Secoiridoids Metabolism Response to Wounding in Common Centaury (Centaurium erythraea Rafn) Leaves


 ,
,  , and
, and
Abstract
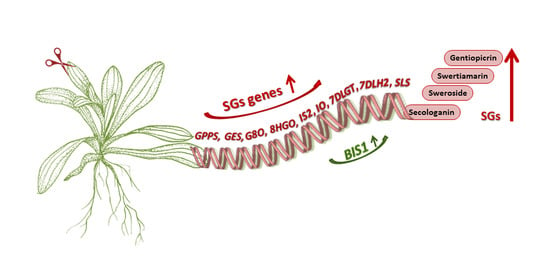
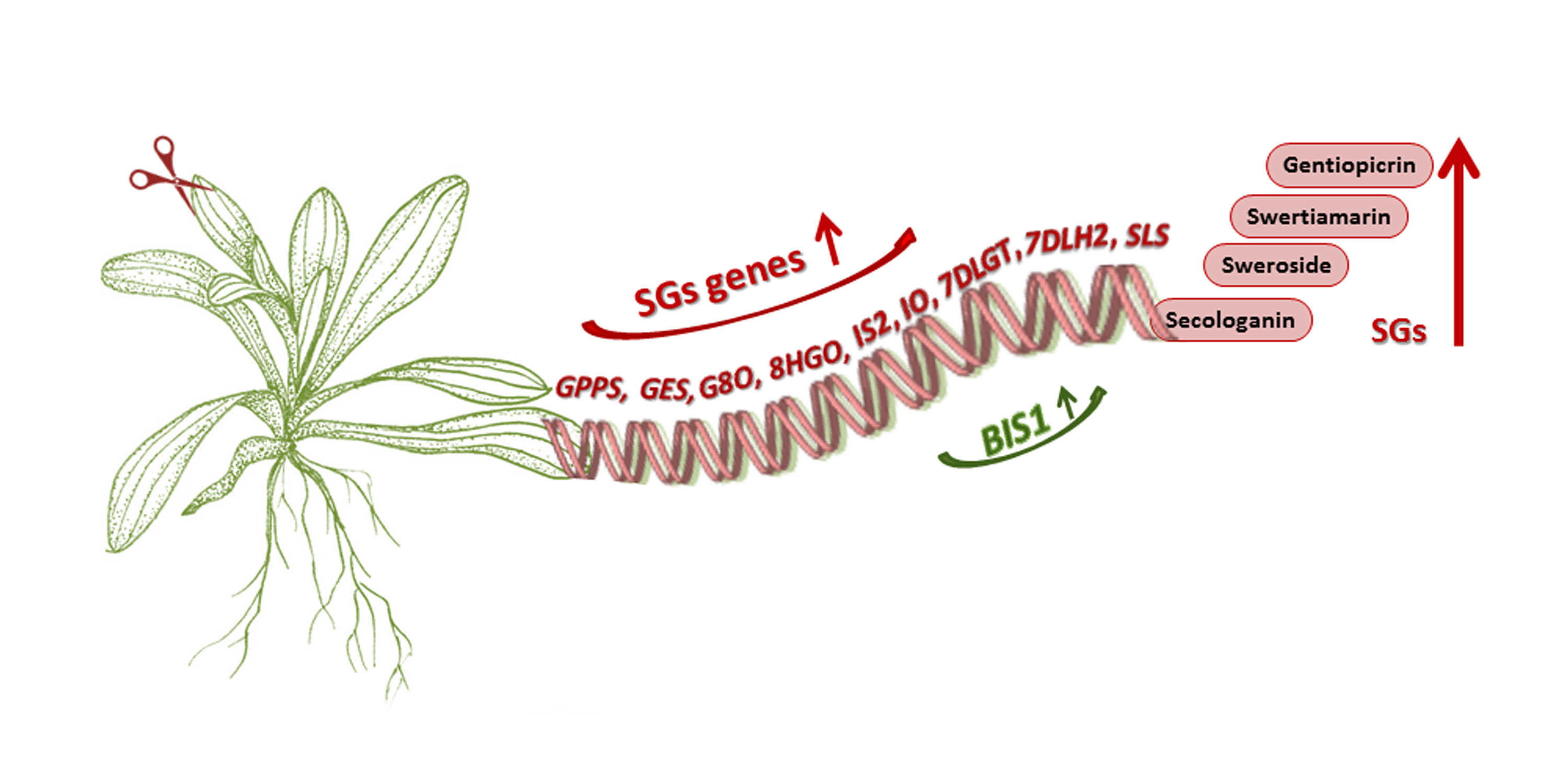
:
1. Introduction
2. Results and Discussion
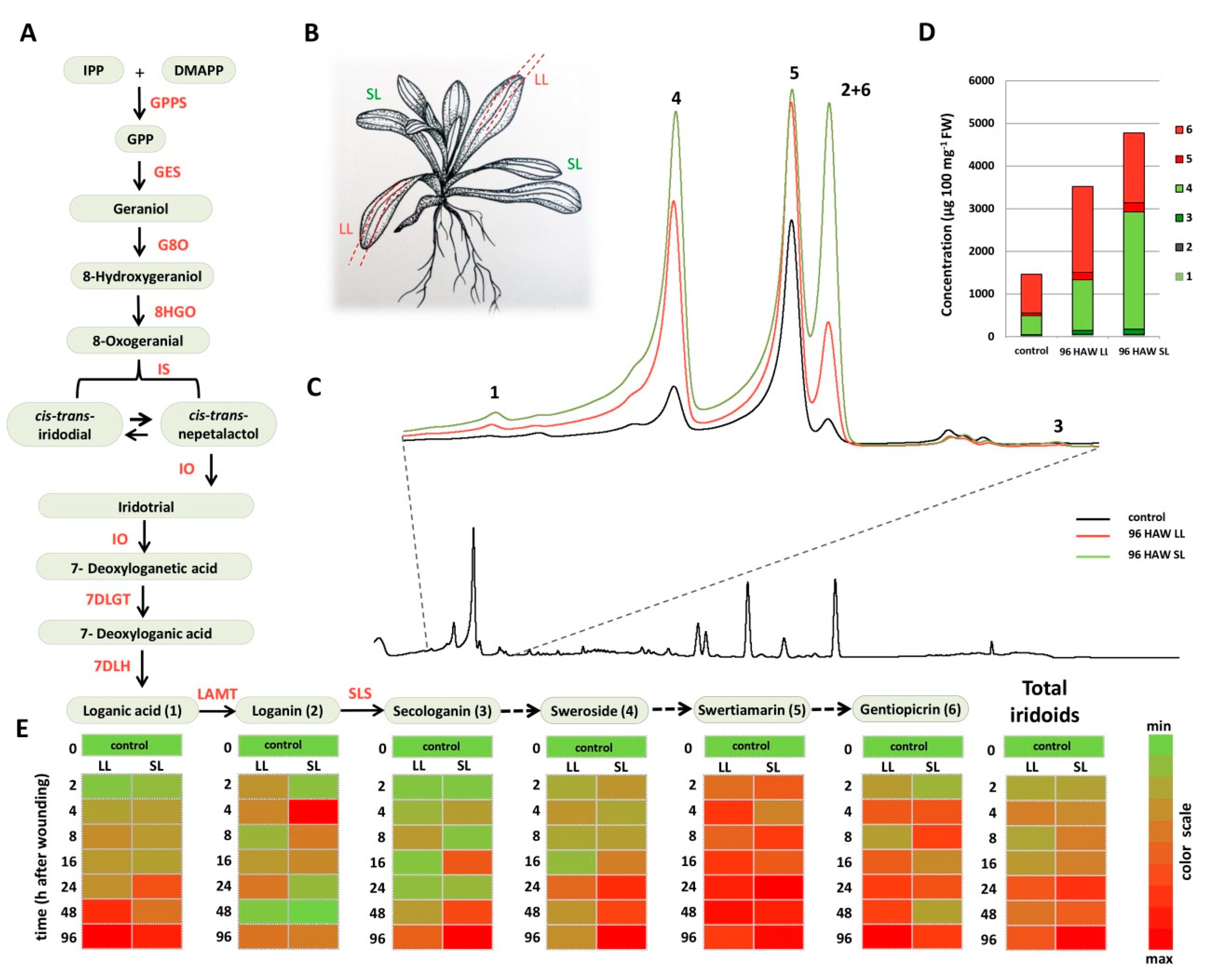
2.1. Wounding-Induced Alterations in the SGs Content
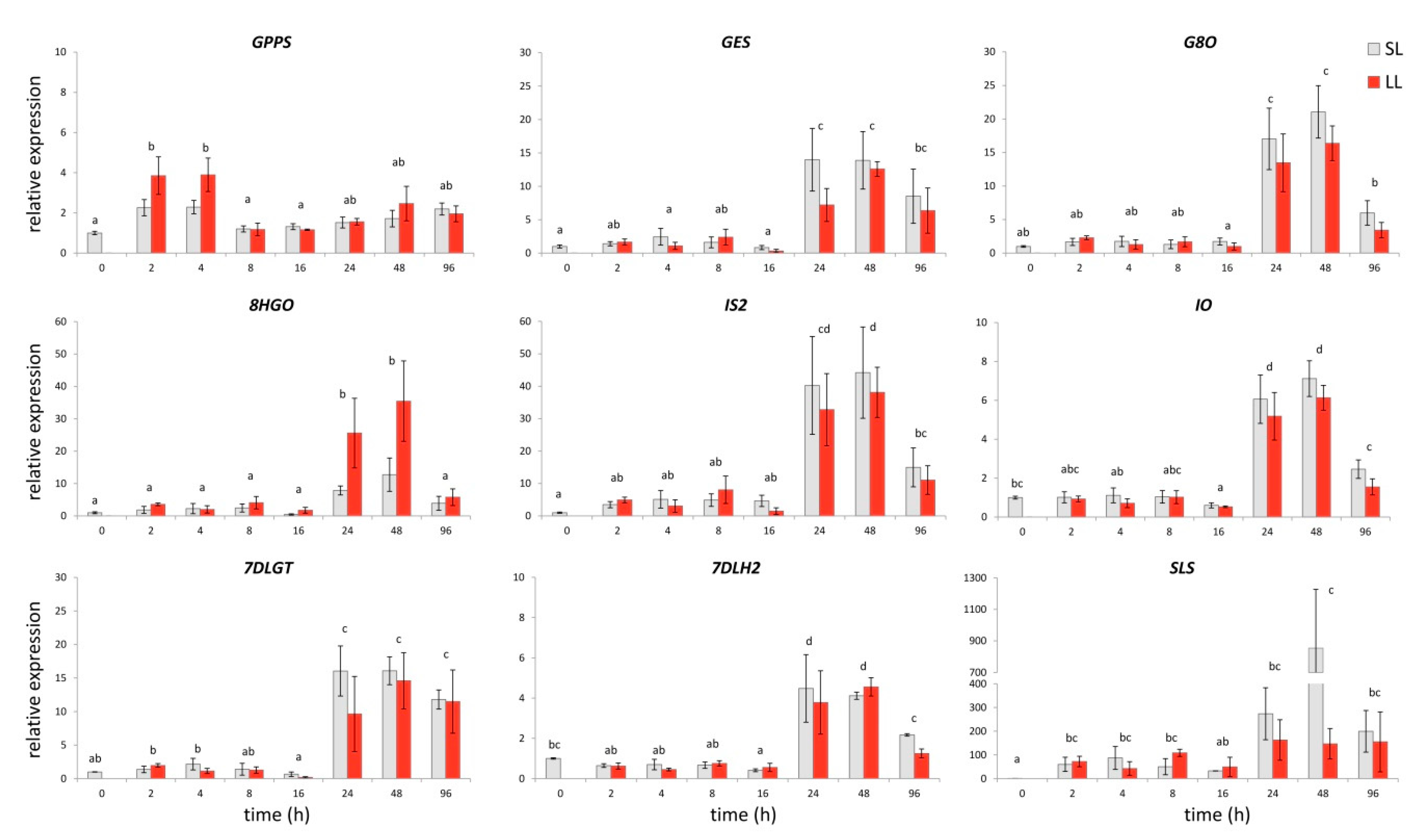
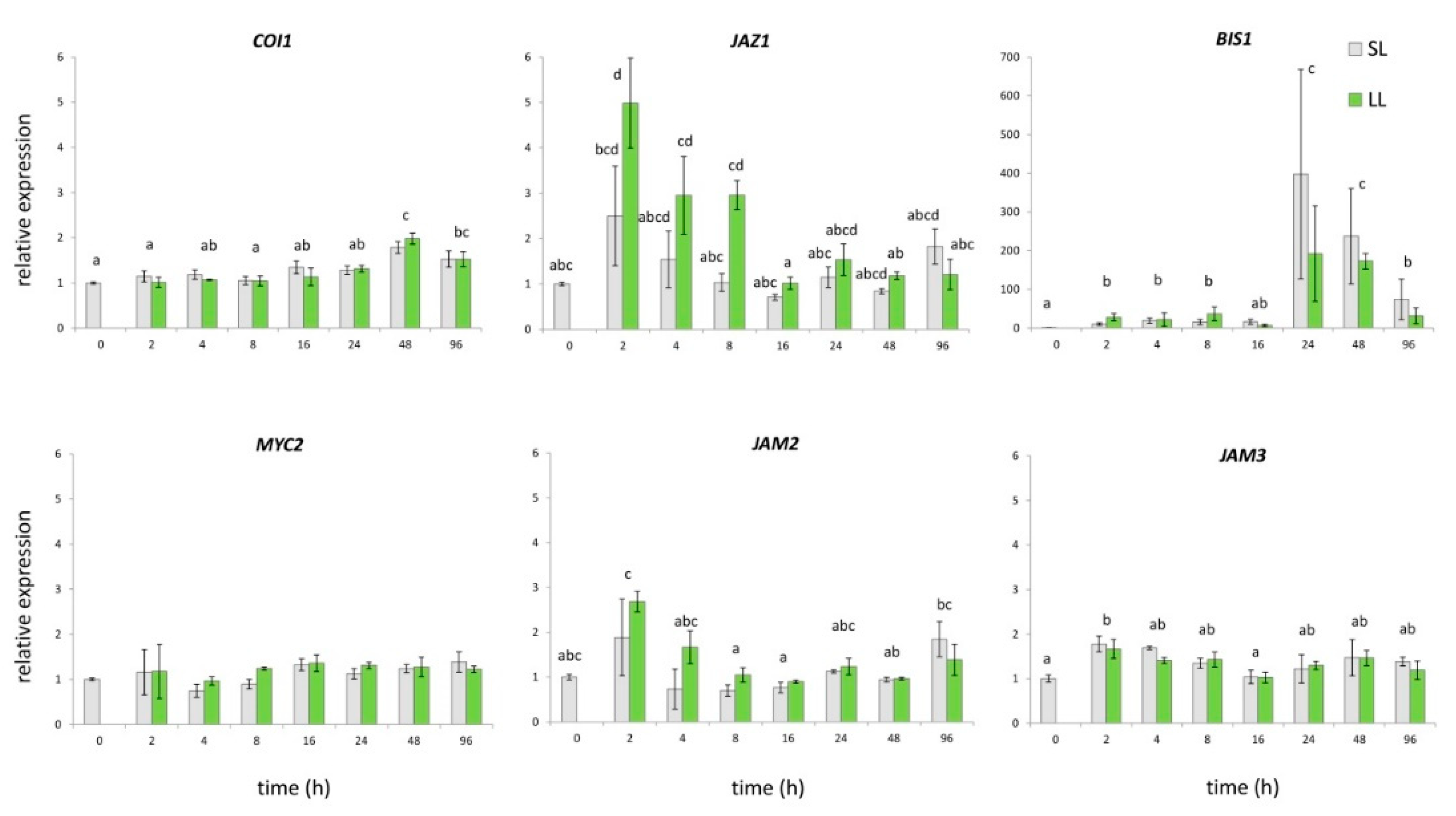
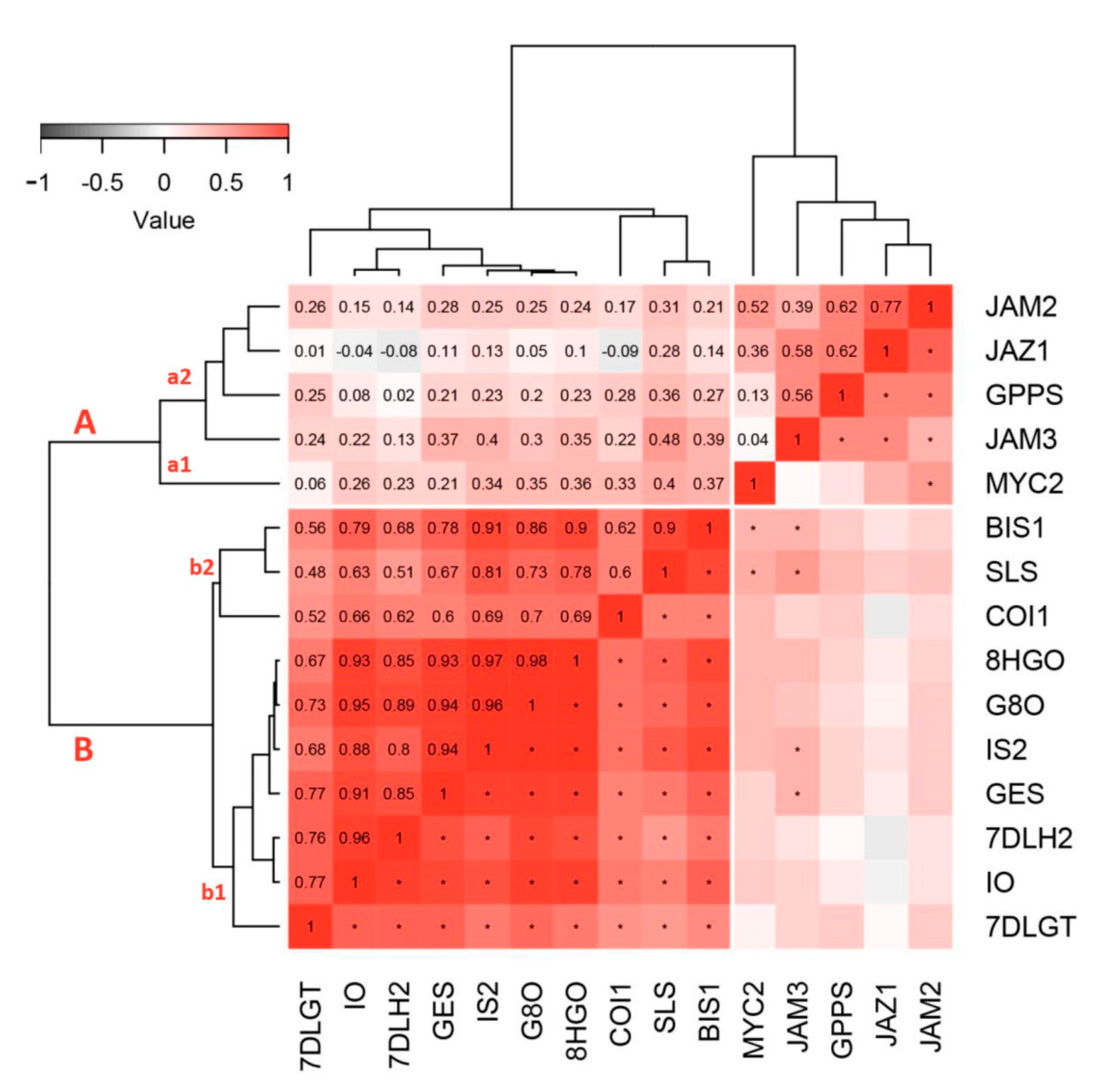
2.2. Secoiridoids-Related Biosynthetic Genes Expression Profiles in Response to Wounding
3. Materials and Methods
3.1. Plant Material
3.2. Experimental Setup
3.3. Plant Methanol Extracts Preparation
3.4. Ultra-High Performance Liquid Chromatography–Tandem Mass Spectrometry (UHPLC–MS/MS) Analysis
3.5. Gene Expression Analysis
3.6. Phylogenetic Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zlatković, B.K.; Bogosavljević, S.S.; Radivojević, A.R.; Pavlović, M.A. Traditional use of the native medicinal plant resource of Mt. Rtanj (Eastern Serbia): Ethnobotanical evaluation and comparison. J. Ethnopharmacol. 2014, 151, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Šiler, B.; Mišić, D. Biologically Active Compounds from the Genus Centaurium sl (Gentianaceae): Current Knowledge and Future Prospects in Medicine. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Nedherlands, 2016; pp. 363–397. [Google Scholar]
- Božunović, J.; Živković, S.; Gašić, U.; Glamočlija, J.; Ćirić, A.; Matekalo, D.; Šiler, B.; Soković, M.; Tešić, Ž.; Mišić, D. In vitro and in vivo transformations of Centaurium erythraea secoiridoid glucosides alternate their antioxidant and antimicrobial capacity. Ind. Crops Prod. 2018, 111, 705–721. [Google Scholar] [CrossRef]
- Matekalo, D.; Skorić, M.; Nikolić, T.; Novaković, L.; Lukić, M.; Božunović, J.; Aničić, N.; Filipović, B.; Mišić, D. Organ-specific and genotype-dependent constitutive biosynthesis of secoiridoid glucosides in Centaurium erythraea Rafn, and its elicitation with methyl jasmonate. Phytochemistry 2018, 155, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Šiler, B.; Živković, S.; Banjanac, T.; Cvetković, J.; Nestorović Živković, J.; Ćirić, A.; Soković, M.; Mišić, D. Centauries as underestimated food additives: Antioxidant and antimicrobial potential. Food Chem. 2014, 147, 367–376. [Google Scholar] [CrossRef]
- Kachmar, M.R.; Oliveira, A.P.; Valentão, P.; Gil-Izquierdo, A.; Domínguez-Perles, R.; Ouahbi, A.; El Badaoui, K.; Andrade, P.B.; Ferreres, F. HPLC-DAD-ESI/MS n phenolic profile and in vitro biological potential of Centaurium erythraea Rafn aqueous extract. Food Chem. 2019, 278, 424–433. [Google Scholar] [CrossRef]
- St-Pierre, B.; Besseau, S.; Clastre, M.; Courdavault, V.; Courtois, M.; Crèche, J.; Ducos, E.; de Bernonville, T.D.; Dutilleul, C.; Glévarec, G.; et al. Deciphering the evolution, cell biology and regulation of monoterpene indole alkaloids. In Advances in Botanical Research; Giglioli-Guivarc’h, N., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 68, pp. 73–109. [Google Scholar] [CrossRef]
- Miettinen, K.; Dong, L.; Navrot, N.; Schneider, T.; Burlat, V.; Pollier, J.; Woittiez, L.; van der Krol, S.; Lugan, R.; Ilc, T.; et al. The seco-iridoid pathway from Catharanthus roseus. Nat. Commun. 2014, 5, 3606. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; Miettinen, K.; Claudel, P.; Burlat, V.; Guirimand, G.; Courdavault, V.; Papon, N.; Meyer, S.; Godet, S.; St-Pierre, B.; et al. Characterization of the plastidial geraniol synthase from Madagascar periwinkle which initiates the monoterpenoid branch of the alkaloid pathway in internal phloem associated parenchyma. Phytochemistry 2013, 85, 36–43. [Google Scholar] [CrossRef]
- Pan, Q.; Mustafa, N.R.; Tang, K.; Choi, Y.H.; Verpoorte, R. Monoterpenoid indole alkaloids biosynthesis and its regulation in Catharanthus roseus: A literature review from genes to metabolites. Phytochem. Rev. 2016, 15, 221–250. [Google Scholar] [CrossRef]
- Kubo, I.; Matsumoto, A.; Takase, I. A multichemical defense mechanism of bitter olive Olea europaea (Oleaceae). J. Chem. Ecol. 1985, 11, 251–263. [Google Scholar] [CrossRef]
- Bowers, M.D.; Puttick, M.G. Response of generalist and specialist insects to qualitative allelochemical variation. J. Chem. Ecol. 1988, 14, 319–334. [Google Scholar] [CrossRef]
- Puttick, M.G.; Bowers, M.D. Effect of qualitative and quantitative variation in allelochemicals on a generalist insect: Iridoid glycosides and the southern armyworm. J. Chem. Ecol. 1988, 14, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Marak, H.B.; Biere, A.; Van Damme, J.M.M. Two herbivore-deterrent iridoid glycosides reduce the in-vitro growth of a specialist but not of a generalist pathogenic fungus of Plantago lanceolata L. Chemoecology 2002, 12, 185–192. [Google Scholar] [CrossRef]
- Marak, H.B.; Biere, A.; Van Damme, J.M.M. Systemic, genotype-specific induction of two herbivore-deterrent iridoid glycosides in Plantago lanceolata L. in response to fungal infection by Diaporthe adunca (ROB.) niessel. J. Chem. Ecol. 2002, 28, 2429–2448. [Google Scholar] [CrossRef] [PubMed]
- León, E.; Rojo, J.; Sánchez-Serrano, J. Wound signalling in plants. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Radušienė, J. Biological peculiarities and possibilities of introduction of Centaurium erythraea Rafn. Biologija 1995, 3, 55–57. [Google Scholar]
- Van Rossum, F. Succession stage variation in population size in an early-successional herb in a peri-urban forest. Acta Oecologica 2009, 35, 261–268. [Google Scholar] [CrossRef]
- Malook, S.U.; Qi, J.; Hettenhausen, C.; Xu, Y.; Zhang, C.; Zhang, J.; Lu, C.; Li, J.; Wang, L.; Wu, J. The oriental armyworm (Mythimna separata) feeding induces systemic defence responses within and between maize leaves. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180307. [Google Scholar] [CrossRef] [Green Version]
- Delessert, C.; Wilson, I.; Van Der Straeten, D.; Dennis, E.; Dolferus, R. Spatial and temporal analysis of the local response to wounding. Plant Mol. Biol. 2004, 55, 165–181. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; González-Aguëro, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding stress response. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef] [Green Version]
- Reymond, P.; Weber, H.; Damond, M.; Farmer, E.E. Differential Gene Expression in Response to Mechanical Wounding and Insect Feeding in Arabidopsis. Plant Cell 2000, 12, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef] [PubMed]
- De Bernonville, T.D.; Carqueijeiro, I.; Lanoue, A.; Lafontaine, F.; Sánchez Bel, P.; Liesecke, F.; Musset, K.; Oudin, A.; Glévarec, G.; Pichon, O.; et al. Folivory elicits a strong defense reaction in Catharanthus roseus: Metabolomic and transcriptomic analyses reveal distinct local and systemic responses. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Colinas, M.; Goossens, A. Combinatorial Transcriptional Control of Plant Specialized Metabolism. Trends Plant Sci. 2018, 23, 324–336. [Google Scholar] [CrossRef]
- Pauwels, L.; Goossens, A. The JAZ Proteins: A Crucial Interface in the Jasmonate Signaling Cascade. Plant Cell 2011, 23, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2019, 48, 1–11. [Google Scholar] [CrossRef]
- Schweizer, F.; Colinas, M.; Pollier, J.; Van Moerkercke, A.; Vanden Bossche, R.; de Clercq, R.; Goossens, A. An engineered combinatorial module of transcription factors boosts production of monoterpenoid indole alkaloids in Catharanthus roseus. Metab. Eng. 2018, 48, 150–162. [Google Scholar] [CrossRef]
- Van Moerkercke, A.; Steensma, P.; Schweizer, F.; Pollier, J.; Gariboldi, I.; Payne, R.; Vanden Bossche, R.; Miettinen, K.; Espoz, J.; Purnama, P.C.; et al. The bHLH transcription factor BIS1 controls the iridoid branch of the monoterpenoid indole alkaloid pathway in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2015, 112, 8130–8135. [Google Scholar] [CrossRef] [Green Version]
- Van Moerkercke, A.; Steensma, P.; Gariboldi, I.; Espoz, J.; Purnama, P.C.; Schweizer, F.; Miettinen, K.; Vanden Bossche, R.; De Clercq, R.; Memelink, J.; et al. The basic helix-loop-helix transcription factor BIS2 is essential for monoterpenoid indole alkaloid production in the medicinal plant Catharanthus roseus. Plant J. 2016, 88, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Sasaki-Sekimoto, Y.; Jikumaru, Y.; Obayashi, T.; Saito, H.; Masuda, S.; Kamiya, Y.; Ohta, H.; Shirasu, K. Basic Helix-Loop-Helix Transcription Factors JASMONATE-ASSOCIATED MYC2-LIKE1 (JAM1), JAM2, and JAM3 Are Negative Regulators of Jasmonate Responses in Arabidopsis. Plant Physiol. 2013, 163, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Patra, B.; Pattanaik, S.; Schluttenhofer, C.; Yuan, L. A network of jasmonate-responsive bHLH factors modulate monoterpenoid indole alkaloid biosynthesis in Catharanthus roseus. New Phytol. 2018, 217, 1566–1581. [Google Scholar] [CrossRef] [Green Version]
- Cisneros-Zevallos, L. The Use of Controlled Postharvest Abiotic Stresses as a Tool for Enhancing the Nutraceutical Content and Adding-Value. J. Food Sci. 2003, 68, 1560–1565. [Google Scholar] [CrossRef]
- Villarreal-García, D.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as Biofactories: Postharvest Stress-Induced Accumulation of Phenolic Compounds and Glucosinolates in Broccoli Subjected to Wounding Stress and Exogenous Phytohormones. Front. Plant Sci. 2016, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.R.; Schripsema, J. Chemotaxonomy and pharmacology of Gentianaceae. In Gentianaceae-Systematics and Natural History; Struwe, L., Albert, V., Eds.; Cambridge University Press: London, UK, 2002; pp. 573–631. [Google Scholar]
- Cao, X.Y.; Wang, Z.J.; Wang, Z.Z. Comparative analysis of contents of four iridoid glucosides in different organs of four species of Gentiana L. J. Plant Resour. Environ. 2012, 21, 58–63. [Google Scholar]
- Padhan, J.K.; Kumar, V.; Sood, H.; Singh, T.R.; Chauhan, R.S. Contents of therapeutic metabolites in Swertia chirayita correlate with the expression profiles of multiple genes in corresponding biosynthesis pathways. Phytochemistry 2015, 116, 38–47. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Guo, F.; Zhan, L.; Mohr, T.; Cheng, P.; Huo, N.; Gu, R.; Pei, D.; Sun, J.; et al. Deep sequencing and transcriptome analyses to identify genes involved in secoiridoid biosynthesis in the Tibetan medicinal plant Swertia mussotii. Sci. Rep. 2017, 7, 43108. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, J.; Shen, T.; Zhao, Y.L.; Zuo, Z.T.; Wang, Y.Z.; Li, W.Y. Investigation of chemical diversity in different parts and origins of ethnomedicine Gentiana rigescens Franch using targeted metabolite profiling and multivariate statistical analysis. Biomed. Chromatogr. 2016, 30, 232–240. [Google Scholar] [CrossRef]
- Šiler, B.; Avramov, S.; Banjanac, T.; Cvetković, J.; Nestorović Živković, J.; Patenković, A.; Mišić, D. Secoiridoid glycosides as a marker system in chemical variability estimation and chemotype assignment of Centaurium erythraea Rafn from the Balkan Peninsula. Ind. Crops Prod. 2012, 40, 336–344. [Google Scholar] [CrossRef]
- Nishanth, M.J.; Sheshadri, S.A.; Rathore, S.S.; Srinidhi, S.; Simon, B. Expression analysis of Cell wall invertase under abiotic stress conditions influencing specialized metabolism in Catharanthus roseus. Sci. Rep. 2018, 8, 15059. [Google Scholar] [CrossRef] [Green Version]
- Darrow, K.; Bowers, M.D. Effects of herbivore damage and nutrient level on induction of iridoid glycosides in Plantago lanceolata. J. Chem. Ecol. 1999, 25, 1427–1440. [Google Scholar] [CrossRef]
- Meelaph, T.; Kobtrakul, K.; Chansilpa, N.N.; Han, Y.; Rani, D.; De-Eknamkul, W.; Vimolmangkang, S. Coregulation of Biosynthetic Genes and Transcription Factors for Aporphine-Type Alkaloid Production in Wounded Lotus Provides Insight into the Biosynthetic Pathway of Nuciferine. ACS Omega 2018, 3, 8794–8802. [Google Scholar] [CrossRef] [PubMed]
- Banjanac, T.; Dragićević, M.; Šiler, B.; Gašić, U.; Bohanec, B.; Nestorović Živković, J.; Trifunović, S.; Mišić, D. Chemodiversity of two closely related tetraploid Centaurium species and their hexaploid hybrid: Metabolomic search for high-resolution taxonomic classifiers. Phytochemistry 2017, 140, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Kucharska, A.Z.; Fecka, I. Identification of iridoids in edible honeysuckle berries (Lonicera caerulea L. var. kamtschatica Sevast.) by UPLC-ESI-qTOF-MS/MS. Molecules 2016, 21, 1157. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, M.; Lin, Z.; Jiang, H.; Niu, Y.; Wang, H.; Chen, S. Comprehensive identification of 125 multifarious constituents in Shuang-huang-lian powder injection by HPLC-DAD-ESI-IT-TOF-MS. J. Pharm. Biomed. Anal. 2015, 115, 86–106. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A.; Gilroy, S. Glutamate triggers long-distance, calcium-based plant defense signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Vázquez-Flota, F.; Carrillo-Pech, M.; Minero-García, Y.; De Lourdes Miranda-Ham, M. Alkaloid metabolism in wounded Catharanthus roseus seedlings. Plant Physiol. Biochem. 2004, 42, 623–628. [Google Scholar] [CrossRef]
- Jablonická, V.; Ziegler, J.; Vatehová, Z.; Lišková, D.; Heilmann, I.; Obložinský, M.; Heilmann, M. Inhibition of phospholipases influences the metabolism of wound-induced benzylisoquinoline alkaloids in Papaver somniferum L. J. Plant Physiol. 2018, 223, 1–8. [Google Scholar] [CrossRef]
- Alves, M.N.; Sartoratto, A.; Trigo, J.R. Scopolamine in Brugmansia suaveolens (Solanaceae): Defense, allocation, costs, and induced response. J. Chem. Ecol. 2007, 33, 297–309. [Google Scholar] [CrossRef]
- Kidd, T.; Easson, M.L.; Qu, Y.; Jones, G.; De Luca, V. Inter-organ transport of secologanin allows assembly of monoterpenoid indole alkaloids in a Catharanthus roseus mutant. Phytochemistry 2019, 159, 119–126. [Google Scholar] [CrossRef]
- Munkert, J.; Pollier, J.; Miettinen, K.; Van Moerkercke, A.; Payne, R.; Müller-Uri, F.; Burlat, V.; O’Connor, S.E.; Memelink, J.; Kreis, W.; et al. Iridoid synthase activity is common among the plant progesterone 5β-reductase family. Mol. Plant 2015, 8, 136–152. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Sen, J.; Deswal, R. Downregulation of terpenoid indole alkaloid biosynthetic pathway by low temperature and cloning of a AP2 type C-repeat binding factor (CBF) from Catharanthus roseus (L). G. Don. Plant Cell Rep. 2007, 26, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, D.; Wang, Y.; Xie, D. Jasmonate action in plant defense against insects. J. Exp. Bot. 2019, 70, 3391–3400. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Fernández-Calvo, P.; Schweizer, F.; Goossens, A. Jasmonates: Signal transduction components and their roles in environmental stress responses. Plant Mol. Biol. 2016, 91, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Koo, A.J.K.; Gao, X.; Daniel Jones, A.; Howe, G.A. A rapid wound signal activates the systemic synthesis of bioactive jasmonates in Arabidopsis. Plant J. 2009, 59, 974–986. [Google Scholar] [CrossRef]
- Sanchez-Serrano, J.J. Plant Responses to Wounding; eLS, John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 1–7. [Google Scholar] [CrossRef]
- Malkov, S.; Simonovic, A. Shotgun assembly of Centaurium erythraea transcriptome. In Proceedings of the 19th Symposium of the Serbian Plant Physiology Society, Banja Vrujci, Serbia, 13–15 June 2011; p. 16. [Google Scholar]
- Chini, A.; Gimenez-Ibanez, S.; Goossens, A.; Solano, R. Redundancy and specificity in jasmonate signalling. Curr. Opin. Plant Biol. 2016, 33, 147–156. [Google Scholar] [CrossRef]
- Cao, X.; Guo, X.; Yang, X.; Wang, H.; Hua, W.; He, Y.; Kang, J.; Wang, Z. Transcriptional responses and gentiopicroside biosynthesis in methyl jasmonate-treated Gentiana macrophylla seedlings. PLoS ONE 2016, 11, e0166493. [Google Scholar] [CrossRef]
- Mertens, J.; Pollier, J.; Vanden Bossche, R.; Lopez-Vidriero, I.; Franco-Zorrilla, J.M.; Goossens, A. The bHLH Transcription Factors TSAR1 and TSAR2 Regulate Triterpene Saponin Biosynthesis in Medicago truncatula. Plant Physiol. 2016, 170, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, Y.; Cai, Y.; Zhang, F.; Xia, G.; Xiang, F. Cloning and Functional Analysis of Geraniol 10-Hydroxylase, a Cytochrome P450 from Swertia mussotii Franch. Biosci. Biotechnol. Biochem. 2010, 74, 1583–1590. [Google Scholar] [CrossRef]
- Wang, C.T.; Liu, H.; Gao, X.S.; Zhang, H.X. Overexpression of G10H and ORCA3 in the hairy roots of Catharanthus roseus improves catharanthine production. Plant Cell Rep. 2010, 29, 887–894. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Primer3Plus. Available online: http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi (accessed on 8 November 2019).
- Gasic, K.; Hernandez, A.; Korban, S.S. RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction. Plant Mol. Biol. Report. 2004, 22, 437–438. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2 ΔΔ C T Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Box, G.E.P.; Cox, D.R. An analysis of transformations. J. R. Stat. Soc. Ser. B 1964, 26, 211–243. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. 2008. Available online: http://softlibre.unizar.es/manuales/aplicaciones/r/fullrefman.pdf (accessed on 8 November 2019).
- Venables, W.N.; Ripley, B.D. Modern applied statistics with S-PLUS; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. gplots: various R programming tools for plotting data. R package version 3.0. 1. 2016. Available online: https://rdrr.io/cran/gplots/ (accessed on 8 November 2019).
- Inoue, K.; Takeda, Y.; Tanahashi, T.; Inouye, H. Studies on Monoterpene Glucosides and Related Natural Products. XLII. On the Possibility of the Intermediacy of 10-Hydroxyloganin in the Biosynthesis of Secologanin. Chem. Pharm. Bull. 1981, 29, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Battersby, A.R.; Thompson, M.; Glüsenkamp, K.-H.; Tietze, L.-F. Untersuchngen zur Biogenese der Indolalkaloide. Synthese und Verfütterung radioaktive markierter Monoterpenaldehyde. Chem. Ber. 1981, 114, 3430–3438. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UHPLC-MS Data | UHPLC-DAD Data | ||||||
|---|---|---|---|---|---|---|---|
| Peak No. | Assignment | tR (min) | [M + CH3COOH + H]− [m/z] | SRM MS2 Fragments [m/z (Intensity)] | cE (eV) | tR (min) | λmax [nm] |
| 1 | Loganic acid S,R | 2.01 | 375 | 213 (100), 168 (<5) | 20 | 1.95 | 240 |
| 5 | Swertiamarin S,R | 2.61 | 433 | 179 (100); 161 (15) | 20 | 2.52 | 240 |
| 2 | Loganin S,R | 2.88 | 449 | 227 (85); 127 (100) | 30 | 2.85 | 240 |
| 6 | Gentiopicrin S,R | 2.90 | 415 | 179 (60); 119 (100) | 20 | 2.83 | 250, 280, 370 |
| 4 | Sweroside S,R | 3.00 | 417 | 195 (100); 179 (85) | 20 | 2.94 | 250 |
| 3 | Secologanin S,R | 3.66 | 447 | 155 (100); 123 (30) | 30 | 3.58 | 240 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Božunović, J.; Skorić, M.; Matekalo, D.; Živković, S.; Dragićević, M.; Aničić, N.; Filipović, B.; Banjanac, T.; Šiler, B.; Mišić, D. Secoiridoids Metabolism Response to Wounding in Common Centaury (Centaurium erythraea Rafn) Leaves. Plants 2019, 8, 589. https://doi.org/10.3390/plants8120589
Božunović J, Skorić M, Matekalo D, Živković S, Dragićević M, Aničić N, Filipović B, Banjanac T, Šiler B, Mišić D. Secoiridoids Metabolism Response to Wounding in Common Centaury (Centaurium erythraea Rafn) Leaves. Plants. 2019; 8(12):589. https://doi.org/10.3390/plants8120589
Chicago/Turabian StyleBožunović, Jelena, Marijana Skorić, Dragana Matekalo, Suzana Živković, Milan Dragićević, Neda Aničić, Biljana Filipović, Tijana Banjanac, Branislav Šiler, and Danijela Mišić. 2019. "Secoiridoids Metabolism Response to Wounding in Common Centaury (Centaurium erythraea Rafn) Leaves" Plants 8, no. 12: 589. https://doi.org/10.3390/plants8120589