The Development and Characterization of Near-Isogenic and Pyramided Lines Carrying Resistance Genes to Brown Planthopper with the Genetic Background of Japonica Rice (Oryza sativa L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
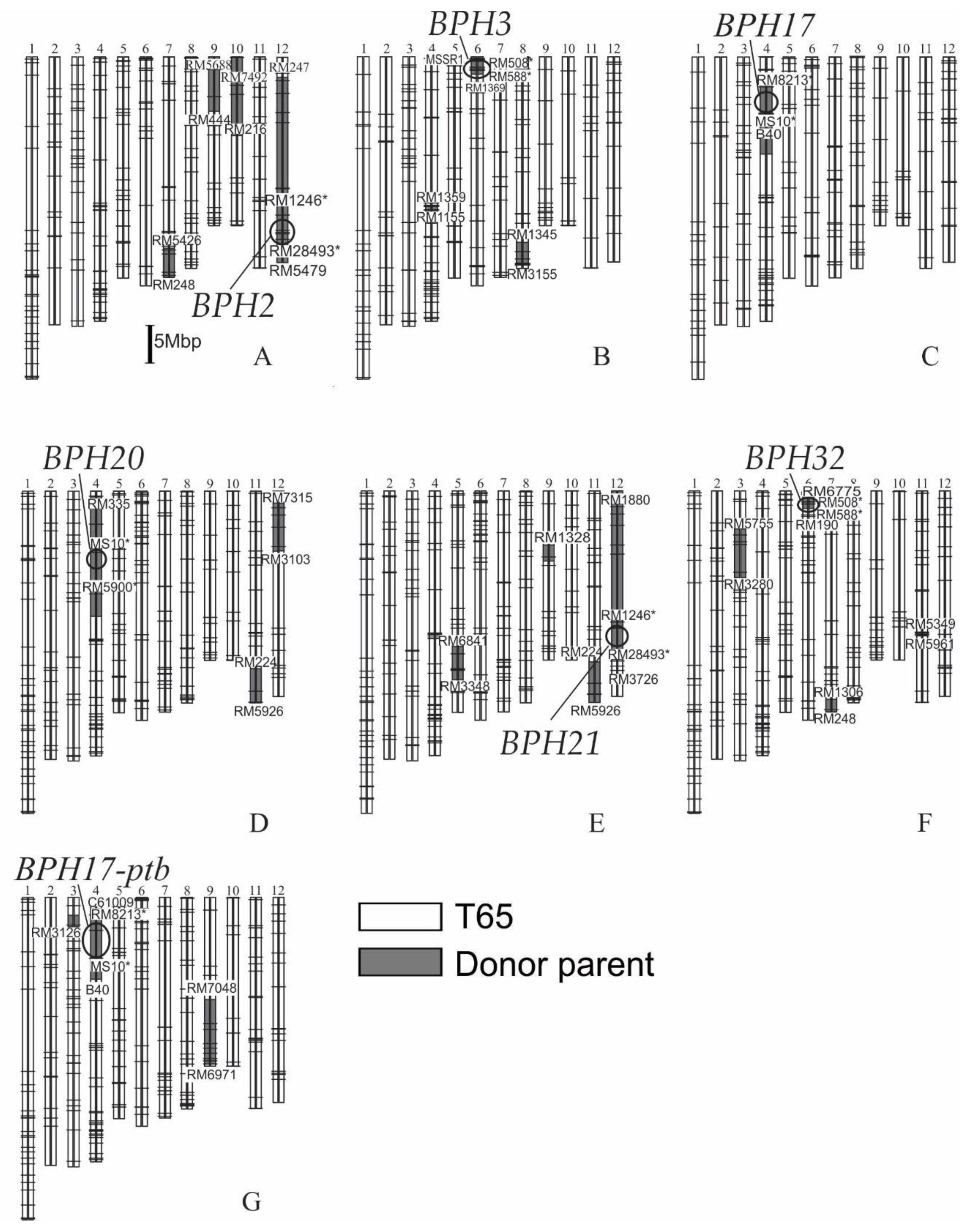
2.1. Development of Seven NILs for BPH Resistance
2.2. Development of 15 PYLs Carrying Two or Three BPH Resistance Genes
2.3. Comparison of Resistance Levels against Hadano-66 by Modified Seedbox Screening Test ( MSST)
2.4. Comparison of Adult Mortality (ADM) of the Hadano-66 Population to the NILs and PYLs
2.5. Comparison of ADM Rates for the Koshi-2013 Population on the NILs and PYLs
2.6. Agronomic Characteristics of the NILs and PYLs
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. The Development of PYLs with BPH Resistance Genes
4.3. The MAS for BPH Resistance Genes
4.4. The Genetic Background Survey of the NILs
4.5. The BPH Populations and the Characterization of BPH Resistance
4.6. Antibiosis Tests
4.7. Characterization of NILs and PYLs for Agronomic Traits
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Wei, J.; Jia, D.; Mao, Q.; Zhang, X.; Chen, Q.; Wu, W.; Chen, H.; Wei, T. Complex interactions between insect-borne rice viruses and their vectors. Curr. Opin. Virol. 2018, 33, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice resistance to planthoppers and leafhoppers. CRC Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Horgan, F.G. Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers. Crop Prot. 2018, 110, 21–33. [Google Scholar] [CrossRef]
- Horgan, F.G. Insect herbivores of rice: Their natural regulation and ecologically based management. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer: Cham, Switzerland, 2017; pp. 279–302. [Google Scholar]
- Zhao, Y.; Huang, J.; Wang, Z.; Jing, S.; Wang, Y.; Ouyang, Y.; Cai, B.; Xin, X.-F.; Liu, X.; Zhang, C.; et al. Allelic diversity in an NLR gene BPH9 enables rice to combat planthopper variation. Proc. Natl. Acad. Sci. USA 2016, 113, 12850–12855. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B.; et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2014, 33, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Kim, S.R.; Kim, Y.H.; Suh, J.P.; Park, H.M.; Sreenivasulu, N.; Misra, G.; Kim, S.M.; Hechanova, S.L.; Kim, H.; et al. Map-based cloning and characterization of the BPH18 gene from wild rice conferring resistance to brown planthopper (BPH) insect pest. Sci. Rep. 2016, 6, 34376. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Hattori, M.; Yoshioka, H.; Yoshioka, M.; Takahashi, A.; Wu, J.; Sentoku, N.; Yasui, H. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci. Rep. 2014, 4, 5872. [Google Scholar] [PubMed]
- Wang, Y.; Cao, L.; Zhang, Y.; Cao, C.; Liu, F.; Huang, F.; Qiu, Y.; Li, R.; Luo, X. Map-based cloning and characterization of BPH29, a B3 domain-containing recessive gene conferring brown planthopper resistance in rice. J. Exp. Bot. 2015, 66, 6035–6045. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J.; Su, X.; Luo, H.; Ren, G. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 37645. [Google Scholar] [CrossRef] [PubMed]
- Kamolsukyeunyong, W.; Ruengphayak, S.; Chumwong, P.; Kusumawati, L.; Chaichoompu, E.; Jamboonsri, W.; Saensuk, C.; Phoonsiri, K.; Toojinda, T.; Vanavichit, A. Identification of spontaneous mutation for broad-spectrum brown planthopper resistance in a large, long-term fast neutron mutagenized rice population. Rice 2019, 12, 16. [Google Scholar] [PubMed]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. Development and characterization of japonica rice lines carrying the brown planthopper-resistance genes BPH12 and BPH6. Theor. Appl. Genet. 2012, 124, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; You, A.; Yang, Z.; Zhang, F.; He, R.; Zhu, L.; He, G. High-resolution genetic mapping at the Bph15 locus for brown planthopper resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2004, 110, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Su, C.; Wang, C.; Zhai, H.; Wan, J. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed. Sci. 2005, 55, 391–396. [Google Scholar] [CrossRef]
- Rahman, M.L.; Jiang, W.; Chu, S.H.; Qiao, Y.; Ham, T.H.; Woo, M.O.; Lee, J.; Khanam, M.S.; Chin, J.H.; Jeung, J.U.; et al. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta. Theor. Appl. Genet. 2009, 119, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Xiao, C.; Cheng, M.; Gao, G.; Zhang, Q.; He, Y. Fine mapping and pyramiding of brown planthopper resistance genes QBph3 and Qbph4 in an introgression line from wild rice O. officinalis. Mol. Breed. 2015, 35, 3. [Google Scholar] [CrossRef]
- Hu, J.; Xiao, C.; Cheng, M.; Gao, G.; Zhang, Q.; He, Y. A new finely mapped Oryza australiensis-derived QTL in rice confers resistance to brown planthopper. Gene 2015, 561, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.K.; Panda, R.S.; Mohapatra, S.L.; Nanda, A.; Behera, L.; Jena, M.; Sahu, R.K.; Sahu, S.C.; Mohapatra, T. Identification of novel quantitative trait loci associated with brown planthopper resistance in the rice landrace Salkathi. Euphytica 2017, 213, 38. [Google Scholar] [CrossRef]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds. Theor. Appl. Genet. 2010, 121, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, H.; Kaji, R.; Angeles, E.R.; Ogawa, T.; Brar, D.S.; Khush, G.S. RFLP analysis of a new gene for resistance to brown planthopper derived from O. officinalis on rice chromosome 4. Breed. Res. 1999, 48 (Suppl. 1), 82. (In Japanese) [Google Scholar]
- Li, R.; Li, L.; Wei, S.; Wei, Y.; Chen, Y.; Bai, D.; Yang, L.; Huang, F.; Lu, W.; Zhang, X.; et al. The evaluation and utilization of new genes for brown planthopper resistance in common wild rice (Oryza rufipogon Griff.). Mol. Entomol. 2010, 1, 1. [Google Scholar]
- Huang, D.; Qiu, Y.; Zhang, Y.; Huang, F.; Meng, J.; Wei, S.; Li, R.; Chen, B. Fine mapping and characterization of BPH27, a brown planthopper resistance gene from wild rice (Oryza rufipogon Griff.). Theor. Appl. Genet. 2013, 126, 219–229. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, Y.; Liu, Y.; Jiang, L.; Wu, H.; Kang, H.; Liu, S.; Chen, L.; Liu, X.; Cheng, X.; et al. High-resolution mapping of brown planthopper (BPH) resistance gene Bph27(t) in rice (Oryza sativa L.). Mol. Breed. 2013, 31, 549–557. [Google Scholar] [CrossRef]
- Lakshminarayana, A.; Khush, G.S. New genes for resistance to the brown planthopper in rice. Crop Sci. 1977, 17, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 2007, 19, 35–44. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Murata, K.; Ishii, T.; Takumi, S.; Mori, N.; Nakamura, C. Assignment of a brown planthopper (Nilaparvata lugens Stål) resistance gene bph4 to the rice chromosome 6. Breed. Sci. 2001, 51, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 2012, 124, 495–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Liu, Y.; Jiang, L.; Su, C.; Wang, C.; Zhai, H.; Wan, J. Identification of quantitative trait loci associated with resistance to brown planthopper in the indica rice cultivar Col.5 Thailand. Hereditas 2007, 144, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Athwal, D.S.; Pathak, M.D.; Bacalangco, E.H.; Pura, C.D. Genetics of resistance to brown planthoppers and green leafhoppers in Oryza sativa L. Crop Sci. 1971, 11, 747–750. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.H.; Ahn, S.N.; Choi, H.C.; Hahn, T.R.; Moon, H.P. Identification of a RAPD marker linked to a brown planthopper resistance gene in rice. Euphytica 1999, 107, 23–28. [Google Scholar] [CrossRef]
- Sun, L.H.; Wang, C.M.; Su, C.C.; Liu, Y.Q.; Zhai, H.Q.; Wan, J.M. Mapping and marker-assisted selection of a brown planthopper resistance gene bph2 in rice (Oryza sativa L.). Acta Genet. Sin. 2006, 33, 717–723. [Google Scholar] [CrossRef]
- Kabir, M.A.; Khush, G.S. Genetic analysis of resistance to brown planthopper in rice (Oryza sativa L.). Plant Breed. 1988, 100, 54–58. [Google Scholar]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. Fine mapping of the rice brown planthopper resistance gene BPH7 and characterization of its resistance in the 93-11 background. Euphytica 2014, 198, 369–379. [Google Scholar] [CrossRef]
- Murata, K.; Fujiwara, M.; Murai, H.; Takumi, S.; Mori, N.; Nakamura, C. Bph9, a dominant brown planthopper resistance gene, is located on the long arm of rice chromosome 12. Rice Genet. Newslett. 2000, 17, 84–86. [Google Scholar]
- Ishii, T.; Brar, D.S.; Multani, D.S.; Khush, G.S. Molecular tagging of genes for brown planthopper resistance and earliness introgressed from Oryza australiensis into cultivated rice, O. sativa. Genome 1994, 37, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Jena, K.K.; Jeung, J.U.; Lee, J.H.; Choi, H.C.; Brar, D.S. High-resolution mapping of a new brown planthopper (BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 112, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Jairin, J.; Teangdeerith, S.; Leelagud, P.; Phengrat, K.; Vanavichit, A.; Toojinda, T. Detection of brown planthopper resistance genes from different rice mapping populations in the same genomic location. ScienceAsia 2007, 33, 347–352. [Google Scholar] [CrossRef]
- Sidhu, G.S.; Khush, G.S. Genetic analysis of brown planthopper resistance in twenty varieties of rice, Oryza sativa L. Theor. Appl. Genet. 1978, 53, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Angeles, E.R.; Khush, G.S.; Heinrichs, E.A. Inheritance of resistance to planthoppers and leafhoppers in rice. In Proceedings of the International Rice Genetics Symposium, Manila, Philippines, 27–31 May 1985; International Rice Research Institute: Manila, Philippines, 1986; pp. 537–549. [Google Scholar]
- Saxena, R.C.; Barrion, A.A. Biotypes of the brown planthopper Nilaparvata lugens (Stål) and strategies in deployment of host plant resistance. Insect Sci. Appl. 1985, 6, 271–289. [Google Scholar] [CrossRef]
- Ketipearachchi, Y.; Kaneda, C.; Nakamura, C. Adaptation of the brown planthopper (BPH), Nilaparvata lugens (Stål) (Homoptera: Delphacidae), to BPH resistant rice cultivars carrying bph8 or Bph9. Appl. Entomol. Zool. 1998, 33, 497–505. [Google Scholar] [CrossRef]
- Pathak, M.D.; Khush, G.S. Studies of varietal resistance in rice to the brown planthopper at the International Rice Research Institute. In Brown Planthopper—Threat to Rice Production in Asia; International Rice Research Institute: Los Baños, Philippines, 1979; pp. 285–301. [Google Scholar]
- Horgan, F.G.; Ramal, A.F.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Iswanto, E.H.; Van Chien, H.; Phyu, M.H.; Bernal, C.C.; et al. Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia against resistant rice varieties. Crop Prot. 2015, 78, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Claridge, M.F.; Hollander, J.D. Virulence to rice cultivars and selection for virulence in populations of the brown planthopper Nilaparvata lugens. Entomol. Exp. Appl. 1982, 32, 213–221. [Google Scholar] [CrossRef]
- Alam, S.N.; Cohen, M.B. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a doubled-haploid rice population. Theor. Appl. Genet. 1998, 97, 1370–1379. [Google Scholar] [CrossRef]
- Peñalver Cruz, A.; Arida, A.; Heong, K.L.; Horgan, F.G. Aspects of brown planthopper adaptation to resistant rice varieties with the Bph3 gene. Entomol. Exp. Appl. 2011, 141, 245–257. [Google Scholar] [CrossRef]
- Ferrater, J.B.; Naredo, A.I.; Almazan, M.L.P.; de Jong, P.W.; Dicke, M.; Horgan, F.G. Varied responses by yeast-like symbionts during virulence adaptation in a monophagous phloem-feeding insect. Arthropod-Plant Interact. 2015, 9, 215–224. [Google Scholar] [CrossRef]
- Horgan, F.G.; Ferrater, J.B. Benefits and potential trade-offs associated with yeast-like symbionts during virulence adaptation in a phloem-feeding planthopper. Entomol. Exp. Appl. 2017, 163, 112–125. [Google Scholar] [CrossRef]
- Myint, K.K.M.; Yasui, H.; Takagi, M.; Matsumura, M. Virulence of long-term laboratory populations of the brown planthopper, Nilaparvata lugens (Stål), and whitebacked planthopper, Sogatella furcifera (Horváth) (Homoptera: Delphacidae), on rice differential varieties. Appl. Entomol. Zool. 2009, 44, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Guo, J.; Jing, S.; Tang, M.; Zhu, L.; He, G. Identification of antibiosis and tolerance in rice varieties carrying brown planthopper resistance genes. Entomol. Exp. Appl. 2011, 141, 224–231. [Google Scholar] [CrossRef]
- Ali, M.P.; Alghamdi, S.; Begum, M.; Anwar Uddin, A.B.M.; Alam, M.Z.; Huang, D. Screening of rice genotypes for resistance to the brown planthopper, Nilaparvata lugens Stål. Cereal Res. Commun. 2012, 40, 502–508. [Google Scholar] [CrossRef]
- Yara, A.; Phi, C.N.; Matsumura, M.; Yoshimura, A.; Yasui, H. Development of near-isogenic lines for BPH25(t) and BPH26(t), which confer resistance to the brown planthopper, Nilaparvata lugens (Stål.) in indica rice ‘ADR52’. Breed. Sci. 2010, 60, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, L.; Liu, Y.; Dai, H.; He, J.; Kang, H.; Pan, G.; Huang, J.; Qiu, Z.; Wang, Q.; et al. Marker assisted pyramiding of two brown planthopper resistance genes, Bph3 and Bph27 (t), into elite rice Cultivars. Rice 2016, 9, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Li, J.; Yu, J.; Meng, Q.; Deng, X.; Yi, Z.; Xiao, G. Improvement of bacterial blight and brown planthopper resistance in an elite restorer line Huazhan of Oryza. F. Crop. Res. 2016, 186, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Jena, K.K.; Hechanova, S.L.; Verdeprado, H.; Prahalada, G.D.; Kim, S.R. Development of 25 near-isogenic lines (NILs) with ten BPH resistance genes in rice (Oryza sativa L.): Production, resistance spectrum, and molecular analysis. Theor. Appl. Genet. 2017, 130, 2345–2360. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shi, S.; Guo, Q.; Nie, L.; Du, B.; Chen, R.; Zhu, L.; He, G. High-resolution mapping of a gene conferring strong antibiosis to brown planthopper and developing resistant near-isogenic lines in 9311 background. Mol. Breed. 2018, 38, 107. [Google Scholar] [CrossRef]
- Horgan, F.G.; Almazan, M.L.P.; Vu, Q.; Ramal, A.F.; Bernal, C.C.; Yasui, H.; Fujita, D. Unanticipated benefits and potential ecological costs associated with pyramiding leafhopper resistance loci in rice. Crop Prot. 2019, 115, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, X.; Wu, C.; Yang, C.; Hua, H.; Gao, G.; Xiao, J.; He, Y. Pyramiding and evaluation of the brown planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Breed. 2012, 29, 61–69. [Google Scholar] [CrossRef]
- Fujita, D.; Myint, K.K.M.; Matsumura, M.; Yasui, H. The genetics of host-plant resistance to rice planthopper and leafhopper. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hard, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 389–399. [Google Scholar]
- McCouch, S.R.; Teytelman, L.; Xu, Y.; Lobos, K.; Clare, K.; Walton, M.; Fu, B.; Maghirang, R.; Li, Z.; Xing, Y.; et al. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 2002, 9, 199–207. [Google Scholar] [CrossRef] [PubMed]
- International Rice Genome Sequencing Project (IRGSP). The map-based sequence of the rice genome. Nature 2005, 436, 793–800. [Google Scholar]
- Temnykh, S.; DeClerck, G.; Lukashova, A.; Lipovich, L.; Cartinhour, S.; McCouch, S. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): Frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 2001, 11, 1441–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temnykh, S.; Park, W.D.; Ayres, N.; Cartinhour, S.; Hauck, N.; Lipovich, L.; Cho, Y.G.; Ishii, T.; McCouch, S.R. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor. Appl. Genet. 2000, 100, 697–712. [Google Scholar] [CrossRef]
- The Rice Annotation Project (RAP). Available online: https://rapdb.dna.affrc.go.jp/index.html (accessed on 8 November 2019).
- Khush, G.S.; Virk, P.S. IR Varieties and Their Impact; International Rice Research Institute: Los Baños, Philippines, 2005. [Google Scholar]
- Xiao, C.; Hu, J.; Ao, Y.T.; Cheng, M.X.; Gao, G.J.; Zhang, Q.L.; He, G.C.; He, Y.Q. Development and evaluation of near-isogenic lines for brown planthopper resistance in rice cv. 9311. Sci. Rep. 2016, 6, 38159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jairin, J.; Teangdeerith, S.; Leelagud, P.; Phengrat, K.; Vanavichit, A.; Toojinda, T. Physical mapping of Bph3, a brown planthopper resistance locus in rice. Maejo Int. J. Sci. Technol. 2007, 1, 166–177. [Google Scholar]
- Sharma, P.N.; Torii, A.; Takumi, S.; Mori, N.; Nakamura, C. Marker-assisted pyramiding of brown planthopper (Nilaparvata lugens Stål) resistance genes Bph1 and Bph2 on rice chromosome 12. Hereditas 2004, 140, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Myint, K.K.M.; Matsumura, M.; Takagi, M.; Yasui, H. Demographic parameters of long-term laboratory strains of the brown planthopper, Nilaparvata lugens Stål, (Homoptera: Delphacidae) on resistance genes, bph20(t) and Bph21(t) in rice. J. Fac. Agric. Kyushu Univ. 2009, 54, 159–164. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Young, N.D.; Tanksley, S.D. Restriction fragment length polymorphism maps and the concept of graphical genotypes. Theor. Appl. Genet. 1989, 77, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Naeemullah, M.; Sharma, P.N.; Tufail, M.; Mori, N.; Matsumura, M.; Takeda, M.; Nakamura, C. Characterization of brown planthopper strains based on their differential responses to introgressed resistance genes and on mitochondrial DNA polymorphism. Appl. Entomol. Zool. 2009, 44, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Standard Evaluation System for Rice (SES), 5th ed.; International Rice Research Institute: Manila, Philippines, 2014.




{kind=link}
{kind=link}
{kind=link}
| Marker | Resistance Gene Tagged | Chromosome | Forward Primer Sequence (5′→3′) | Reverse Primer Sequence (5′→3′) | Physical Position (Mbp) * | Predicted Size (bp) ** |
|---|---|---|---|---|---|---|
| RM1246 a | BPH2, BPH21 | 12 | GGCTCACCTCGTTCTCGATCC | CATAAATAAATAGGGCGCCACACC | 19.16 | 195 |
| RM28493 b | BPH2, BPH21 | 12 | ACCGTTAGATGACACAAGCAACG | GGTTAGCAAGACTGGAGGAGACG | 23.28 | 259 |
| RM508 c | BPH3, BPH32 | 6 | AGAAGCCGGTTCATAGTTCATGC | ACCCGTGAACCACAAAGAACG | 0.44 | 158 |
| RM588 c | BPH3, BPH32 | 6 | TCTTGCTGTGCTGTTAGTGTACG | GCAGGACATAAATACTAGGCATGG | 1.61 | 97 |
| RM8213 a | BPH17, BPH17-ptb | 4 | TGTTGGGTGGGTAAAGTAGATGC | CCCAGTGATACAAAGATGAGTTGG | 4.42 | 178 |
| MS10 d | BPH17, BPH17-ptb, BPH20 | 4 | CAATACGAGAAGCCCCTCAC | CTGAAGGAACACGCGGTAGT | 8.08 | 167 |
| RM5900 a | BPH20 | 4 | TTCTACGTTTGACCGTCA | TCTAGGAGCGTTTGTAGGAG | 13.77 | 248 |
| S00310 e | BPH25 | 6 | CAACAAGATGGACGGCAAGG | TTGGAAGAAAAGGCAGGCAC | 0.21 | 215 |
| MSSR1 e | BPH25 | 6 | CTAGCTGCTCTGCTCTGCTG | CGGCAATCTCTCCGAATC | 0.22 | 114 |
| RM309 f | BPH26 | 12 | CACGCACCTTTCTGGCTTTCAGC | AGCAACCTCCGACGGGAGAAGG | 21.52 | 177 |
| RM28438 b | BPH26 | 12 | GTTCGTGAGCCACAACAAATCC | GTTAAATGCTCCACCAAACACACC | 22.59 | 216 |
| InD14 g | BPH26 | 12 | GGCCGAGTAGGATACTCTAGAAA | CTGCGAGAAAGGAGAGGTGG | 22.87 | 387 |
| RM28466 b | BPH26 | 12 | CCGACGAAGAAGACGAGGAGTAGCC | AGGCCGGAGAGCAATCATGTCG | 22.98 | 93 |
| RM28481 b | BPH26 | 12 | GTCAATTAACCATTGCCCATGC | TTCACGTGGGAACTACTCATGC | 23.14 | 242 |
| MSSR2 e | BPH26 | 12 | CATGTCGAAGAGGTTGCAGA | GGTTTCATCCAAGTCCACGA | 25.03 | 265 |
| Entry | Gene (Donor Parent) | Generation | |||||
|---|---|---|---|---|---|---|---|
| BPH2-NIL | BPH2 | (PTB33) | BC4F3 | ||||
| BPH3-NIL | BPH3 | (Rathu Heenati) | BC4F4 | ||||
| BPH17-NIL | BPH17 | (Rathu Heenati) | BC4F4 | ||||
| BPH20-NIL | BPH20 | (IR71033-121-15) | BC4F5 | ||||
| BPH21-NIL | BPH21 | (IR71033-121-15) | BC4F5 | ||||
| BPH32-NIL | BPH32 | (PTB33) | BC4F4 | ||||
| BPH17-ptb-NIL | BPH17-ptb | (PTB33) | BC4F3 | ||||
| BPH2+BPH17-PYL | BPH2 | (PTB33) | BPH17 | (Rathu Heenati) | BC4F3 equivalent | ||
| BPH2+BPH25-PYL | BPH2 | (PTB33) | BPH25 | (ADR52) | BC4F3 equivalent | ||
| BPH2+BPH32-PYL | BPH2 | (PTB33) | BPH32 | (PTB33) | BC4F3 equivalent | ||
| BPH2+BPH17-ptb-PYL | BPH2 | (PTB33) | BPH17-ptb | (PTB33) | BC4F3 equivalent | ||
| BPH3+BPH17-PYL | BPH3 | (Rathu Heenati) | BPH17 | (Rathu Heenati) | BC4F4 equivalent | ||
| BPH17+BPH21-PYL | BPH17 | (Rathu Heenati) | BPH21 | (IR71033-121-15) | BC4F3 equivalent | ||
| BPH20+BPH21-PYL | BPH20 | (IR71033-121-15) | BPH21 | (IR71033-121-15) | BC3F8 equivalent | ||
| BPH20+BPH32-PYL | BPH20 | (IR71033-121-15) | BPH32 | (PTB33) | BC4F3 equivalent | ||
| BPH21+BPH25-PYL | BPH21 | (IR71033-121-15) | BPH25 | (ADR52) | BC4F3 equivalent | ||
| BPH21+BPH17-ptb-PYL | BPH21 | (IR71033-121-15) | BPH17-ptb | (PTB33) | BC4F3 equivalent | ||
| BPH25+BPH17-ptb-PYL | BPH25 | (ADR52) | BPH17-ptb | (PTB33) | BC4F3 equivalent | ||
| BPH32+BPH17-ptb-PYL | BPH32 | (PTB33) | BPH17-ptb | (PTB33) | BC3F8 equivalent | ||
| BPH2+BPH3+BPH17-PYL | BPH2 | (PTB33) | BPH3 | (Rathu Heenati) | BPH17 | (Rathu Heenati) | BC4F3 equivalent |
| BPH2+BPH32+BPH17-ptb-PYL | BPH2 | (PTB33) | BPH32 | (PTB33) | BPH17-ptb | (PTB33) | BC4F3 equivalent |
| BPH20+BPH21+BPH32-PYL | BPH20 | (IR71033-121-15) | BPH21 | (IR71033-121-15) | BPH32 | (PTB33) | BC4F3 equivalent |
| NIL | Donor | No. of SSR Markers | Genome Ratio (%) | Total Physical Distance of Donor Segment (Mbp) * | ||||
|---|---|---|---|---|---|---|---|---|
| T65 | Donor | Total | T65 | Donor | ||||
| bph2-NIL | PTB33 | 183 | 20 | 203 | 85.2–90.9 | 9.1–14.8 | 33.9 | 55.0 |
| Bph3-NIL | Rathu Heenati | 181 | 14 | 195 | 97.0–99.0 | 1.0–3.0 | 3.8 | 11.3 |
| Bph17-NIL | Rathu Heenati | 170 | 3 | 173 | 95.2–99.0 | 1.0–4.8 | 3.8 | 17.6 |
| BPH17-ptb-NIL | PTB33 | 219 | 10 | 229 | 92.4–97.2 | 2.8–7.6 | 10.5 | 28.1 |
| Bph20-NIL | IR71033-121-15 | 224 | 13 | 237 | 90.4–94.4 | 5.6–9.6 | 20.6 | 35.5 |
| Bph21-NIL | IR71033-121-15 | 210 | 19 | 229 | 88.4–92.9 | 7.1–11.6 | 26.4 | 43.1 |
| BPH32-NIL | PTB33 | 220 | 13 | 233 | 95.9–98.1 | 1.9–4.1 | 7.1 | 15.1 |
| Entry | Adult Mortality (%) | |
|---|---|---|
| Hadano-66 | Koshi-2013 | |
| BPH2-NIL | 68.9 ± 28.5 abcd | 4.0 ± 8.9 b |
| BPH3-NIL | 30.0 ± 38.0 def | 0.0 ± 0.0 b |
| BPH17-NIL | 59.0 ± 25.1 abcde | 20.0 ± 14.1 b |
| BPH20-NIL | 24.0 ± 22.7 def | 4.0 ± 8.9 b |
| BPH21-NIL | 36.0 ± 37.5 cdef | 12.0 ± 17.9 b |
| BPH25-NIL | 16.0 ± 15.8 f | 16.0 ± 16.7 b |
| BPH26-NIL | 50.0 ± 41.4 abcdef | 4.0 ± 8.9 b |
| BPH32-NIL | 14.0 ± 16.5 f | 12.0 ± 17.9 b |
| BPH17-ptb-NIL | 22.0 ± 19.9 ef | 20.0 ± 20.0 b |
| BPH2+BPH17-PYL | 75.0 ± 19.5 abcd | 32.0 ± 17.9 b |
| BPH2+BPH25-PYL | 87.5 ± 17.9 ab | 12.0 ± 11.0 b |
| BPH2+BPH32-PYL | 84.0 ± 16.7 abc | 16.0 ± 16.7 b |
| BPH2+BPH17-ptb-PYL | 84.0 ± 16.7 abc | 16.0 ± 16.7 b |
| BPH3+BPH17-PYL | 50.0 ± 32.5 abcdef | 24.0 ± 16.7 b |
| BPH17+BPH21-PYL | 58.3 ± 27.5 abcdef | 16.0 ± 21.9 b |
| BPH20+BPH21-PYL | 33.3 ± 22.2 cdef | 24.0 ± 16.7 b |
| BPH20+BPH32-PYL | 62.5 ± 38.5 abcde | 36.0 ± 21.9 b |
| BPH21+BPH25-PYL | 64.0 ± 26.1 abcde | 24.0 ± 16.7 b |
| BPH21+BPH17-ptb-PYL | 54.2 ± 35.0 abcdef | 8.0 ± 17.9 b |
| BPH25+BPH17-ptb-PYL | 68.0 ± 26.8 abcde | 16.0 ± 16.7 b |
| BPH32+BPH17-ptb-PYL | 62.5 ± 32.9 abcde | 20.0 ± 14.1 b |
| BPH2+BPH3+BPH17-PYL | 96.0 ± 8.9 ab | 36.0 ± 38.5 b |
| BPH2+BPH32+BPH17-ptb-PYL | 95.8 ± 8.9 ab | 20.8 ± 20.1 b |
| BPH20+BPH21+BPH32-PYL | 92.0 ± 11.0 ab | 28.0 ± 26.8 b |
| IR71033-121-15 | 100.0 ± 0.0 a | 44.0 ± 16.7 ab |
| PTB33 | 100.0 ± 0.0 a | 36.0 ± 21.9 b |
| Rathu Heenati | 100.0 ± 0.0 a | 84.0 ± 35.8 a |
| Taichung 65 | 17.6 ± 16.7 f | 5.0 ± 10.0 b |
| Entry | Average of Agronomic Trait (AVE ± SD) | |||||
|---|---|---|---|---|---|---|
| DTH (day) | CL (cm) | PL (cm) | LL (cm) | LW (cm) | PN | |
| BPH2-NIL | 104.0 ± 2.0 | 120.3 ± 3.2 * | 23.8 ± 0.7 | 36.8 ± 4.5 | 1.1 ± 0.0 | 16.6 ± 3.2 |
| BPH3-NIL | 100.8 ± 1.3 | 104.3 ± 4.8 | 18.9 ± 1.0 | 30.2 ± 3.3 | 1.3 ± 0.0 * | 16.2 ± 1.5 |
| BPH17-NIL | 98.6 ± 1.3 | 103.2 ± 7.5 | 23.4 ± 2.2 | 33.8 ± 8.0 | 1.2 ± 0.1 | 15.2 ± 3.1 |
| BPH20-NIL | 100.8 ± 1.6 | 94.2 ± 5.6 | 20.5 ± 2.0 | 29.7 ± 4.5 | 1.1 ± 0.0 | 14.8 ± 4.1 |
| BPH21-NIL | 100.8 ± 1.5 | 104.2 ± 2.9 | 20.9 ± 1.2 | 32.6 ± 3.0 | 1.0 ± 0.1 | 13.8 ± 2.6 |
| BPH25-NIL | 100.4 ± 0.9 | 100.0 ± 1.9 | 18.1 ± 1.9 ** | 26.7 ± 2.8 | 1.1 ± 0.1 | 17.6 ± 1.1 |
| BPH26-NIL | 98.4 ± 0.9 | 102.1 ± 0.9 | 22.0 ± 1.6 | 27.3 ± 3.2 | 1.1 ± 0.0 | 13.4 ± 1.8 |
| BPH32-NIL | 100.2 ± 1.8 | 97.4 ± 1.5 | 20.9 ± 0.9 | 29.3 ± 1.0 | 1.2 ± 0.0 | 15.2 ± 1.6 |
| BPH17-ptb-NIL | 99.0 ± 0.0 | 97.4 ± 2.3 | 20.7 ± 1.0 | 29.6 ± 1.9 | 1.1 ± 0.0 | 14.6 ± 1.7 |
| BPH2+BPH17-PYL | 104.6 ± 1.3 | 115.1 ± 7.9 * | 19.8 ± 1.6 | 27.3 ± 2.6 | 1.2 ± 0.0 | 13.8 ± 2.0 |
| BPH2+BPH25-PYL | 103.0 ± 1.4 | 116.2 ± 1.7 * | 21.0 ± 0.9 | 30.2 ± 4.7 | 1.1 ± 0.1 | 14.2 ± 1.6 |
| BPH2+BPH32-PYL | 104.8 ± 1.3 | 111.4 ± 8.0 * | 19.6 ± 1.6 | 27.8 ± 5.5 | 1.0 ± 0.0 ** | 17.8 ± 3.7 |
| BPH2+BPH17-ptb-PYL | 100.8 ± 0.8 | 110.7 ± 2.3 ** | 23.0 ± 1.1 | 26.9 ± 5.0 | 1.2 ± 0.1 | 15.2 ± 2.7 |
| BPH3+BPH17-PYL | 102.4 ± 1.3 | 103.3 ± 0.5 | 18.8 ± 0.9 | 29.7 ± 3.3 | 1.5 ± 0.1 * | 17.4 ± 2.8 |
| BPH17+BPH21-PYL | 98.0 ± 0.0 | 91.5 ± 1.8 | 21.9 ± 1.7 | 30.4 ± 3.7 | 1.0 ± 0.1 * | 14.6 ± 1.5 |
| BPH20+BPH21-PYL | 102.8 ± 1.1 | 77.9 ± 3.3 | 23.2 ± 2.4 | 34.7 ± 6.1 | 1.2 ± 0.0 | 18.2 ± 2.3 |
| BPH20+BPH32-PYL | 105.2 ± 0.4 | 88.3 ± 1.8 | 19.3 ± 1.7 | 34.3 ± 5.8 | 0.9 ± 0.1 * | 16.0 ± 5.2 |
| BPH21+BPH25-PYL | 102.8 ± 1.3 | 110.5 ± 4.9 ** | 23.0 ± 1.9 | 41.7 ± 3.5 * | 1.1 ± 0.1 | 18.0 ± 3.6 |
| BPH21+BPH17-ptb-PYL | 102.8 ± 2.5 | 107.5 ± 4.6 | 22.4 ± 1.6 | 33.6 ± 4.5 | 1.1 ± 0.1 | 16.0 ± 2.6 |
| BPH25+BPH17-ptb-PYL | 99.6 ± 1.9 | 99.6 ± 2.4 | 19.9 ± 2.0 | 28.9 ± 3.7 | 1.2 ± 0.1 | 15.8 ± 4.8 |
| BPH32+BPH17-ptb-PYL | 99.0 ± 0.0 | 107.6 ± 3.5 | 20.4 ± 2.2 | 26.7 ± 3.4 | 1.1 ± 0.0 | 18.0 ± 1.6 |
| BPH2+BPH3+BPH17-PYL | 103.4 ± 2.9 | 119.0 ± 2.6 * | 22.4 ± 0.7 | 31.4 ± 2.2 | 1.1 ± 0.1 | 16.0 ± 3.4 |
| BPH2+BPH32+BPH17-ptb-PYL | 100.6 ± 1.1 | 102.4 ± 4.4 | 19.1 ± 0.3 | 26.2 ± 3.2 | 1.0 ± 0.1 | 16.6 ± 5.2 |
| BPH20+BPH21+BPH32-PYL | 101.2 ± 0.4 | 91.7 ± 3.1 | 19.3 ± 1.4 | 26.6 ± 4.5 | 1.0 ± 0.1 | 17.8 ± 2.8 |
| Taichung 65 | 99.2 ± 0.4 | 91.4 ± 2.7 | 21.2 ± 0.7 | 29.7 ± 4.2 | 1.1 ± 0.1 | 14.6 ± 1.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, C.D.; Verdeprado, H.; Zita, D.; Sanada-Morimura, S.; Matsumura, M.; Virk, P.S.; Brar, D.S.; Horgan, F.G.; Yasui, H.; Fujita, D. The Development and Characterization of Near-Isogenic and Pyramided Lines Carrying Resistance Genes to Brown Planthopper with the Genetic Background of Japonica Rice (Oryza sativa L.). Plants 2019, 8, 498. https://doi.org/10.3390/plants8110498
Nguyen CD, Verdeprado H, Zita D, Sanada-Morimura S, Matsumura M, Virk PS, Brar DS, Horgan FG, Yasui H, Fujita D. The Development and Characterization of Near-Isogenic and Pyramided Lines Carrying Resistance Genes to Brown Planthopper with the Genetic Background of Japonica Rice (Oryza sativa L.). Plants. 2019; 8(11):498. https://doi.org/10.3390/plants8110498
Chicago/Turabian StyleNguyen, Cuong D., Holden Verdeprado, Demeter Zita, Sachiyo Sanada-Morimura, Masaya Matsumura, Parminder S. Virk, Darshan S. Brar, Finbarr G. Horgan, Hideshi Yasui, and Daisuke Fujita. 2019. "The Development and Characterization of Near-Isogenic and Pyramided Lines Carrying Resistance Genes to Brown Planthopper with the Genetic Background of Japonica Rice (Oryza sativa L.)" Plants 8, no. 11: 498. https://doi.org/10.3390/plants8110498