temperature stress decreased antioxidant enzyme activity | Promoting phenols accumulation and enhancing antioxidant enzyme activity | [37] |
| Cucumber | Low temperature stress influenced the expression of related genes | Upregulating the expression of Cucurbitacin C synthetase-encoding genes | [30] |
| Tobacco | Heat temperature stress decreased vitality of cells | Improving vitality of cells and alleviating electrolyte leakage | [92] |
| Poplar | Heat temperature stress reduced S-nitrosoglutathione reductase activity | Increasing S-nitrosoglutathione reductase activity and reducing reactive oxygen/nitrogen damage | [31] |
| Soybean | Drought stress affected plant photosynthesis | Enhancing chlorophyll contents and decreasing the production of H2O2 | [93] |
| Arabidopsis | Drought stress changed the expression of drought associated genes | Stimulating the expression of drought associated genes | [27] |
| Arabidopsis | Drought stress influenced the transcriptional expression of the ABA receptor | Decreasing transcriptional expression of ABA receptor | [94] |
| Wheat | Drought stress changed MDA contents | Increasing antioxidant enzymes activity and reducing MDA contents | [28] |
| Wheat | Osmotic stress destroyed cysteine homeostasis | Sustaining antioxidant enzymes and cysteine homeostasis | [35] |
| Plant Species | Abiotic Stress and Its Effect | NO Roles under Stress | Reference |
|---|---|---|---|
| Lichen | Cd stress decreased the content of ionic permeate | Regulating ROS balance, increasing Pro and AsA contents | [50] |
| Peanut | Al stress promoted the production of harmful substances | Upregulating AhHsp70 expression and reducing cytochrome c release | [48] |
| Wheat | Al stress destroyed the antioxidant defense system | Enhancing antioxidant defense, improving H2O2 levels | [5] |
| Wheat | Al stress inhibited auxin flow | Improving the oxidized protein levels, guaranteeing normal indole-3-acetic acid flow | [47] |
| Mangrove | Salt stress induced lipid peroxidation | Reducing hydrogen peroxide content and lipid peroxidation | [96] |
| Arabidopsis | Salt stress broke the ion balance | Downregulating the expression of PIN genes, stabilizing IAA17 | [43] |
| Tobacco | Salt affected the activity of antioxidant enzymes | Enhancing the activity of antioxidant enzymes and H2S levels | [42] |
| Chickpea | Salt stress increased electrolyte Leakage and the levels of osmolytes | Enhancing the biosyntheses of antioxidant enzymes | [97] |
| Mustard | Salt stress accelerated oxidative damage | Regulating oxidative stress and photosynthetic performance | [7] |
| Mustard | Salt stress influenced ion balance | Decreasing electrolytic leakage and K+/Na+ ratio | [98] |
| Pisum sativum L. | Salt stress triggered the membrane lipid peroxidation | Reducing accumulations of ROS and MDA | [99] |
| Jatropha curcas | Salt stress accelerated toxic ion accumulation | Ameliorating oxidative damage and toxic ion accumulation | [100] |
| Wheat | Salt stress reduced biomass production and grain yield | Enhancing physiological and biochemical parameters | [1] |
| Wheat | Temperature stress induced oxidative damage | Enhancing the accumulation of gliadin protein and starch | [101] |
| Cherry | Temperature stress destroyed membrane integrity | Maintaining antioxidant system activity and membrane integrity | [102] |
| Marigold | Drought stress induced carbohydrate and nitrogen accumulation | Increasing chlorophyll content and protein content | [45] |
| Wheat | Osmotic stress created oxidative damage | Enhancing the antioxidant defense system, reducing the methyl-glyoxal content | [44] |
| White clover | Drought stress influenced metabolic regulation and transform | Inducing changes of metabolic profiles | [103] |
| Alfalfa | Drought stress inhibited growth physiological processes | Alleviating loss of water content and embryo axis elongation | [104] |
| Plant Species | Abiotic Stress and Its Effect | CO Roles under Stress | Reference |
|---|---|---|---|
| Alfalfa | Cd stress destroyed antioxidation enzymatic activities | Modulating glutathione metabolism | [57] |
| Alfalfa | Cd induced a loss of plasma membrane integrity, lipid peroxidation | Upregulating expression of HO-1 gene | [106] |
| Rapeseed | Hg stress inhibited growth and development | Improving antioxidation capacity and expression of BnHO-1 | [3] |
| Mustard | Hg triggered production of O2⋅− and H2O2, as well as peroxides | Improving antioxidative enzymes, reducing oxidative stress | [107] |
| Rice | Zn stress inhibited root elongation | Downregulating of the expression of homeostasis-related genes | [108] |
| Wheat | Salt stress induced oxidative damage | Enhancing the activities of antioxidant enzymes | [109] |
| Wheat | Salt stress caused oxidative damage | Counteracting lipid peroxidation | [110] |
| Rice | Salt stress inhibited seed germination | Alleviating oxidative damage | [111] |
| Wheat | Salt stress reduced antioxidant enzyme activities | Decreasing of superoxide anion overproduction | [112] |
| Cassia obtusifolia L. | Salt stress lowered chlorophyll concentration | Alleviating oxidative damage, improving membrane permeability | [113] |
| Soybean | Salt stress affected the parameters of lipid peroxidation | Improving lipid peroxidation and ureide metabolism | [58] |
| Wheat | Osmotic stress-induced seed germination inhibition | Increasing in the activities of amylase and antioxidant enzyme | [114] |
| Rice | Drought stress inhibited HO activity | Improving the level of HO-1 gene expression and HO activity | [115] |
| Canola | Temperature stress delayed plant development | Enhancing the expression of BnDHN types gene | [15] |
| Plant Species | Abiotic Stress and Its Effect | CH4 Roles under Stress | Reference |
|---|---|---|---|
| Alfalfa | Al stress influenced the physiological roles of alfalfa | Enhancing resistance seedlings, regulating organic acid metabolism | [13] |
| Alfalfa | Cu-triggered oxidative stress | Increasing amylase activities, reducing Cu accumulation | [67] |
| Alfalfa | Cd stress decreased the ratio of reduced/oxidized (homo)glutathione | Re-establishing glutathione homeostasis, reducing lipid peroxidation | [68] |
| Alfalfa | Salt reduced the activities of representative antioxidant enzymes | Reducing reactive oxygen species over accumulation | [65] |
| Maize | Osmotic stress decreased biomass and relative water contents | Modulating sugar and AsA metabolism | [66] |
| Mung bean | Osmotic stress broke the ion balance | Re-establishing redox balance, alleviating seed germination inhibition | [46] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.; Yang, Y.; Li, C.; Huang, D.; Zhang, J.; Wang, C.; Li, W.; Wang, N.; Deng, Y.; Liao, W. Research Progress on the Functions of Gasotransmitters in Plant Responses to Abiotic Stresses. Plants 2019, 8, 605. https://doi.org/10.3390/plants8120605
Yao Y, Yang Y, Li C, Huang D, Zhang J, Wang C, Li W, Wang N, Deng Y, Liao W. Research Progress on the Functions of Gasotransmitters in Plant Responses to Abiotic Stresses. Plants. 2019; 8(12):605. https://doi.org/10.3390/plants8120605
Chicago/Turabian StyleYao, Yandong, Yan Yang, Changxia Li, Dengjing Huang, Jing Zhang, Chunlei Wang, Weifang Li, Ni Wang, Yuzheng Deng, and Weibiao Liao. 2019. "Research Progress on the Functions of Gasotransmitters in Plant Responses to Abiotic Stresses" Plants 8, no. 12: 605. https://doi.org/10.3390/plants8120605