The Effects of Endogenous Hormones on the Flowering and Fruiting of Glycyrrhiza uralensis

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Changes in Endogenous Hormones during the Growth of G. uralensis Fisch.
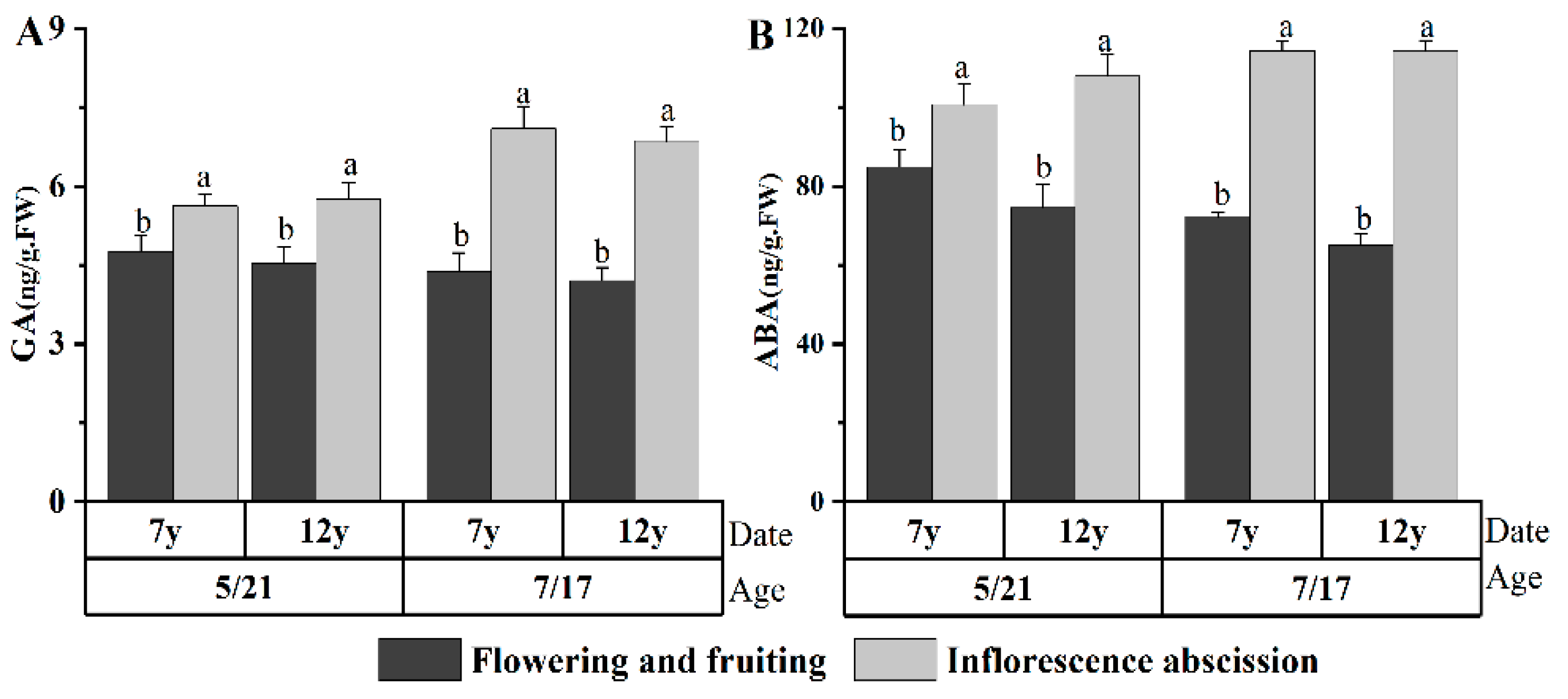
2.2. Changes in ZR and IAA Levels during Flower Bud Differentiation of G. uralensis
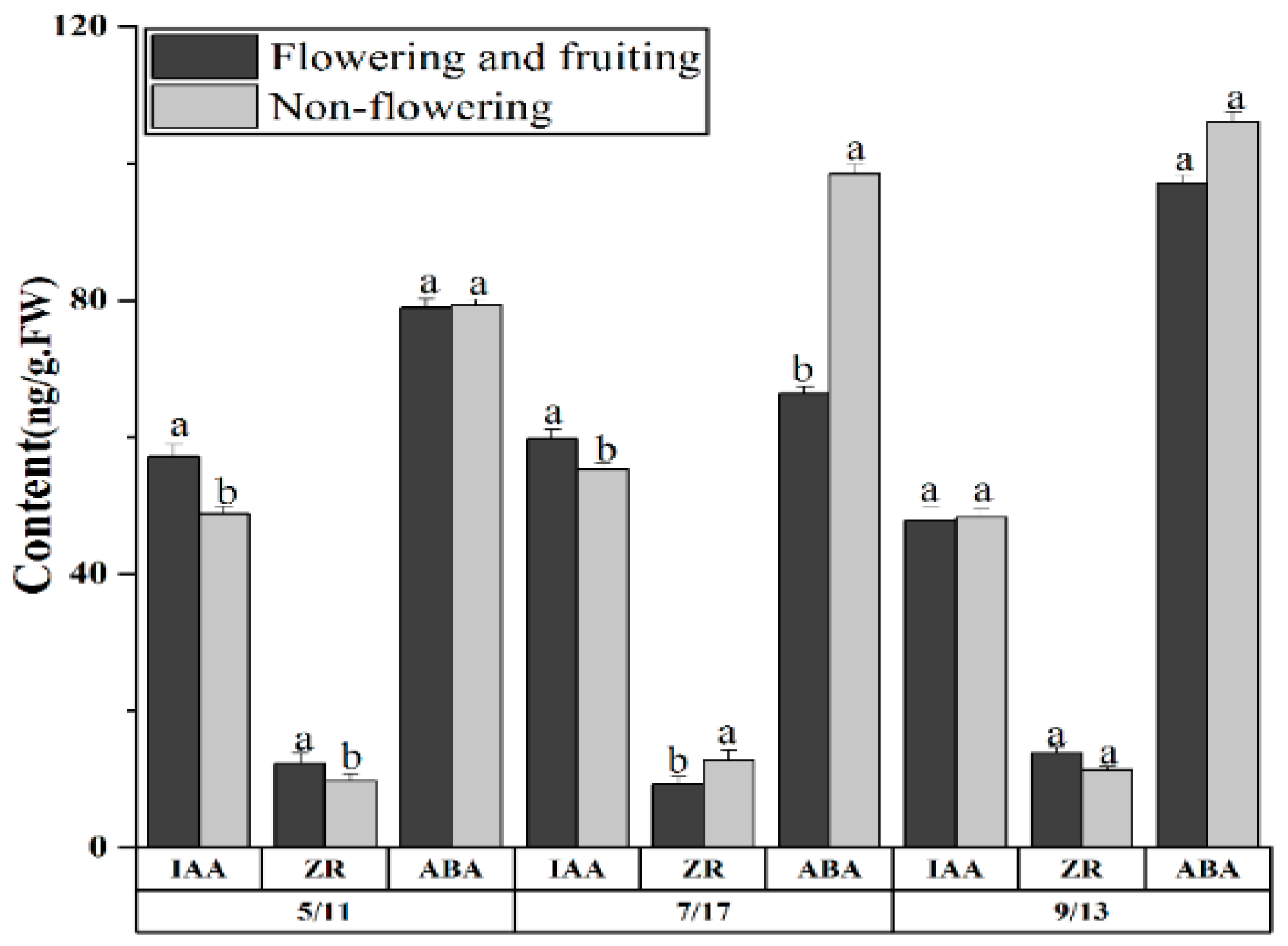
2.3. Differences of the Endogenous Hormones between Fruiting and Nonflowering Plants at Different Stages
2.4. Correlation between the Endogenous Hormones and Flower and Fruit Falling Rates
2.5. Correlations of the Endogenous Hormone with the Number of Empty Seeds, Number of Shrunken Seeds, and TKW
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Experimental Design
4.3. Determination of the Endogenous Hormone Content
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China, Volume I; China Medical Science Press: Beijing, China, 2015; pp. 86–87. (In Chinese) [Google Scholar]
- Vaya, J.; Belinky, P.A.; Aviram, M. Antioxidant constituents from licorice roots: Isolation, structure elucidation and antioxidative capacity toward LDL oxidation. Free Radic. Biol. Med. 1997, 23, 302–313. [Google Scholar] [CrossRef]
- Wang, Q.E.; Ren, H.; Cao, X.L. Research and Utilization Statue of Licorice. Chin. Agric. Sci. Bull. 2011, 27, 290–295. [Google Scholar]
- Seki, H.; Sawai, S.; Ohyama, K. Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin. Plant Cell 2011, 23, 4112–4123. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wang, W.Q.; Liu, Y. Research progress in the biological characteristic and resource cultivation of Glycyrrhiza uralensis Fisch. World For. Res. 2003, 16, 28–32. [Google Scholar]
- Meijón, M.; Jesús Cañal, M.; Valledor, L.; Rodríguez, R.; Feito, I. Epigenetic and physiological effects of gibberellin inhibitors and chemical pruners on the floral transition of azalea. Physiol. Plant. 2011, 141, 276–288. [Google Scholar] [CrossRef]
- Domagalska, M.A.; Sarnowska, E.; Nagy, F.; Davis, S.J. Genetic analyses of interactions among gibberellin, abscisic acid, and brassinosteroids in the control of flowering time in Arabidopsis thaliana. PLoS ONE 2010, 5, e14012. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Oliveros, R.; Guevara-Olvera, L.; Beltrán, J.P.; Gómez-Mena, C.G.A. Developmental landmarks during floral ontogeny of jalapeño chili pepper (Capsicum annuum L.) and the effect of gibberellin on ovary growth. Plant Reprod. 2017, 30, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Popa, C. Ethylene measurements from sweet fruits flowers using photoacoustic spectroscopy. Molecules 2019, 24, 1144. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Trivellini, A.; Fatma, M.; Masood, A.; Francini, A.; Iqbal, N.; Ferrante, A.; Khan, N.A. Role of ethylene in responses of plants to nitrogen availability. Front. Plant Sci. 2015, 6, 927. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Chen, Y.; Gao, M.; Zhao, Y.; Xu, Z.; Cao, P.; Zhang, Q.; Jiao, Y.; Li, H.; Wu, L.; et al. Transcriptome analysis of Litsea cubeba floral buds reveals the role of hormones and transcription factors in the differentiation process. G3 2018, 8, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.W.; Xie, W.H.; Ma, K. Correlation between the endogenous hormones contents and flower bud differentiation stage of Ficus carica L. Acta Bot. Boreali Occident. Sin. 2007, 27, 1399–1404. [Google Scholar]
- Zhang, D.; Ren, L.; Yue, J.H.; Wang, L.; Zhuo, L.H.; Shen, X.H. GA 4 and IAA were involved in the morphogenesis and development of flowers in Agapanthus praecox ssp. orientalis. J. Plant Physiol. 2014, 11, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Bartrina, I.; Jensen, H.; Novák, O.; Strnad, M.; Werner, T.; Schmülling, T. Gain-of-function mutants of the cytokinin receptors AHK2 and AHK3 regulate plant organ size, flowering time and plant longevity. Plant Physiol. 2017, 173, 1783–1797. [Google Scholar] [CrossRef]
- Li, D.; Guo, Z.; Liu, C.; Li, J.; Xu, W.; Chen, Y. Quantification of near-attomole gibberellins in floral organs dissected from a single Arabidopsis thaliana flower. Plant J. 2017, 91, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, M.A.; Weigel, D. Independent regulation of flowering by phytochrome b and gibberellins in Arabidopsis. Plant Physiol. 1999, 120, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.Q.; Wang, L.H. Quantitative variation of the endogenous hormones in Rosa rugose leaf during flower bud differentiation. Plant Res. 2006, 26, 206–210. [Google Scholar]
- Feng, H.; Wang, J.H.; Wang, M.L. The role of hormones in flowering and fruiting of plants. Beijing Gard. 2006, 77, 28–30. [Google Scholar]
- Mutasa-Gottgens, E.; Hedden, P. Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 2009, 60, 1979–1989. [Google Scholar] [CrossRef]
- Li, T.H.; Huang, W.D.; Meng, S.Q. Study on the mechanism of apple flower bud incubation. J. Plant Physiol. 1996, 22, 251–257. [Google Scholar]
- Liang, W.Y.; Liang, L.F.; Ji, Z.L.; Li, P.W. Content dynamics of the endogenous gibberellin and indole acetic acid during flower bud differentiation of Litchi. Acta Hortic. Sin. 1987, 14, 145–152. [Google Scholar]
- Yamaguchi, N.; Winter, C.M.; Wu, M.F. Gibberellin Acts Positively Then Negatively to Control Onset of Flower Formation in Arabidopsis. Science 2014, 344, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; McCourt, P. Cross-talk in plant hormone signalling: What Arabidopsis mutants are telling us. Ann. Bot. 2003, 91, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.Y.; Zhang, J.C.; Wei, L.H. Studies on the changes of the endogenous hormones in the differentiation period of flower bud in apple trees. Plant Sci. 2000, 4, 244–248. [Google Scholar]
- Wang, L.; Tang, G.G.; Liu, T. Variation of the endogenous hormone and nucleicacid content during flower bud differentiation in Lycorisradiata. J. Nanjing For. Univ. Nat. Sci. Ed. 2008, 32, 67–70. [Google Scholar]
- Fang, S.; Gao, K.; Hu, W.; Snider, J.L.; Wang, S.; Chen, B.; Zhou, Z. Chemical priming of seed alters cotton floral bud differentiation by inducing changes in hormones, metabolites and gene expression. Plant Physiol. Biochem. 2018, 130, 633–640. [Google Scholar] [CrossRef]
- Huang, H.N.; He, J.H.; Zhao, X.Q.; Chen, H.L.; Chen, Y.Y. Research progress oil the relationship between the change of the endogenous hormones and embryo abortion. Anhui Agric. Sci. 2013, 41, 12534–12536. [Google Scholar]
- Yin, X.; He, D.; Gupta, R.; Yang, P. Physiological and proteomic analyses on artificially aged Brassica napus seed. Front. Plant Sci. 2015, 6, 112. [Google Scholar] [CrossRef]
- Liu, Y.; Han, J.; Liu, D.; Gu, D.; Wang, Y.; Liao, Y.; Wen, X. Effect of plastic film mulching on the grain filling and hormonal changes of maize under different irrigation conditions. PLoS ONE 2015, 10, e122791. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Liu, K.; Wang, Z.; Liu, L. Abscisic acid and ethylene interact in wheat grains in response to soil drying during grain filling. N. Phytol. 2006, 171, 293–303. [Google Scholar] [CrossRef]
- Liu, Y.; Han, J.; Wen, X.; Wu, W.; Guo, Q.; Zeng, A.; Liao, Y. The effect of plastic-covered ridge and furrow planting on the grain filling and hormonal changes of winter wheat. J. Integr. Agric. 2013, 12, 1771–1782. [Google Scholar] [CrossRef]
- Sehra, B.; Franks, R.G. Auxin and cytokinin act during gynoecial patterning and the development of ovules from the meristematic medial Auxin and cytokinin act during gynoecial patterning and the development of ovules from the meristematic medial domain. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tian, M.; Liu, F.; Wang, C.; Zhang, Y. Hormonal and morphological changes during seed development of Cypripedium japonicum. Protoplasma 2017, 254, 2315–2322. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.F.; Liu, M.J. Change of the endogenous hormone in cultivars of chinese jujube with different type of embryo abortion. Acta Hortic. Sin. 2004, 31, 800–802. [Google Scholar]
- Zhang, Y.S.; Xiang, X.; Huang, S.Z.; Fu, J.R. Changes of the endogenous hormones and protein content during Litchi embryo abortion. J. Plant Physiol. Mol. Biol. 2003, 29, 233–238. [Google Scholar]
- Gao, Q.Y.; Yan, C.; Zhang, B.X.; Wang, Z.W.; Wang, X.J.; Zhang, Y. Changes of Endogenous Hormone Content during the Growth of Raspberry Leaves. Hubei Agric. Sci. 2015, 54, 6280–6283. [Google Scholar]
- Ja, N.; Tian, H.; Peng, Z.J.; Wang, Y.; Wu, T.; Zhang, X.Z. Changes in IAA and ABA levels and PpPINs genes expression during the bud dormancy induction in peach. Chin. Agric. Sci. Bull. 2015, 31, 100–106. [Google Scholar]
- Wang, Y.F.; Cui, Z.H.; Ruan, Y.H.; Ma, X.L.; Guan, Y.X.; Zhang, L.J. Changes in endogenous hormone of IAA, GA, ZR and ABA in kernels during grain-filling stage in different types of spring maize (Zea mays L.). Plant Physiol. Commun. 2006, 42, 225–228. [Google Scholar]
- Li, B.W.; Huang, Y.H.; Zheng, H.F.; Zeng, Z.; Liu, X.L. Study on the change of endogenous hormone content in the process of tea annual growth. Fujian Tea 2003, 2–3. [Google Scholar]
- Wang, W.J.; Gao, S.J. The relationship between the changes of the contents of endogenous ga3, aba and source2sink of different kernel wheat varieties. Plant Physiol. Commun. 2002, 38, 324–326. [Google Scholar]
- Li, G.; Zhao, J.; Qin, B.; Yin, Y.; An, W.; Mu, Z.; Cao, Y. ABA mediates development-dependent anthocyanin biosynthesis and fruit coloration in Lycium plants. BMC Plant Biol. 2019, 19, 317. [Google Scholar] [CrossRef]
- Soto, A.; Ruiz, K.B.; Ravaglia, D.; Costa, G.; Torrigiani, P. ABA may promote or delay peach fruit ripening through modulation of ripening- and hormone-related gene expression depending on the developmental stage. Plant Physiol. Bioch. 2013, 64, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Harshavardhan, V.T.; Govind, G.; Seiler, C.; Kohli, A. Contrapuntal role of ABA: Does it mediate stress tolerance or plant growth retardation under long-term drought stress? Gene 2012, 506, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Razem, F.A.; El-Kereamy, A.; Abrams, S.R.; Hill, R.D. The RNA-binding protein FCA is an abscisic acid receptor. Nature 2008, 456, 824. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.Z.; Li, W.J.; Ren, G.H.; Chen, Y.Z. Changes of IAA content and activities of peroxidase during cotton floral bud initiation. Cotton Sci. 1999, 11, 303–305. [Google Scholar]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef]
- Zhang, G.H.; Zhang, Y.J.; Cong, R.C. Advances on gibberellins mechanism. Acta Bot. Boreali Occident. Sin. 2009, 29, 412–419. [Google Scholar]
- Luo, P.Y.; Shi, J.K.; Zhang, W.P. Changes of the endogenous hormones, carbohydrates and mineral nutrition during female flower bud differentiation of Ginkgo biloba. J. Zhejiang For. Colleg. 2006, 23, 532–537. [Google Scholar]
- Yang, Q.; Fu, Y.; Liu, Q.X. Dynamic changes of the endogenous hormone content and seed abortion in Loqua. China Fruits 2015, 4, 19–23. [Google Scholar]
- Reed, A.J.; Singletary, G.W. Roles of carbohydrate supply and phytohormones in maize kernel abortion. Plant Physiol. 1989, 91, 986–992. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, Z.; Zhou, Q.; Chen, J.; Xu, G.; Gu, J.; Liu, L.; Wang, Z.; Yang, J.; Zhang, H. Grain filling characteristics and their relations with endogenous hormones in large- and small-grain mutants of rice. PLoS ONE 2016, 11, e165321. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Fan, S.; Du, L.; Shen, Y.; Xing, L.; Li, Y.; Ma, J.; Han, M. Effect of exogenous GA 3 and its inhibitor paclobutrazol on floral formation, endogenous hormones, and flowering-associated genes in ‘Fuji’ apple ( Malus domestica Borkh.). Plant Physiol. Bioch. 2016, 107, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Irshad Ahmad, M.K.S.A. Seed filling in maize and hormones crosstalk regulated by exogenous application of uniconazole in semiarid regions. Environ. Sci. Pollut. Res. Int. 2018, 25, 33225–33239. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fang, Y.; Huang, M.; Jin, Y.; Sun, J.; Tao, X.; Zhang, G.; He, K.; Zhao, Y.; Zhao, H. Uniconazole-induced starch accumulation in the bioenergy crop duckweed (Landoltia punctata) II: Transcriptome alterations of pathways involved in carbohydrate metabolism and endogenous hormone crosstalk. Biotechnol. Biofuels 2015, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing Qiao, Z.L.Y.L. Effect of abscisic acid on accumulation of five active components in root of Glycyrrhiza uralensis. Molecules 2017, 22, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the Glycyrrhiza uralensis leaves are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pearson Coefficient | Date | ABA | IAA |
|---|---|---|---|
| Rates of falling flowers and fruit | 5/21 | −0.231 | −0.825 ** |
| 7/17 | 0.805 ** | −0.759 * |
| Pearson Coefficient | Date | IAA | ZR | ZR:GA | IAA:GA | IAA:ABA |
|---|---|---|---|---|---|---|
| Rate of empty seeds | 7/17 | −0.786 * | −0.105 | −0.198 | −0.701 * | −0.782 * |
| Rate of shrunken seeds | 7/17 | −0.631 | −0.415 | −0.569 | −0.667 * | −0.789 * |
| TKW | 7/17 | 0.527 | 0.773 * | 0.839 ** | 0.482 | 0.675 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, B.; Hou, J.; Cui, J.; He, C.; Li, W.; Chen, X.; Li, M.; Wang, W. The Effects of Endogenous Hormones on the Flowering and Fruiting of Glycyrrhiza uralensis. Plants 2019, 8, 519. https://doi.org/10.3390/plants8110519
Yan B, Hou J, Cui J, He C, Li W, Chen X, Li M, Wang W. The Effects of Endogenous Hormones on the Flowering and Fruiting of Glycyrrhiza uralensis. Plants. 2019; 8(11):519. https://doi.org/10.3390/plants8110519
Chicago/Turabian StyleYan, Binbin, Junling Hou, Jie Cui, Chao He, Wenbin Li, Xiaoyu Chen, Min Li, and Wenquan Wang. 2019. "The Effects of Endogenous Hormones on the Flowering and Fruiting of Glycyrrhiza uralensis" Plants 8, no. 11: 519. https://doi.org/10.3390/plants8110519