
Decrypting Molecular Mechanisms Involved in Counteracting Copper and Nickel Toxicity in Jack Pine (Pinus banksiana) Based on Transcriptomic Analysis

Abstract
:1. Introduction
2. Results
2.1. Nickel and Copper Toxicity
2.2. Transcriptome Analysis
3. Discussion
3.1. Physiological Mechanisms in P. banksiana Dealing with Soil Nickel and Copper Contamination
3.2. DEG Analysis Reveals Different Patterns of Gene Expression between Copper RG and Nickel RG
3.3. Analysis of the Top Upregulated Genes in Copper-Resistant Plants Compared to Nickel-Resistant Genotypes Reveals Mechanisms That Are Associated with Copper Resistance in Pinus banksiana
3.4. Analysis of the Top Downregulated Genes for Copper-Resistant Plants Compared to Nickel-Resistant Genotypes Reveal Mechanisms That Are Associated with the Response to Excess Nickel
3.5. Analysis of the Top Upregulated Genes for Copper-Susceptible Plants Compared to Nickel-Susceptible Plants
4. Materials and Methods
4.1. Plant Growth and Treatment
4.2. Transcriptome Analysis of Pinus banksiana
4.3. RNA Extraction
4.4. RNA Sequencing and De Novo Transcriptome Assembly
4.5. BLAT Matching and Annotation of Pinus banksiana Genes
4.6. Quantification of Gene Expression and Quality Control (QC) Analysis
4.7. Differential Gene Expression (DGE) Analysis of Pairwise Comparisons
4.8. Analysis of Top Differentially Regulated Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. In Molecular, Clinical and Environmental Toxicology; Luch, A., Ed.; Experientia Supplementum; Springer: Basel, Switzerland, 2012; Volume 101, pp. 133–164. ISBN 978-3-7643-8339-8. [Google Scholar]
- DeForest, D.K.; Brix, K.V.; Adams, W.J. Assessing Metal Bioaccumulation in Aquatic Environments: The Inverse Relationship between Bioaccumulation Factors, Trophic Transfer Factors and Exposure Concentration. Aquat. Toxicol. Amst. Neth. 2007, 84, 236–246. [Google Scholar] [CrossRef]
- Jara-Marini, M.E.; Soto-Jiménez, M.F.; Páez-Osuna, F. Trophic Relationships and Transference of Cadmium, Copper, Lead and Zinc in a Subtropical Coastal Lagoon Food Web from SE Gulf of California. Chemosphere 2009, 77, 1366–1373. [Google Scholar] [CrossRef]
- Jewiss, T. The Mining History of the Sudbury Area|Earth Sciences Museum. Available online: https://uwaterloo.ca/earth-sciences-museum/resources/mining-canada/mining-history-sudbury-area (accessed on 1 January 2022).
- Kramer, D.M.; Holness, D.L.; Haynes, E.; McMillan, K.; Berriault, C.; Kalenge, S.; Lightfoot, N. From Awareness to Action: Sudbury, Mining and Occupational Disease in a Time of Change. Work 2017, 58, 149–162. [Google Scholar] [CrossRef]
- Nkongolo, K.K.; Spiers, G.; Beckett, P.; Narendrula, R.; Theriault, G.; Tran, A.; Kalubi, K.N. Long-Term Effects of Liming on Soil Chemistry in Stable and Eroded Upland Areas in a Mining Region. Water. Air. Soil Pollut. 2013, 224, 1618. [Google Scholar] [CrossRef]
- Schindler, M. A Mineralogical and Geochemical Study of Slag from the Historical O’donnell Roast Yards, Sudbury, Ontario, Canada. Can. Mineral. 2014, 52, 433–452. [Google Scholar] [CrossRef]
- Andreazza, R.; Okeke, B.C.; Pieniz, S.; Bento, F.M.; Camargo, F.A.O. Biosorption and Bioreduction of Copper from Different Copper Compounds in Aqueous Solution. Biol. Trace Elem. Res. 2013, 152, 411–416. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An Overview of Uptake, Essentiality and Toxicity in Plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef]
- Höhner, R.; Pribil, M.; Herbstová, M.; Lopez, L.S.; Kunz, H.-H.; Li, M.; Wood, M.; Svoboda, V.; Puthiyaveetil, S.; Leister, D.; et al. Plastocyanin Is the Long-Range Electron Carrier between Photosystem II and Photosystem I in Plants. Proc. Natl. Acad. Sci. USA 2020, 117, 15354–15362. [Google Scholar] [CrossRef]
- Cao, J.; Li, X.; Lv, Y.; Ding, L. Comparative Analysis of the Phytocyanin Gene Family in 10 Plant Species: A Focus on Zea Mays. Front. Plant Sci. 2015, 6, 146056. [Google Scholar] [CrossRef]
- Mansilla, N.; Welchen, E.; Gonzalez, D.H. Arabidopsis SCO Proteins Oppositely Influence Cytochrome c Oxidase Levels and Gene Expression during Salinity Stress. Plant Cell Physiol. 2019, 60, 2769–2784. [Google Scholar] [CrossRef]
- Barcelos, J.P.Q.; Reis, H.P.G.; Godoy, C.V.; Gratão, P.L.; Furlani Junior, E.; Putti, F.F.; Campos, M.; Reis, A.R. Impact of Foliar Nickel Application on Urease Activity, Antioxidant Metabolism and Control of Powdery Mildew (Microsphaera diffusa) in Soybean Plants. Plant Pathol. 2018, 67, 1502–1513. [Google Scholar] [CrossRef]
- Urra, M.; Buezo, J.; Royo, B.; Cornejo, A.; López-Gómez, P.; Cerdán, D.; Esteban, R.; Martínez-Merino, V.; Gogorcena, Y.; Tavladoraki, P.; et al. The Importance of the Urea Cycle and Its Relationships to Polyamine Metabolism during Ammonium Stress in Medicago truncatula. J. Exp. Bot. 2022, 73, 5581–5595. [Google Scholar] [CrossRef]
- Lin, S.-L.; Wu, L. Effects of Copper Concentration on Mineral Nutrient Uptake and Copper Accumulation in Protein of Copper-Tolerant and Nontolerant Lotus purshianus L. Ecotoxicol. Environ. Saf. 1994, 29, 214–228. [Google Scholar] [CrossRef]
- Ivanov, Y.V.; Kartashov, A.V.; Ivanova, A.I.; Savochkin, Y.V.; Kuznetsov, V.V. Effects of Copper Deficiency and Copper Toxicity on Organogenesis and Some Physiological and Biochemical Responses of Scots Pine (Pinus Sylvestris L.) Seedlings Grown in Hydroculture. Environ. Sci. Pollut. Res. 2016, 23, 17332–17344. [Google Scholar] [CrossRef]
- Martins, L.L.; Mourato, M.P. Effect of Excess Copper on Tomato Plants: Growth Parameters, Enzyme Activities, Chlorophyll, and Mineral Content. J. Plant Nutr. 2006, 29, 2179–2198. [Google Scholar] [CrossRef]
- Ghasemi, R.; Ghaderian, S.M.; Krämer, U. Interference of Nickel with Copper and Iron Homeostasis Contributes to Metal Toxicity Symptoms in the Nickel Hyperaccumulator Plant Alyssum Inflatum. New Phytol. 2009, 184, 566–580. [Google Scholar] [CrossRef]
- Rubio, M.I.; Escrig, I.; Martínez-Cortina, C.; López-Benet, F.J.; Sanz, A. Cadmium and Nickel Accumulation in Rice Plants. Effects on Mineral Nutrition and Possible Interactions of Abscisic and Gibberellic Acids. Plant Growth Regul. 1994, 14, 151–157. [Google Scholar] [CrossRef]
- Yang, X.; Baligar, V.C.; Martens, D.C.; Clark, R.B. Plant Tolerance to Nickel Toxicity: II Nickel Effects on Influx and Transport of Mineral Nutrients in Four Plant Species. J. Plant Nutr. 1996, 19, 265–279. [Google Scholar] [CrossRef]
- Jegerschoeld, C.; Arellano, J.B.; Schroeder, W.P.; van Kan, P.J.M.; Baron, M.; Styring, S. Copper(II) Inhibition of Electron Transfer through Photosystem II Studied by EPR Spectroscopy. Biochemistry 1995, 34, 12747–12754. [Google Scholar] [CrossRef]
- Mohanty, N.; Vass, I.; Demeter, S. Copper Toxicity Affects Photosystem II Electron Transport at the Secondary Quinone Acceptor, QB1. Plant Physiol. 1989, 90, 175–179. [Google Scholar] [CrossRef]
- Mohanty, N.; Vass, I.; Demeter, S. Impairment of Photosystem 2 Activity at the Level of Secondary Quinone Electron Acceptor in Chloroplasts Treated with Cobalt, Nickel and Zinc Ions. Physiol. Plant. 1989, 76, 386–390. [Google Scholar] [CrossRef]
- Boisvert, S.; Joly, D.; Leclerc, S.; Govindachary, S.; Harnois, J.; Carpentier, R. Inhibition of the Oxygen-Evolving Complex of Photosystem II and Depletion of Extrinsic Polypeptides by Nickel. BioMetals 2007, 20, 879–889. [Google Scholar] [CrossRef]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental Relevance of Heavy Metal-Substituted Chlorophylls Using the Example of Water Plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef]
- Batool, S. Effect of Nickel Toxicity on Growth, Photosynthetic Pigments and Dry Matter Yield of Cicer Arietinum L. Varieties. Asian J. Agri. Biol. 2018, 6, 143–148. [Google Scholar]
- Baran, U.; Ekmekçi, Y. Correction to: Physiological, Photochemical, and Antioxidant Responses of Wild and Cultivated Carthamus Species Exposed to Nickel Toxicity and Evaluation of Their Usage Potential in Phytoremediation. Environ. Sci. Pollut. Res. 2021, 28, 61869–61870. [Google Scholar] [CrossRef]
- Küpper, H.; Küpper, F.; Spiller, M. In Situ Detection of Heavy Metal Substituted Chlorophylls in Water Plants. Photosynth. Res. 1998, 58, 123–133. [Google Scholar] [CrossRef]
- Opdenakker, K.; Remans, T.; Keunen, E.; Vangronsveld, J.; Cuypers, A. Exposure of Arabidopsis Thaliana to Cd or Cu Excess Leads to Oxidative Stress Mediated Alterations in MAPKinase Transcript Levels. Environ. Exp. Bot. 2012, 83, 53–61. [Google Scholar] [CrossRef]
- Wang, L.; Yang, L.; Yang, F.; Li, X.; Song, Y.; Wang, X.; Hu, X. Involvements of H2O2 and Metallothionein in NO-Mediated Tomato Tolerance to Copper Toxicity. J. Plant Physiol. 2010, 167, 1298–1306. [Google Scholar] [CrossRef]
- Kucková, L.; Jomová, K.; Švorcová, A.; Valko, M.; Segľa, P.; Moncoľ, J.; Kožíšek, J. Synthesis, Crystal Structure, Spectroscopic Properties and Potential Biological Activities of Salicylate–Neocuproine Ternary Copper(II) Complexes. Molecules 2015, 20, 2115–2137. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Effect of Nickel on ROS Content and Antioxidative Enzyme Activities in Wheat Leaves. BioMetals 2006, 20, 27–36. [Google Scholar] [CrossRef]
- Baccouch, S.; Chaoui, A.; El Ferjani, E. Nickel Toxicity Induces Oxidative Damage in Zea mays Roots. J. Plant Nutr. 2001, 24, 1085–1097. [Google Scholar] [CrossRef]
- Panou-Filotheou, H. Effects of Copper Toxicity on Leaves of Oregano (Origanum Vulgare Subsp. Hirtum). Ann. Bot. 2001, 88, 207–214. [Google Scholar] [CrossRef]
- Molas, J. Changes in Morphological and Anatomical Structure of Cabbage (Brassica Oleracea L.) Outer Leaves and in Ultrastructure of Their Chloroplasts Caused by an in Vitro Excess of Nickel. Photosynthetica 1998, 34, 513–522. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Riffat, A.; Hussain, M.; Hameed, M.; Alvi, A.K. Toxicity and Tolerance of Nickel in Sunflower (Helianthus Annuus L.). Environ. Sci. Pollut. Res. 2023, 30, 50346–50363. [Google Scholar] [CrossRef]
- Marques, D.M.; da Silva, A.B.; Mantovani, J.R.; Magalhães, P.C.; de Souza, T.C. Root Morphology and Leaf Gas Exchange in Peltophorum Dubium (Spreng.) Taub. (Caesalpinioideae) Exposed to Copper-Induced Toxicity. S. Afr. J. Bot. 2019, 121, 186–192. [Google Scholar] [CrossRef]
- Kulikova, A.L.; Kuznetsova, N.A.; Kholodova, V.P. Effect of Copper Excess in Environment on Soybean Root Viability and Morphology. Russ. J. Plant Physiol. 2011, 58, 836–843. [Google Scholar] [CrossRef]
- Sancenón, V.; Puig, S.; Mateu-Andrés, I.; Dorcey, E.; Thiele, D.J.; Peñarrubia, L. The Arabidopsis Copper Transporter COPT1 Functions in Root Elongation and Pollen Development*. J. Biol. Chem. 2004, 279, 15348–15355. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 Functions as a Metal Efflux Transporter Limiting Excess Zinc, Copper and Cadmium Accumulation in Rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef]
- del Pozo, T.; Cambiazo, V.; González, M. Gene Expression Profiling Analysis of Copper Homeostasis in Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2010, 393, 248–252. [Google Scholar] [CrossRef]
- Wintz, H.; Fox, T.; Wu, Y.-Y.; Feng, V.; Chen, W.; Chang, H.-S.; Zhu, T.; Vulpe, C. Expression Profiles of Arabidopsis Thaliana in Mineral Deficiencies Reveal Novel Transporters Involved in Metal Homeostasis. J. Biol. Chem. 2003, 278, 47644–47653. [Google Scholar] [CrossRef]
- Nishida, S.; Tanikawa, R.; Ishida, S.; Yoshida, J.; Mizuno, T.; Nakanishi, H.; Furuta, N. Elevated Expression of Vacuolar Nickel Transporter Gene IREG2 Is Associated With Reduced Root-to-Shoot Nickel Translocation in Noccaea Japonica. Front. Plant Sci. 2020, 11, 610. [Google Scholar] [CrossRef]
- Kozak, K.; Papierniak-Wygladala, A.; Palusińska, M.; Barabasz, A.; Antosiewicz, D.M. Regulation and Function of Metal Uptake Transporter NtNRAMP3 in Tobacco. Front. Plant Sci. 2022, 13, 867967. [Google Scholar] [CrossRef]
- Shaheen, S.; Ahmad, R.; Mahmood, Q.; Pervez, A.; Maroof Shah, M.; Hafeez, F. Gene Expression and Biochemical Response of Giant Reed under Ni and Cu Stress. Int. J. Phytoremediation 2019, 21, 1474–1485. [Google Scholar] [CrossRef]
- Andrés-Colás, N.; Sancenón, V.; Rodríguez-Navarro, S.; Mayo, S.; Thiele, D.J.; Ecker, J.R.; Puig, S.; Peñarrubia, L. The Arabidopsis Heavy Metal P-Type ATPase HMA5 Interacts with Metallochaperones and Functions in Copper Detoxification of Roots. Plant J. 2006, 45, 225–236. [Google Scholar] [CrossRef]
- Deng, F.; Yamaji, N.; Xia, J.; Ma, J.F. A Member of the Heavy Metal P-Type ATPase OsHMA5 Is Involved in Xylem Loading of Copper in Rice. Plant Physiol. 2013, 163, 1353–1362. [Google Scholar] [CrossRef]
- Lee, S.; Kim, Y.-Y.; Lee, Y.; An, G. Rice P1B-Type Heavy-Metal ATPase, OsHMA9, Is a Metal Efflux Protein. Plant Physiol. 2007, 145, 831–842. [Google Scholar] [CrossRef]
- Seigneurin-Berny, D.; Gravot, A.; Auroy, P.; Mazard, C.; Kraut, A.; Finazzi, G.; Grunwald, D.; Rappaport, F.; Vavasseur, A.; Joyard, J.; et al. HMA1, a New Cu-ATPase of the Chloro Plast Envelope, Is Essential for Growth under Adverse Light Conditions. J. Biol. Chem. 2006, 281, 2882–2892. [Google Scholar] [CrossRef]
- Boutigny, S.; Sautron, E.; Finazzi, G.; Rivasseau, C.; Frelet-Barrand, A.; Pilon, M.; Rolland, N.; Seigneurin-Berny, D. HMA1 and PAA1, Two Chloroplast-Envelope PIB-ATPases, Play Distinct Roles in Chloroplast Copper Homeostasis. J. Exp. Bot. 2014, 65, 1529–1540. [Google Scholar] [CrossRef]
- Sautron, E.; Mayerhofer, H.; Giustini, C.; Pro, D.; Crouzy, S.; Ravaud, S.; Pebay-Peyroula, E.; Rolland, N.; Catty, P.; Seigneurin-Berny, D. HMA6 and HMA8 Are Two Chloroplast Cu+-ATPases with Different Enzymatic Properties. Biosci. Rep. 2015, 35, e00201. [Google Scholar] [CrossRef]
- Shikanai, T.; Müller-Moulé, P.; Munekage, Y.; Niyogi, K.K.; Pilon, M. PAA1, a P-Type ATPase of Arabidopsis, Functions in Copper Transport in Chloroplasts. Plant Cell 2003, 15, 1333–1346. [Google Scholar] [CrossRef]
- Mayerhofer, H.; Sautron, E.; Rolland, N.; Catty, P.; Seigneurin-Berny, D.; Pebay-Peyroula, E.; Ravaud, S. Structural Insights into the Nucleotide-Binding Domains of the P1B-Type ATPases HMA6 and HMA8 from Arabidopsis Thaliana. PLoS ONE 2016, 11, e0165666. [Google Scholar] [CrossRef] [PubMed]
- Tapken, W.; Ravet, K.; Pilon, M. Plastocyanin Controls the Stabilization of the Thylakoid Cu-Transporting P-Type ATPase PAA2/HMA8 in Response to Low Copper in Arabidopsis. J. Biol. Chem. 2012, 287, 18544–18550. [Google Scholar] [CrossRef] [PubMed]
- Tapken, W.; Ravet, K.; Shahbaz, M.; Pilon, M. Regulation of Cu Delivery to Chloroplast Proteins. Plant Signal. Behav. 2015, 10, e1046666. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lacey, R.F.; Ye, Y.; Lu, J.; Yeh, K.-C.; Xiao, Y.; Li, L.; Wen, C.-K.; Binder, B.M.; Zhao, Y. Triplin, a Small Molecule, Reveals Copper Ion Transport in Ethylene Signaling from ATX1 to RAN1. PLoS Genet. 2017, 13, e1006703. [Google Scholar] [CrossRef]
- Mira, H.; Martínez-García, F.; Peñarrubia, L. Evidence for the Plant-Specific Intercellular Transport of the Arabidopsis Copper Chaperone CCH. Plant J. 2001, 25, 521–528. [Google Scholar] [CrossRef]
- Mari, S.; Gendre, D.; Pianelli, K.; Ouerdane, L.; Lobinski, R.; Briat, J.-F.; Lebrun, M.; Czernic, P. Root-to-Shoot Long-Distance Circulation of Nicotianamine and Nicotianamine-Nickel Chelates in the Metal Hyperaccumulator Thlaspi Caerulescens. J. Exp. Bot. 2006, 57, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Ingle, R.A.; Mugford, S.T.; Rees, J.D.; Campbell, M.M.; Smith, J.A.C. Constitutively High Expression of the Histidine Biosynthetic Pathway Contributes to Nickel Tolerance in Hyperaccumulator Plants. Plant Cell 2005, 17, 2089–2106. [Google Scholar] [CrossRef] [PubMed]
- Irtelli, B.; Petrucci, W.A.; Navari-Izzo, F. Nicotianamine and Histidine/Proline Are, Respectively, the Most Important Copper Chelators in Xylem Sap of Brassica Carinata under Conditions of Copper Deficiency and Excess. J. Exp. Bot. 2008, 60, 269–277. [Google Scholar] [CrossRef]
- Guo, W.-J.; Bundithya, W.; Goldsbrough, P.B. Characterization of the Arabidopsis Metallothionein Gene Family: Tissue-Specific Expression and Induction during Senescence and in Response to Copper. New Phytol. 2003, 159, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Pich, A.; Scholz, G. Translocation of Copper and Other Micronutrients in Tomato Plants (Lycopersicon esculentum Mill.): Nicotianamine-Stimulated Copper Transport in the Xylem. J. Exp. Bot. 1996, 47, 41–47. [Google Scholar] [CrossRef]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal Movement within the Plant: Contribution of Nicotianamine and Yellow Stripe 1-like Transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gendre, D.; Czernic, P.; Conéjéro, G.; Pianelli, K.; Briat, J.-F.; Lebrun, M.; Mari, S. TcYSL3, a Member of the YSL Gene Family from the Hyper-Accumulator Thlaspi Caerulescens, Encodes a Nicotianamine-Ni/Fe Transporter. Plant J. 2006, 49, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hou, M.; Cao, L.; Xia, Y.; Shen, Z.; Hu, Z. Glutathione S-Transferases Modulate Cu Tolerance in Oryza Sativa. Environ. Exp. Bot. 2018, 155, 313–320. [Google Scholar] [CrossRef]
- Li, G.-Z.; Zheng, Y.-X.; Chen, S.-J.; Liu, J.; Wang, P.-F.; Wang, Y.-H.; Guo, T.-C.; Kang, G.-Z. TaWRKY74 Participates Copper Tolerance through Regulation of TaGST1 Expression and GSH Content in Wheat. Ecotoxicol. Environ. Saf. 2021, 221, 112469. [Google Scholar] [CrossRef]
- Arduini, I.; Godbold, D.L.; Onnis, A. Cadmium and Copper Uptake and Distribution in Mediterranean Tree Seedlings. Physiol. Plant. 1996, 97, 111–117. [Google Scholar] [CrossRef]
- Çomaklı, E.; Bingöl, M.S. Heavy Metal Accumulation of Urban Scots Pine (Pinus Sylvestris L.) Plantation. Environ. Monit. Assess. 2021, 193, 192. [Google Scholar] [CrossRef] [PubMed]
- Juranović Cindrić, I.; Zeiner, M.; Starčević, A.; Stingeder, G. Metals in Pine Needles: Characterisation of Bio-Indicators Depending on Species. Int. J. Environ. Sci. Technol. 2019, 16, 4339–4346. [Google Scholar] [CrossRef]
- Beckett, P.; Spiers, G. Sudbury, Canada—40 Years of a Community Regreening on a Smelter-Impacted Landscape Before After. Available online: https://www.atlanticclra.ca/wp-content/uploads/2018/11/Beckett-Presentation.pdf (accessed on 4 October 2022).
- Moarefi, N.; Nkongolo, K.K. Contrasting Tolerance and Gene Expression between White Pine (Pinus strobus) and Jack Pine (P. banksiana) Exposed to an Increasing Nickel Concentration. Ecol. Genet. Genomics 2022, 24, 100124. [Google Scholar] [CrossRef]
- Mehes-Smith, M.; Nkongolo, K.; Cholewa, E.; Mehes-Smith, M.; Nkongolo, K.; Cholewa, E. Coping Mechanisms of Plants to Metal Contaminated Soil. In Environmental Change and Sustainability; IntechOpen: London, UK, 2013; ISBN 978-953-51-1094-1. [Google Scholar]
- Narendrula-Kotha, R.; Theriault, G.; Mehes-Smith, M.; Kalubi, K.; Nkongolo, K. Metal Toxicity and Resistance in Plants and Microorganisms in Terrestrial Ecosystems. In Reviews of Environmental Contamination and Toxicology Volume 249; De Voogt, P., Ed.; Reviews of Environmental Contamination and Toxicology; Springer International Publishing: Cham, Switzerland, 2019; Volume 249, pp. 1–27. ISBN 978-3-030-20193-7. [Google Scholar]
- Park, W.; Ahn, S.-J. HMA3 Is a Key Factor for Differences in Cd- and Zn-Related Phenotype between Arabidopsis Ws and Col-0 Ecotypes. Plant Biotechnol. Rep. 2017, 11, 209–218. [Google Scholar] [CrossRef]
- Song, X.-Q.; Liu, L.-F.; Jiang, Y.-J.; Zhang, B.-C.; Gao, Y.-P.; Liu, X.-L.; Lin, Q.-S.; Ling, H.-Q.; Zhou, Y.-H. Disruption of Secondary Wall Cellulose Biosynthesis Alters Cadmium Translocation and Tolerance in Rice Plants. Mol. Plant 2013, 6, 768–780. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Chen, D.; Han, X.; Li, B.; Yang, Y.; Yang, Y. Comparative Expression Analysis of Heavy Metal ATPase Subfamily Genes between Cd-Tolerant and Cd-Sensitive Turnip Landraces. Plant Divers. 2019, 41, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Theriault, G.; Michael, P.; Nkongolo, K. Comprehensive Transcriptome Analysis of Response to Nickel Stress in White Birch (Betula papyrifera). PLoS ONE 2016, 11, e0153762. [Google Scholar] [CrossRef] [PubMed]
- García de la Torre, V.S.; Coba de la Peña, T.; Pueyo, J.J.; Lucas, M.M. Cadmium-Tolerant and -Sensitive Cultivars Identified by Screening of Medicago Truncatula Germplasm Display Contrasting Responses to Cadmium Stress. Front. Plant Sci. 2021, 12, 595001. [Google Scholar] [CrossRef] [PubMed]
- Repka, V.; Fiala, R.; Čiamporová, M.; Pavlovkin, J. Effects of ZnCl2 on ROS Generation, Plasma Membrane Properties, and Changes in Protein Expression in Grapevine Root Explants. Biologia 2016, 71, 528–537. [Google Scholar] [CrossRef]
- Takács, Z.; Tuba, Z.; Smirnoff, N. Exaggeration of Desiccation Stress by Heavy Metal Pollution in Tortula Ruralis: A Pilot Study. Plant Growth Regul. 2001, 35, 157–160. [Google Scholar] [CrossRef]
- Gantayat, S.; Mania, S.; Pradhan, C.; Das, A.B. Ionic Stress Induced Cytotoxic Effect of Cadmium and Nickel Ions on Roots of Allium Cepa L. Cytologia 2017, 83, 143–148. [Google Scholar] [CrossRef]
- Hung, W.-C.; Huang, D.-D.; Chien, P.-S.; Yeh, C.-M.; Chen, P.-Y.; Chi, W.-C.; Huang, H.-J. Protein Tyrosine Dephosphorylation during Copper-Induced Cell Death in Rice Roots. Chemosphere 2007, 69, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Domash, V.I.; Sharpio, T.P.; Zabreiko, S.A.; Sosnovskaya, T.F. Proteolytic Enzymes and Trypsin Inhibitors of Higher Plants under Stress Conditions. Russ. J. Bioorganic Chem. 2008, 34, 318–322. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [CrossRef] [PubMed]
- Faè, M.; Balestrazzi, A.; Confalonieri, M.; Donà, M.; Macovei, A.; Valassi, A.; Giraffa, G.; Carbonera, D. Copper-Mediated Genotoxic Stress Is Attenuated by the Overexpression of the DNA Repair Gene MtTdp2α (Tyrosyl-DNA Phosphodiesterase 2) in Medicago truncatula Plants. Plant Cell Rep. 2014, 33, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Balestrazzi, A.; Confalonieri, M.; Carbonera, D. The Tyrosyl-DNA Phosphodiesterase Gene Family in Medicago Truncatula Gaertn.: Bioinformatic Investigation and Expression Profiles in Response to Copper- and PEG-Mediated Stress. Planta 2010, 232, 393–407. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Sofy, M.R.; Aldaej, M.I.; Mohamed, H.I. Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage. Sustainability 2020, 12, 4732. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Li, S.; Wang, Z. Effect of Nickel Chloride on Arabidopsis Genomic DNA and Methylation of 18S rDNA. Electron. J. Biotechnol. 2015, 18, 51–57. [Google Scholar] [CrossRef]
- Manna, I.; Bandyopadhyay, M. Differential Expression of Nickel Toxicity on Allium Cepa L. Seeds and Seedlings. Int. J. Environ. Sci. Technol. 2022, 20, 11321–11340. [Google Scholar] [CrossRef]
- Wang, J.; Moeen-ud-din, M.; Yin, R.; Yang, S. ROS Homeostasis Involved in Dose-Dependent Responses of Arabidopsis Seedlings to Copper Toxicity. Genes 2023, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-S.; Im, M.-J.; Nkongolo, K. Determination of DNA Methylation Associated with Acer Rubrum (Red Maple) Adaptation to Metals: Analysis of Global DNA Modifications and Methylation-Sensitive Amplified Polymorphism. Ecol. Evol. 2016, 6, 5749–5760. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Zhuang, K.; Xia, Y.; Zhu, C.; Chen, C.; Hu, Z.; Shen, Z. Hydrilla Verticillata Employs Two Different Ways to Affect DNA Methylation under Excess Copper Stress. Aquat. Toxicol. 2017, 193, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Ren, J.; Wang, X.H.; Liu, T.K.; Hou, X.L.; Li, Y. Ascorbic Acid Alleviates Toxicity Induced by Excess Copper in Brassica Campestris Ssp. Chinensis Makino. Commun. Soil Sci. Plant Anal. 2017, 48, 656–664. [Google Scholar] [CrossRef]
- Ince, A.G.; Karaca, M. Tissue and/or Developmental Stage Specific Methylation of nrDNA in Capsicum Annuum. J. Plant Res. 2021, 134, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Pätsikkä, E.; Kairavuo, M.; Šeršen, F.; Aro, E.-M.; Tyystjärvi, E. Excess Copper Predisposes Photosystem II to Photoinhibition in Vivo by Outcompeting Iron and Causing Decrease in Leaf Chlorophyll. Plant Physiol. 2002, 129, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Pető, A.; Lehotai, N.; Lozano-Juste, J.; León, J.; Tari, I.; Erdei, L.; Kolbert, Z. Involvement of Nitric Oxide and Auxin in Signal Transduction of Copper-Induced Morphological Responses in Arabidopsis Seedlings. Ann. Bot. 2011, 108, 449–457. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M. Inhibition of Photosystem II in the Green Alga Scenedesmus Obliquus by Nickel. Biochem. Physiol. Pflanz. 1993, 188, 363–372. [Google Scholar] [CrossRef]
- Lešková, A.; Zvarík, M.; Araya, T.; Giehl, R.F.H. Nickel Toxicity Targets Cell Wall-Related Processes and PIN2-Mediated Auxin Transport to Inhibit Root Elongation and Gravitropic Responses in Arabidopsis. Plant Cell Physiol. 2020, 61, 519–535. [Google Scholar] [CrossRef] [PubMed]
- Moy, A.; Czajka, K.; Michael, P.; Nkongolo, K. Gene Expression Profiling of Jack Pine (Pinus banksiana) under Copper Stress: Identification of Genes Associated with Copper Resistance. PLoS ONE 2021, 19, e0296027. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, J.; Roland, J.; Peeters, E.; Trombik, T.; Ducos, E.; Nader, J.; Boutry, M. NtPDR1, a Plasma Membrane ABC Transporter from Nicotiana Tabacum, Is Involved in Diterpene Transport. Plant Mol. Biol. 2013, 82, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.B.; Selter, L.L.; Baranwal, V.K.; Arora, D.; Mishra, S.K.; Sirohi, P.; Poonia, A.K.; Chaudhary, R.; Kumar, R.; Krattinger, S.G.; et al. Updated Inventory, Evolutionary and Expression Analyses of G (PDR) Type ABC Transporter Genes of Rice. Plant Physiol. Biochem. 2019, 142, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.R.; Vandenbussche, G.; Roosens, N.; Govaerts, C.; Goormaghtigh, E.; Verbruggen, N. Metal Binding Properties and Structure of a Type III Metallothionein from the Metal Hyperaccumulator Plant Noccaea Caerulescens. Biochim. Biophys. Acta BBA—Proteins Proteomics 2012, 1824, 1016–1023. [Google Scholar] [CrossRef]
- Ogasawara, F.; Kodan, A.; Ueda, K. ABC Proteins in Evolution. FEBS Lett. 2020, 594, 3876–3881. [Google Scholar] [CrossRef] [PubMed]
- Moody, J.E.; Millen, L.; Binns, D.; Hunt, J.F.; Thomas, P.J. Cooperative, ATP-Dependent Association of the Nucleotide Binding Cassettes during the Catalytic Cycle of ATP-Binding Cassette Transporters *. J. Biol. Chem. 2002, 277, 21111–21114. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC Transporter AtPDR8 Is a Cadmium Extrusion Pump Conferring Heavy Metal Resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Lee, K.; Lee, J.; Noh, E.W.; Lee, Y. AtPDR12 Contributes to Lead Resistance in Arabidopsis. Plant Physiol. 2005, 138, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Moons, A. Ospdr9, Which Encodes a PDR-Type ABC Transporter, Is Induced by Heavy Metals, Hypoxic Stress and Redox Perturbations in Rice Roots 1. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jing, W.; Zheng, J.; Jin, Y.; Wu, D.; Cao, C.; Dong, Y.; Shi, X.; Zhang, W. The ATP-Binding Cassette Transporter OsPDR1 Regulates Plant Growth and Pathogen Resistance by Affecting Jasmonates Biosynthesis in Rice. Plant Sci. 2020, 298, 110582. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.; Honsbein, A.; Meda, A.R.; Kirchner, S.; Wipf, D.; von Wirén, N. AtIREG2 Encodes a Tonoplast Transport Protein Involved in Iron-Dependent Nickel Detoxification in Arabidopsis Thaliana Roots. J. Biol. Chem. 2006, 281, 25532–25540. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Jiang, Y.; Rahmati, M.; Chia, J.-C.; Dokuchayeva, T.; Kavulych, Y.; Zavodna, T.-O.; Mendoza, P.N.; Huang, R.; Smieshka, L.M.; et al. YSL3-Mediated Copper Distribution Is Required for Fertility, Seed Size and Protein Accumulation in Brachypodium. Plant Physiol. 2021, 186, 655–676. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Nallusamy, S.; Shanmugam, V.; Loganathan, A.; Muthurajan, R.; Sivathapandian, S.K.; Paramasivam, J.; Duraialagaraja, S. Genome-Wide Understanding of Evolutionary and Functional Relationships of Rice Yellow Stripe-Like (YSL) Transporter Family in Comparison with Other Plant Species. Biologia 2022, 77, 39–53. [Google Scholar] [CrossRef]
- Inoue, H.; Kobayashi, T.; Nozoye, T.; Takahashi, M.; Kakei, Y.; Suzuki, K.; Nakazono, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Rice OsYSL15 Is an Iron-Regulated Iron(III)-Deoxymugineic Acid Transporter Expressed in the Roots and Is Essential for Iron Uptake in Early Growth of the Seedlings *. J. Biol. Chem. 2009, 284, 3470–3479. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.A.; Pierson, A.J.; Panaviene, Z.; Walker, E.L. Yellow Stripe1. Expanded Roles for the Maize Iron-Phytosiderophore Transporter. Plant Physiol. 2004, 135, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and Characterization of a Novel Cadmium-Regulated Yellow Stripe-Like Transporter (SnYSL3) in Solanum Nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Yamaji, N.; Yokosho, K.; Ma, J.F. YSL16 Is a Phloem-Localized Transporter of the Copper-Nicotianamine Complex That Is Responsible for Copper Distribution in Rice. Plant Cell 2012, 24, 3767–3782. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, I.; Nagayama, H. Induction of β-Glucosidase in Botrytis Cinerea by Cell Wall Fractions of the Host Plant. Biosci. Biotechnol. Biochem. 1997, 61, 1073–1076. [Google Scholar] [CrossRef]
- Wang, J.-W.; Li, Y.; Zhang, Y.-X.; Chai, T.-Y. Molecular Cloning and Characterization of a Brassica Juncea Yellow Stripe-like Gene, BjYSL7, Whose Overexpression Increases Heavy Metal Tolerance of Tobacco. Plant Cell Rep. 2013, 32, 651–662. [Google Scholar] [CrossRef] [PubMed]
- DiDonato Jr, R.J.; Roberts, L.A.; Sanderson, T.; Eisley, R.B.; Walker, E.L. Arabidopsis Yellow Stripe-Like2 (YSL2): A Metal-Regulated Gene Encoding a Plasma Membrane Transporter of Nicotianamine–Metal Complexes. Plant J. 2004, 39, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, N.; Xiong, H.; Qiu, W.; Nakanishi, H.; Kobayashi, T.; Nishizawa, N.K.; Zuo, Y. The Yellow Stripe-Like (YSL) Gene Functions in Internal Copper Transport in Peanut. Genes 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Matijevic, L.; Romic, D.; Romic, M. Soil Organic Matter and Salinity Affect Copper Bioavailability in Root Zone and Uptake by Vicia faba L. Plants. Environ. Geochem. Health 2014, 36, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Czajka, K.M.; Michael, P.; Nkongolo, K. Differential Effects of Nickel Dosages on in Vitro and in Vivo Seed Germination and Expression of a High Affinity Nickel-Transport Family Protein (AT2G16800) in Trembling Aspen (Populus tremuloides). Ecotoxicology 2019, 28, 92–102. [Google Scholar] [CrossRef] [PubMed]
- de Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; de Oliveira, L.F.V.; Bodanese Zanettini, M.H.; Margis-Pinheiro, M. Heavy Metal-Associated Isoprenylated Plant Protein (HIPP): Characterization of a Family of Proteins Exclusive to Plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef]
- Crowell, D.N. Functional Implications of Protein Isoprenylation in Plants. Prog. Lipid Res. 2000, 39, 393–408. [Google Scholar] [CrossRef]
- Tehseen, M.; Cairns, N.; Sherson, S.; Cobbett, C.S. Metallochaperone-like Genes in Arabidopsis thaliana. Metallomics 2010, 2, 556–564. [Google Scholar] [CrossRef]
- Barth, O.; Vogt, S.; Uhlemann, R.; Zschiesche, W.; Humbeck, K. Stress Induced and Nuclear Localized HIPP26 from Arabidopsis Thaliana Interacts via Its Heavy Metal Associated Domain with the Drought Stress Related Zinc Finger Transcription Factor ATHB29. Plant Mol. Biol. 2009, 69, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xiao, S.; Li, H.-Y.; Tsao, S.-W.; Chye, M.-L. Arabidopsis Thaliana Acyl-CoA-Binding Protein ACBP2 Interacts with Heavy-Metal-Binding Farnesylated Protein AtFP6. New Phytol. 2009, 181, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Dykema, P.E.; Sipes, P.R.; Marie, A.; Biermann, B.J.; Crowell, D.N.; Randall, S.K. A New Class of Proteins Capable of Binding Transition Metals. Plant Mol. Biol. 1999, 41, 139–150. [Google Scholar] [CrossRef] [PubMed]
- khan, I.u.; Rono, J.K.; Zhang, B.Q.; Liu, X.S.; Wang, M.Q.; Wang, L.L.; Wu, X.C.; Chen, X.; Cao, H.W.; Yang, Z.M. Identification of Novel Rice (Oryza sativa) HPP and HIPP Genes Tolerant to Heavy Metal Toxicity. Ecotoxicol. Environ. Saf. 2019, 175, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Functional Characterization of a Heavy Metal Binding Protein CdI19 from Arabidopsis. Plant J. 2002, 32, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Peng, X.; Wang, X.; Wang, C. The Heavy Metal-Associated Isoprenylated Plant Protein (HIPP) Gene Family Plays a Crucial Role in Cadmium Resistance and Accumulation in the Tea Plant (Camellia sinensis L.). Ecotoxicol. Environ. Saf. 2023, 260, 115077. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Jiang, N.; Wang, M.; Du, Z.; Chen, J.; Huang, Y.; Li, M.; Jin, Y.; Li, J.; Wan, J.; et al. OsHIPP17 Is Involved in Regulating the Tolerance of Rice to Copper Stress. Front. Plant Sci. 2023, 14, 1183445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, H.; Feng, C.; Xu, H.; Huang, X.; Wang, Q.; Duan, X.; Wang, X.; Wei, G.; Huang, L.; et al. Isolation and Characterisation of cDNA Encoding a Wheat Heavy Metal-Associated Isoprenylated Protein Involved in Stress Responses. Plant Biol. 2015, 17, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Buning, C.; Comba, P. Protonation of the Copper(I) Form of the Blue Copper Proteins Plastocyanin and Amicyanin—A Molecular Dynamics Study. Eur. J. Inorg. Chem. 2000, 2000, 1267–1273. [Google Scholar] [CrossRef]
- De Rienzo, F.; Gabdoulline, R.R.; Menziani, M.C.; Wade, R.C. Blue Copper Proteins: A Comparative Analysis of Their Molecular Interaction Properties. Protein Sci. 2000, 9, 1439–1454. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Ramiro, M.V.; Cases, R.; Picorel, R.; Yruela, I. Excess Copper Effect on Growth, Chloroplast Ultrastructure, Oxygen-Evolution Activity and Chlorophyll Fluorescence in Glycine Max Cell Suspensions. Physiol. Plant. 2006, 127, 312–325. [Google Scholar] [CrossRef]
- Nersissian, A.M.; Valentine, J.S.; Mehrabian, Z.B.; Nalbandyan, R.M.; Hart, P.J.; Fraczkiewicz, G.; Czernuszewicz, R.S.; Bender, C.J.; Peisach, J.; Herrmann, R.G. Cloning, Expression, and Spectroscopic Characterization of Cucumis Sativus Stellacyanin in Its Nonglycosylated Form. Protein Sci. 1996, 5, 2184–2192. [Google Scholar] [CrossRef] [PubMed]
- Nersissian, A.M.; Valentine, J.S.; Immoos, C.; Hill, M.G.; Hart, P.J.; Williams, G.; Herrmann, R.G. Uclacyanins, Stellacyanins, and Plantacyanins Are Distinct Subfamilies of Phytocyanins: Plant-Specific Mononuclear Blue Copper Proteins. Protein Sci. 1998, 7, 1915–1929. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.D.; Dias, M.A.D.L.; Funkhouser, E.A.; Newton, R.J.; Cairney, J. Gene Expression under Water Deficit in Loblolly Pine (Pinus taeda): Isolation and Characterization of cDNA Clones. Physiol. Plant. 1996, 97, 139–148. [Google Scholar] [CrossRef]
- Drew, J.E.; Gatehouse, J.A. Isolation and Characterization of a Pea Pod cDNA Encoding a Putative Blue Copper Protein Correlated with Lignin Deposition. J. Exp. Bot. 1994, 45, 1873–1884. [Google Scholar] [CrossRef]
- Zhu, W.; Gao, E.; Shaban, M.; Wang, Y.; Wang, H.; Nie, X.; Zhu, L. GhUMC1, a Blue Copper-Binding Protein, Regulates Lignin Synthesis and Cotton Immune Response. Biochem. Biophys. Res. Commun. 2018, 504, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-P.; Björkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A Pigment-Binding Protein Essential for Regulation of Photosynthetic Light Harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Nield, J.; Funk, C.; Barber, J. Supermolecular Structure of Photosystem II and Location of the PsbS Protein. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2000, 355, 1337–1344. [Google Scholar] [CrossRef]
- Nicol, L.; Nawrocki, W.J.; Croce, R. Disentangling the Sites of Non-Photochemical Quenching in Vascular Plants. Nat. Plants 2019, 5, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Gruszecki, W.I.; Grudzinski, W.; Matula, M.; Kernen, P.; Krupa, Z. Light-Induced Excitation Quenching and Structural Transition in Light-Harvesting Complex II. Photosynth. Res. 1999, 59, 175–185. [Google Scholar] [CrossRef]
- Gilmore, A.M.; Hazlett, T.L.; Debrunner, P.G. Govindjee Photosystem II Chlorophyll a Fluorescence Lifetimes and Intensity Are Independent of the Antenna Size Differences between Barley Wild-Type and Chlorina Mutants: Photochemical Quenching and Xanthophyll Cycle-Dependent Nonphotochemical Quenching of Fluorescence. Photosynth. Res. 1996, 48, 171–187. [Google Scholar] [PubMed]
- Dreuw, A.; Fleming, G.R.; Head-Gordon, M. Role of Electron-Transfer Quenching of Chlorophyll Fluorescence by Carotenoids in Non-Photochemical Quenching of Green Plants. Biochem. Soc. Trans. 2005, 33, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, T.; Feng, B.; Zhang, C.; Peng, S.; Zhang, X.; Fu, G.; Tao, L. Non-Photochemical Quenching Plays a Key Role in Light Acclimation of Rice Plants Differing in Leaf Color. Front. Plant Sci. 2017, 7, 1968. [Google Scholar] [CrossRef] [PubMed]
- Eudes, A.; Pereira, J.H.; Yogiswara, S.; Wang, G.; Teixeira Benites, V.; Baidoo, E.E.K.; Lee, T.S.; Adams, P.D.; Keasling, J.D.; Loqué, D. Exploiting the Substrate Promiscuity of Hydroxycinnamoyl-CoA:Shikimate Hydroxycinnamoyl Transferase to Reduce Lignin. Plant Cell Physiol. 2016, 57, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.L.; Costa, M.A.; Laskar, D.D.; Moinuddin, S.G.A.; Lee, C.; Davin, L.B.; Lewis, N.G. RNAi Modulation of Chlorogenic Acid and Lignin Deposition in Nicotiana Tabacum and Insufficient Compensatory Metabolic Cross-Talk. J. Nat. Prod. 2021, 84, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Kriegshauser, L.; Knosp, S.; Grienenberger, E.; Tatsumi, K.; Gütle, D.D.; Sørensen, I.; Herrgott, L.; Zumsteg, J.; Rose, J.K.C.; Reski, R.; et al. Function of the HYDROXYCINNAMOYL-CoA:SHIKIMATE HYDROXYCINNAMOYL TRANSFERASE Is Evolutionarily Conserved in Embryophytes. Plant Cell 2021, 33, 1472–1491. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Yang, Z.; Ma, Y.; Li, S.; Wang, M.; Li, Y.; Liu, Z.; Jia, C.; Han, L.; Chao, Y. Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia Japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis Stolonifera. Int. J. Mol. Sci. 2022, 23, 9500. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.; Qi, Q.; Li, S.; Ruan, B.; Jiang, X.; Gai, Y. Characterization and Functional Analysis of the Hydroxycinnamoyl-CoA: Shikimate Hydroxycinnamoyl Transferase (HCT) Gene Family in Poplar. PeerJ 2021, 9, e10741. [Google Scholar] [CrossRef] [PubMed]
- Varbanova, M.; Porter, K.; Lu, F.; Ralph, J.; Hammerschmidt, R.; Jones, A.D.; Day, B. Molecular and Biochemical Basis for Stress-Induced Accumulation of Free and Bound p-Coumaraldehyde in Cucumber. Plant Physiol. 2011, 157, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of Hydroxycinnamoyl-Coenzyme A Shikimate/Quinate Hydroxycinnamoyltransferase Affects Phenylpropanoid Biosynthesis[W]. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef]
- Man Ha, C.; Fine, D.; Bhatia, A.; Rao, X.; Martin, M.Z.; Engle, N.L.; Wherritt, D.J.; Tschaplinski, T.J.; Sumner, L.W.; Dixon, R.A. Ectopic Defense Gene Expression Is Associated with Growth Defects in Medicago Truncatula Lignin Pathway Mutants1 [OPEN]. Plant Physiol. 2019, 181, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Ralph, J.; Akiyama, T.; Flint, H.; Phillips, L.; Torr, K.; Nanayakkara, B.; Te Kiri, L. Exploring Lignification in Conifers by Silencing Hydroxycinnamoyl-CoA:Shikimate Hydroxycinnamoyltransferase in Pinus Radiata. Proc. Natl. Acad. Sci. USA 2007, 104, 11856–11861. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Liu, Y.; Lyu, J.; Zhao, S.; Wang, Y. Chemical and Structural Responses to Downregulated P-Hydroxycinnamoyl-Coenzyme A: Quinate/Shikimate p-Hydroxycinnamoyltransferase in Poplar Cell Walls. Front. Plant Sci. 2022, 12, 679230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yang, S.; Lu, M.; Zhao, S.; Cai, L.; Zhang, Y.; Zhao, R.; Lv, J. Structure and Monomer Ratio of Lignin in C3H and HCT RNAi Transgenic Poplar Saplings. ChemistrySelect 2020, 5, 7164–7169. [Google Scholar] [CrossRef]
- Zhao, D.; Xu, C.; Luan, Y.; Shi, W.; Tang, Y.; Tao, J. Silicon Enhances Stem Strength by Promoting Lignin Accumulation in Herbaceous Peony (Paeonia lactiflora Pall.). Int. J. Biol. Macromol. 2021, 190, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Marjamaa, K.; Kukkola, E.; Lundell, T.; Karhunen, P.; Saranpää, P.; Fagerstedt, K.V. Monolignol Oxidation by Xylem Peroxidase Isoforms of Norway Spruce (Picea abies) and Silver Birch (Betula pendula). Tree Physiol. 2006, 26, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kirui, A.; Dickwella Widanage, M.C.; Mentink-Vigier, F.; Cosgrove, D.J.; Wang, T. Lignin-Polysaccharide Interactions in Plant Secondary Cell Walls Revealed by Solid-State NMR. Nat. Commun. 2019, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Feijao, C.; Morreel, K.; Anders, N.; Tryfona, T.; Busse-Wicher, M.; Kotake, T.; Boerjan, W.; Dupree, P. Hydroxycinnamic Acid-Modified Xylan Side Chains and Their Cross-Linking Products in Rice Cell Walls Are Reduced in the Xylosyl Arabinosyl Substitution of Xylan 1 Mutant. Plant J. 2022, 109, 1152–1167. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, C.; Costa, L.T.; Abud, Y.; Biancatto, L.; Miguens, F.C.; de Souza, W. Sugarcane Cell Wall Structure and Lignin Distribution Investigated by Confocal and Electron Microscopy. Microsc. Res. Tech. 2013, 76, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Bezrukova, M.V.; Fatkhutdinova, R.A.; Lubyanova, A.R.; Murzabaev, A.R.; Fedyaev, V.V.; Shakirova, F.M. Lectin Involvement in the Development of Wheat Tolerance to Cadmium Toxicity. Russ. J. Plant Physiol. 2011, 58, 1048–1054. [Google Scholar] [CrossRef]
- Ren, C.; Qi, Y.; Huang, G.; Yao, S.; You, J.; Hu, H. Contributions of Root Cell Wall Polysaccharides to Cu Sequestration in Castor (Ricinus communis L.) Exposed to Different Cu Stresses. J. Environ. Sci. 2020, 88, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bak, G.; Burgin, T.; Barnes, W.J.; Mayes, H.B.; Peña, M.J.; Urbanowicz, B.R.; Nielsen, E. Biochemical and Genetic Analysis Identify CSLD3 as a Beta-1,4-Glucan Synthase That Functions during Plant Cell Wall Synthesis[OPEN]. Plant Cell 2020, 32, 1749–1767. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Brandizzi, F.; Liepman, A.H.; Keegstra, K. Arabidopsis Mannan Synthase CSLA9 and Glucan Synthase CSLC4 Have Opposite Orientations in the Golgi Membrane. Plant J. 2010, 64, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Li, L.; Sun, Y.-H.; Chiang, V.L. The Cellulose Synthase Gene Superfamily and Biochemical Functions of Xylem-Specific Cellulose Synthase-Like Genes in Populus trichocarpa. Plant Physiol. 2006, 142, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Little, A.; Lahnstein, J.; Jeffery, D.W.; Khor, S.F.; Schwerdt, J.G.; Shirley, N.J.; Hooi, M.; Xing, X.; Burton, R.A.; Bulone, V. A Novel (1,4)-β-Linked Glucoxylan Is Synthesized by Members of the Cellulose Synthase-Like F Gene Family in Land Plants. ACS Cent. Sci. 2019, 5, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Cocuron, J.-C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A Gene from the Cellulose Synthase-like C Family Encodes a β-1,4 Glucan Synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550–8555. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xiong, G.; Li, R.; Cui, J.; Tang, D.; Zhang, B.; Pauly, M.; Cheng, Z.; Zhou, Y. Rice Cellulose Synthase-like D4 Is Essential for Normal Cell-Wall Biosynthesis and Plant Growth. Plant J. 2009, 60, 1055–1069. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Himmel, M.E.; Ding, S.-Y. Visualizing Chemical Functionality in Plant Cell Walls. Biotechnol. Biofuels 2017, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Salmén, L.; Bergström, E. Cellulose Structural Arrangement in Relation to Spectral Changes in Tensile Loading FTIR. Cellulose 2009, 16, 975–982. [Google Scholar] [CrossRef]
- McCann, M.C.; Roberts, K. Changes in Cell Wall Architecture during Cell Elongation. J. Exp. Bot. 1994, 45, 1683–1691. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Wang, X.; Durachko, D.M.; Zhang, S.; Cosgrove, D.J. Molecular Insights into the Complex Mechanics of Plant Epidermal Cell Walls. Science 2021, 372, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Teeples, M.; Lin, L.; de Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis Gene Regulatory Network for Secondary Cell Wall Synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, S.; Xiong, L.; Hong, Y.; Chen, Y. Cellulose-Hemicellulose Interaction in Wood Secondary Cell-Wall. Model. Simul. Mater. Sci. Eng. 2015, 23, 085010. [Google Scholar] [CrossRef]
- Xiao, Y.; Wu, X.; Liu, D.; Yao, J.; Liang, G.; Song, H.; Ismail, A.M.; Luo, J.-S.; Zhang, Z. Cell Wall Polysaccharide-Mediated Cadmium Tolerance Between Two Arabidopsis Thaliana Ecotypes. Front. Plant Sci. 2020, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.T.; Kirienko, D.H.; Sylvester, A.W.; Peter, G.F.; McCarty, D.R.; Koch, K.E. Cellulose Synthase-Like D1 Is Integral to Normal Cell Division, Expansion, and Leaf Development in Maize. Plant Physiol. 2012, 158, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, R.; Tang, Y.; Peng, C.; Wu, L.; Feng, S.; Chen, P.; Wang, Y.; Du, X.; Peng, L. Cotton CSLD3 Restores Cell Elongation and Cell Wall Integrity Mainly by Enhancing Primary Cellulose Production in the Arabidopsis Cesa6 Mutant. Plant Mol. Biol. 2019, 101, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, R.; Feng, S.; Wang, Y.; Wang, Y.; Fan, C.; Li, Y.; Liu, Z.; Schneider, R.; Xia, T.; et al. Three AtCesA6-like Members Enhance Biomass Production by Distinctively Promoting Cell Growth in Arabidopsis. Plant Biotechnol. J. 2018, 16, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Lukowitz, W.; Nickle, T.C.; Meinke, D.W.; Last, R.L.; Conklin, P.L.; Somerville, C.R. Arabidopsis Cyt1 Mutants Are Deficient in a Mannose-1-Phosphate Guanylyltransferase and Point to a Requirement of N-Linked Glycosylation for Cellulose Biosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Seifert, G.; Doblin, M.S.; Johnson, K.L.; Ruprecht, C.; Pfrengle, F.; Bacic, A.; Estevez, J.M. Cracking the “Sugar Code”: A Snapshot of N- and O-Glycosylation Pathways and Functions in Plants Cells. Front. Plant Sci. 2021, 12, 640919. [Google Scholar] [CrossRef] [PubMed]
- Łuczak, M.; Bugajewska, A.; Wojtaszek, P. Inhibitors of Protein Glycosylation or Secretion Change the Pattern of Extracellular Proteins in Suspension-Cultured Cells of Arabidopsis Thaliana. Plant Physiol. Biochem. 2008, 46, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Lerouxel, O.; Mouille, G.; Andème-Onzighi, C.; Bruyant, M.-P.; Séveno, M.; Loutelier-Bourhis, C.; Driouich, A.; Höfte, H.; Lerouge, P. Mutants in DEFECTIVE GLYCOSYLATION, an Arabidopsis Homolog of an Oligosaccharyltransferase Complex Subunit, Show Protein Underglycosylation and Defects in Cell Differentiation and Growth. Plant J. 2005, 42, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Lige, B.; Ma, S.; van Huystee, R.B. The Effects of the Site-Directed Removal of N-Glycosylation from Cationic Peanut Peroxidase on Its Function. Arch. Biochem. Biophys. 2001, 386, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Eberhard, S.; Albersheim, P.; Darvill, A.; York, W.S. Effects of the Mur1 Mutation on Xyloglucans Produced by Suspension-Cultured Arabidopsis Thaliana Cells. Planta 2001, 214, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Lamanchai, K.; Salmon, D.L.; Smirnoff, N.; Sutthinon, P.; Roytrakul, S.; Leetanasaksakul, K.; Kittisenachai, S.; Jantasuriyarat, C. OsVTC1-1 RNAi Mutant with Reduction of Ascorbic Acid Synthesis Alters Cell Wall Sugar Composition and Cell Wall-Associated Proteins. Agronomy 2022, 12, 1272. [Google Scholar] [CrossRef]
- Voxeur, A.; Soubigou-Taconnat, L.; Legée, F.; Sakai, K.; Antelme, S.; Durand-Tardif, M.; Lapierre, C.; Sibout, R. Altered Lignification in Mur1-1 a Mutant Deficient in GDP-L-Fucose Synthesis with Reduced RG-II Cross Linking. PLoS ONE 2017, 12, e0184820. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.; Cicéron, F.; Lerouxel, O.; Breton, C.; de Sanctis, D. The Galactoside 2-α-l-Fucosyltransferase FUT1 from Arabidopsis Thaliana: Crystallization and Experimental MAD Phasing. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2016, 72, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Yang, S.H.; Han, K.-H. Upregulation of an Arabidopsis RING-H2 Gene, XERICO, Confers Drought Tolerance through Increased Abscisic Acid Biosynthesis. Plant J. 2006, 47, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.-E.; Hou, P.; Xiao, F.; Liu, Y. Overexpressing a Novel RING-H2 Finger Protein Gene, OsRHP1, Enhances Drought and Salt Tolerance in Rice (Oryza sativa L.). J. Plant Biol. 2014, 57, 357–365. [Google Scholar] [CrossRef]
- Atkinson, R.G.; Sutherland, P.W.; Johnston, S.L.; Gunaseelan, K.; Hallett, I.C.; Mitra, D.; Brummell, D.A.; Schröder, R.; Johnston, J.W.; Schaffer, R.J. Down-Regulation of POLYGALACTURONASE1 Alters Firmness, Tensile Strength and Water Loss in Apple (Malus x domestica) Fruit. BMC Plant Biol. 2012, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Rowell, R.M.; Pettersen, R.; Tshabalala, M.A. Cell Wall Chemistry; Routledge Handbooks Online; Routledge: London, UK, 2012; ISBN 978-1-4398-5380-1. [Google Scholar]
- Cao, C.; Yang, Z.; Han, L.; Jiang, X.; Ji, G. Study on in Situ Analysis of Cellulose, Hemicelluloses and Lignin Distribution Linked to Tissue Structure of Crop Stalk Internodal Transverse Section Based on FTIR Microspectroscopic Imaging. Cellulose 2015, 22, 139–149. [Google Scholar] [CrossRef]
- Orfila, C.; Huisman, M.M.; Willats, W.G.; van Alebeek, G.-J.W.; Schols, H.A.; Seymour, G.B.; Knox, P.J. Altered Cell Wall Disassembly during Ripening of Cnr Tomato Fruit: Implications for Cell Adhesion and Fruit Softening. Planta 2002, 215, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Halder, S.; Chaudhari, S.R.; Netz, R.R.; Mogurampelly, S. Insights into the Structure and Ion Transport of Pectin-[BMIM][PF6] Electrolytes. J. Chem. Phys. 2023, 159, 154902. [Google Scholar] [CrossRef] [PubMed]
- Rondeau-Mouro, C.; Defer, D.; Leboeuf, E.; Lahaye, M. Assessment of Cell Wall Porosity in Arabidopsis Thaliana by NMR Spectroscopy. Int. J. Biol. Macromol. 2008, 42, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Pyun, Y.R.; Kokini, J.L. Sidechains of Pectins: Some Thoughts on Their Role in Plant Cell Walls and Foods. Food Hydrocoll. 1993, 7, 39–53. [Google Scholar] [CrossRef]
- Forand, A.D.; Finfrock, Y.Z.; Lavier, M.; Stobbs, J.; Qin, L.; Wang, S.; Karunakaran, C.; Wei, Y.; Ghosh, S.; Tanino, K.K. With a Little Help from My Cell Wall: Structural Modifications in Pectin May Play a Role to Overcome Both Dehydration Stress and Fungal Pathogens. Plants 2022, 11, 385. [Google Scholar] [CrossRef] [PubMed]
- Paynel, F.; Schaumann, A.; Arkoun, M.; Douchiche, O.; Morvan, C. Temporal Regulation of Cell-Wall Pectin Methylesterase and Peroxidase Isoforms in Cadmium-Treated Flax Hypocotyl. Ann. Bot. 2009, 104, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Uttam, K.N. Early Stage Detection of Stress Due to Copper on Maize (Zea mays L.) by Laser-Induced Fluorescence and Infrared Spectroscopy. J. Appl. Spectrosc. 2018, 85, 771–780. [Google Scholar] [CrossRef]
- Liu, H.; Ma, Y.; Chen, N.; Guo, S.; Liu, H.; Guo, X.; Chong, K.; Xu, Y. Overexpression of Stress-Inducible OsBURP16, the β Subunit of Polygalacturonase 1, Decreases Pectin Content and Cell Adhesion and Increases Abiotic Stress Sensitivity in Rice. Plant Cell Environ. 2014, 37, 1144–1158. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Takeuchi, H.; Sato, J.; Nakamura, A.; Ichikawa, H.; Yokoyama, R.; Nishitani, K.; Minami, E.; Satoh, S.; Iwai, H. Structural Alteration of Rice Pectin Affects Cell Wall Mechanical Strength and Pathogenicity of the Rice Blast Fungus Under Weak Light Conditions. Plant Cell Physiol. 2021, 62, 641–649. [Google Scholar] [CrossRef]
- Khan, S.; Zafar, A.; Naseem, I. Copper-Redox Cycling by Coumarin-Di(2-Picolyl)Amine Hybrid Molecule Leads to ROS-Mediated DNA Damage and Apoptosis: A Mechanism for Cancer Chemoprevention. Chem. Biol. Interact. 2018, 290, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ji, K.; Liang, B.; Du, Y.; Jiang, L.; Wang, J.; Kai, W.; Zhang, Y.; Zhai, X.; Chen, P.; et al. Suppressing ABA Uridine Diphosphate Glucosyltransferase (SlUGT75C1) Alters Fruit Ripening and the Stress Response in Tomato. Plant J. 2017, 91, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-T.; Liu, F.-F.; Xiao, D.-W.; Jiang, X.-Y.; Li, P.; Zhao, S.-M.; Hou, B.; Li, Y. The Arabidopsis UDP-glycosyltransferase75B1, Conjugates Abscisic Acid and Affects Plant Response to Abiotic Stresses. Plant Mol. Biol. 2020, 102, 389–401. [Google Scholar] [CrossRef]
- Song, J.; Shang, L.; Wang, X.; Xing, Y.; Xu, W.; Zhang, Y.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. MAPK11 Regulates Seed Germination and ABA Signaling in Tomato by Phosphorylating SnRKs. J. Exp. Bot. 2021, 72, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Tuan, P.A.; Izydorczyk, M.S.; Ayele, B.T. Ethylene Regulates Post-Germination Seedling Growth in Wheat through Spatial and Temporal Modulation of ABA/GA Balance. J. Exp. Bot. 2020, 71, 1985–2004. [Google Scholar] [CrossRef] [PubMed]
- Tao, Q.; Jupa, R.; Dong, Q.; Yang, X.; Liu, Y.; Li, B.; Yuan, S.; Yin, J.; Xu, Q.; Li, T.; et al. Abscisic Acid-Mediated Modifications in Water Transport Continuum Are Involved in Cadmium Hyperaccumulation in Sedum Alfredii. Chemosphere 2021, 268, 129339. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Kao, C.H. Role of Abscisic Acid in Cadmium Tolerance of Rice (Oryza sativa L.) Seedlings. Plant Cell Environ. 2003, 26, 867–874. [Google Scholar] [CrossRef]
- Deng, B.; Zhang, W.; Yang, H. Abscisic Acid Decreases Cell Death in Malus Hupehensis Rehd. Under Cd Stress by Reducing Root Cd2+ Influx and Leaf Transpiration. J. Plant Growth Regul. 2022, 41, 639–646. [Google Scholar] [CrossRef]
- Estrada-Melo, A.C.; Chao, C.; Reid, M.S.; Jiang, C.-Z. Overexpression of an ABA Biosynthesis Gene Using a Stress-Inducible Promoter Enhances Drought Resistance in Petunia. Hortic. Res. 2015, 2, 15013. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Y.; Zhu, C.; Yao, X.; Zheng, Z.; Tian, Z.; Cai, X. EkFLS Overexpression Promotes Flavonoid Accumulation and Abiotic Stress Tolerance in Plant. Physiol. Plant. 2021, 172, 1966–1982. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Jang, G.; Um, T.; Kim, J.-K.; Lee, J.S.; Do Choi, Y. The Soluble ABA Receptor PYL8 Regulates Drought Resistance by Controlling ABA Signaling in Arabidopsis. Plant Biotechnol. Rep. 2015, 9, 319–330. [Google Scholar] [CrossRef]
- Tung, S.A.; Smeeton, R.; White, C.A.; Black, C.R.; Taylor, I.B.; Hilton, H.W.; Thompson, A.J. Over-Expression of LeNCED1 in Tomato (Solanum lycopersicum L.) with the rbcS3C Promoter Allows Recovery of Lines That Accumulate Very High Levels of Abscisic Acid and Exhibit Severe Phenotypes. Plant Cell Environ. 2008, 31, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Johannesson, H.; Wang, Y.; Hanson, J.; Engström, P. The Arabidopsis Thaliana Homeobox Gene ATHB5 Is a Potential Regulator of Abscisic Acid Responsiveness in Developing Seedlings. Plant Mol. Biol. 2003, 51, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.K.; Fang, X.Z.; Guan, M.Y.; Ye, Y.Q.; Lin, X.Y.; Du, S.T.; Jin, C.W. Exogenous Abscisic Acid Application Decreases Cadmium Accumulation in Arabidopsis Plants, Which Is Associated with the Inhibition of IRT1-Mediated Cadmium Uptake. Front. Plant Sci. 2014, 5, 721. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Jiang, R.F.; Dunham, S.J.; McGrath, S.P. Cadmium Uptake, Translocation and Tolerance in the Hyperaccumulator Arabidopsis Halleri. New Phytol. 2006, 172, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, H.; Wei, Z.; Sun, H.; He, Y.; Gao, J.; Yang, Z.; You, J. Overexpression of UDP-Glycosyltransferase Genes Enhanced Aluminum Tolerance through Disrupting Cell Wall Polysaccharide Components in Soybean. Plant Soil 2021, 469, 135–147. [Google Scholar] [CrossRef]
- Hu, H.; Qian, P.; Ye, M.; Mu, K.; Wang, S.; Chen, M.; Ma, H. GmUGT73F4 Plays Important Roles in Enhancing Seed Vitality and Tolerance to Abiotic Stresses in Transgenic Arabidopsis. Plant Cell Tissue Organ Cult. PCTOC 2022, 150, 313–328. [Google Scholar] [CrossRef]
- Wang, T.; Li, P.; Mu, T.; Dong, G.; Zheng, C.; Jin, S.; Chen, T.; Hou, B.; Li, Y. Overexpression of UGT74E2, an Arabidopsis IBA Glycosyltransferase, Enhances Seed Germination and Modulates Stress Tolerance via ABA Signaling in Rice. Int. J. Mol. Sci. 2020, 21, 7239. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Guo, J.; Dong, D.; Zhang, M.; Li, Q.; Cao, Y.; Dong, Y.; Chen, C.; Jin, X. UDP-Glycosyltransferase Gene SlUGT73C1 from Solanum Lycopersicum Regulates Salt and Drought Tolerance in Arabidopsis thaliana L. Funct. Integr. Genom. 2023, 23, 320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sun, Y.; Li, M.; Long, R. CrUGT87A1, a UDP-Sugar Glycosyltransferases (UGTs) Gene from Carex Rigescens, Increases Salt Tolerance by Accumulating Flavonoids for Antioxidation in Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 159, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.; Wang, B.; Yu, H.; Li, Q.; Hou, B. The Arabidopsis UGT87A2, a Stress-Inducible Family 1 Glycosyltransferase, Is Involved in the Plant Adaptation to Abiotic Stresses. Physiol. Plant. 2017, 159, 416–432. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.S.; Howe, G.A. A Critical Role for the TIFY Motif in Repression of Jasmonate Signaling by a Stabilized Splice Variant of the JASMONATE ZIM-Domain Protein JAZ10 in Arabidopsis. Plant Cell 2009, 21, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Hakata, M.; Muramatsu, M.; Nakamura, H.; Hara, N.; Kishimoto, M.; Iida-Okada, K.; Kajikawa, M.; Imai-Toki, N.; Toki, S.; Nagamura, Y.; et al. Overexpression of TIFY Genes Promotes Plant Growth in Rice through Jasmonate Signaling. Biosci. Biotechnol. Biochem. 2017, 81, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, F.; Yang, G.; Liu, X.; Peng, S. Identification of TIFY Gene Family in Walnut and Analysis of Its Expression under Abiotic Stresses. BMC Genom. 2022, 23, 190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; You, J.; Chan, Z. Identification and Characterization of TIFY Family Genes in Brachypodium Distachyon. J. Plant Res. 2015, 128, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Zheng, L.; Jin, L.-G.; Liu, Y.-X.; Kong, Y.-N.; Wang, Y.-X.; Yu, T.-F.; Chen, J.; Zhou, Y.-B.; Chen, M.; et al. Genome-Wide Analysis of the Soybean TIFY Family and Identification of GmTIFY10e and GmTIFY10g Response to Salt Stress. Front. Plant Sci. 2022, 13, 845314. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Du, H.; Tang, N.; Li, X.; Xiong, L. Identification and Expression Profiling Analysis of TIFY Family Genes Involved in Stress and Phytohormone Responses in Rice. Plant Mol. Biol. 2009, 71, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, R.; Zhang, T.; Zheng, D.; Li, X.; Zhang, Z.B.; Li, L.G.; Wu, Z.Y. ZmTIFY16, a Novel Maize TIFY Transcription Factor Gene, Promotes Root Growth and Development and Enhances Drought and Salt Tolerance in Arabidopsis and Zea mays. Plant Growth Regul. 2023, 100, 149–160. [Google Scholar] [CrossRef]
- Zhu, D.; Li, R.; Liu, X.; Sun, M.; Wu, J.; Zhang, N.; Zhu, Y. The Positive Regulatory Roles of the TIFY10 Proteins in Plant Responses to Alkaline Stress. PLoS ONE 2014, 9, e111984. [Google Scholar] [CrossRef] [PubMed]
- Maksymiec, W.; Krupa, Z. The Effects of Short-Term Exposition to Cd, Excess Cu Ions and Jasmonate on Oxidative Stress Appearing in Arabidopsis Thaliana. Environ. Exp. Bot. 2006, 57, 187–194. [Google Scholar] [CrossRef]
- Brioudes, F.; Joly, C.; Szécsi, J.; Varaud, E.; Leroux, J.; Bellvert, F.; Bertrand, C.; Bendahmane, M. Jasmonate Controls Late Development Stages of Petal Growth in Arabidopsis Thaliana. Plant J. 2009, 60, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Noir, S.; Bömer, M.; Takahashi, N.; Ishida, T.; Tsui, T.-L.; Balbi, V.; Shanahan, H.; Sugimoto, K.; Devoto, A. Jasmonate Controls Leaf Growth by Repressing Cell Proliferation and the Onset of Endoreduplication While Maintaining a Potential Stand-By Mode. Plant Physiol. 2013, 161, 1930–1951. [Google Scholar] [CrossRef] [PubMed]
- Attaran, E.; Major, I.T.; Cruz, J.A.; Rosa, B.A.; Koo, A.J.K.; Chen, J.; Kramer, D.M.; He, S.Y.; Howe, G.A. Temporal Dynamics of Growth and Photosynthesis Suppression in Response to Jasmonate Signaling. Plant Physiol. 2014, 165, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Sirhindi, G.; Mushtaq, R.; Gill, S.S.; Sharma, P.; Abd_Allah, E.F.; Ahmad, P. Jasmonic Acid and Methyl Jasmonate Modulate Growth, Photosynthetic Activity and Expression of Photosystem II Subunit Genes in Brassica oleracea L. Sci. Rep. 2020, 10, 9322. [Google Scholar] [CrossRef] [PubMed]
- Keramat, B.; Kalantari, K.M.; Arvin, M.J. Effects of Methyl Jasmonate Treatment on Alleviation of Cadmium Damages in Soybean. J. Plant Nutr. 2010, 33, 1016–1025. [Google Scholar] [CrossRef]
- Held, M.; Baldwin, I.T. Soil Degradation Slows Growth and Inhibits Jasmonate-Induced Resistance in Artemisia Vulgaris. Ecol. Appl. 2005, 15, 1689–1700. [Google Scholar] [CrossRef]
- Abdollahi, A.; Farsad-Akhtar, N.; Mohajel Kazemi, E.; Kolahi, M. Investigation of the Combined Effects of Cadmium Chloride, Silver Nitrate, Lead Nitrate, Methyl Jasmonate, and Salicylic Acid on Morphometric and Biochemical Characteristics of St. John’s Wort. Physiol. Mol. Biol. Plants 2023, 29, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, X.; Chen, J.; Tam, N.F.-Y. Combined Toxicity of Cadmium and Copper in Avicennia Marina Seedlings and the Regulation of Exogenous Jasmonic Acid. Ecotoxicol. Environ. Saf. 2015, 113, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Wei, S.; Pogrzeba, M.; Rusinowski, S.; Krzyżak, J.; Jia, G. Exogenous Jasmonic Acid Decreased Cu Accumulation by Alfalfa and Improved Its Photosynthetic Pigments and Antioxidant System. Ecotoxicol. Environ. Saf. 2020, 190, 110176. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.R.; Lorenz, W.W.; Dean, J.F.D. Characterization of a 1-Aminocyclopropane-1-Carboxylate Synthase Gene from Loblolly Pine (Pinus taeda L.). Gene 2008, 413, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Stearns, J.C.; Shah, S.; Greenberg, B.M.; Dixon, D.G.; Glick, B.R. Tolerance of Transgenic Canola Expressing 1-Aminocyclopropane-1-Carboxylic Acid Deaminase to Growth Inhibition by Nickel. Plant Physiol. Biochem. 2005, 43, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Oláh, D.; Molnár, Á.; Szőllősi, R.; Erdei, L.; Ördög, A. Distinct Redox Signalling and Nickel Tolerance in Brassica Juncea and Arabidopsis Thaliana. Ecotoxicol. Environ. Saf. 2020, 189, 109989. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Li, C.; Zhang, F.; Yu, Q.; Gao, S.; Zhang, M.; Tian, H.; Zhang, J.; Yuan, X.; Ding, Z. Ethylene Promotes Cadmium-Induced Root Growth Inhibition through EIN3 Controlled XTH33 and LSU1 Expression in Arabidopsis. Plant Cell Environ. 2018, 41, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Wakeel, A.; Gan, Y. A Model for the Ethylene-Mediated Auxin Distribution under Cr(VI) Stress in Arabidopsis Thaliana. Plant Signal. Behav. 2018, 13, e1473685. [Google Scholar] [CrossRef]
- Mir, I.R.; Rather, B.A.; Sehar, Z.; Masood, A.; Khan, N.A. Nitric Oxide in Co-Ordination with Nitrogen Reverses Cadmium-Inhibited Photosynthetic Activity by Interacting with Ethylene Synthesis, Strengthening the Antioxidant System, and Nitrogen and Sulfur Assimilation in Mustard (Brassica juncea L.). Sci. Hortic. 2023, 314, 111958. [Google Scholar] [CrossRef]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense[W]. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Chen, Y.-C.; Kieber, J.J.; Yoon, G.M. Regulation of the Turnover of ACC Synthases by Phytohormones and Heterodimerization in Arabidopsis. Plant J. 2017, 91, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.R.; Rodrigues dos Reis, A.; Prado, E.R.; Lavres, J.; Pompeu, G.B.; Azevedo, R.A.; Gratão, P.L. New Insights into Cadmium Stressful-Conditions: Role of Ethylene on Selenium-Mediated Antioxidant Enzymes. Ecotoxicol. Environ. Saf. 2019, 186, 109747. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gaddam, S.R.; Singh, D.; Trivedi, P.K. Regulation of Arsenic Stress Response by Ethylene Biosynthesis and Signaling in Arabidopsis Thaliana. Environ. Exp. Bot. 2021, 185, 104408. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, M.; Li, Z.; Niu, Y.; Jin, Q.; Zhu, B.; Xu, Y. Effects of Ethylene Biosynthesis and Signaling on Oxidative Stress and Antioxidant Defense System in Nelumbo Nucifera G. under Cadmium Exposure. Environ. Sci. Pollut. Res. 2020, 27, 40156–40170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Shang, S.; Jabeen, Z.; Zhang, G. Involvement of Ethylene in Alleviation of Cd Toxicity by NaCl in Tobacco Plants. Ecotoxicol. Environ. Saf. 2014, 101, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Peng, S.P.; Shao, C.Y.; Shao, H.B.; Yang, H.C. Ethylene Glycol Tetra-Acetic Acid and Salicylic Acid Improve Anti-Oxidative Ability of Maize Seedling Leaves under Heavy-Metal and Polyethylene Glycol 6000-Simulated Drought Stress. Plant Biosyst.—Int. J. Deal. Asp. Plant Biol. 2014, 148, 96–108. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; E, L.; Yang, J.; Liu, W.; Xu, M.; Ye, J. ZmDRR206 Regulates Nutrient Accumulation in Endosperm through Its Role in Cell Wall Biogenesis during Maize Kernel Development. Int. J. Mol. Sci. 2023, 24, 8735. [Google Scholar] [CrossRef] [PubMed]
- Gang, D.R.; Costa, M.A.; Fujita, M.; Dinkova-Kostova, A.T.; Wang, H.-B.; Burlat, V.; Martin, W.; Sarkanen, S.; Davin, L.B.; Lewis, N.G. Regiochemical Control of Monolignol Radical Coupling: A New Paradigm for Lignin and Lignan Biosynthesis. Chem. Biol. 1999, 6, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sun, W.; Zhou, P.; Wei, H.; Wang, P.; Li, H.; Rehman, S.; Li, D.; Zhuge, Q. Genome-Wide Characterization of Dirigent Proteins in Populus: Gene Expression Variation and Expression Pattern in Response to Marssonina Brunnea and Phytohormones. Forests 2021, 12, 507. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, J.; Wang, Y.; Zhang, X.; Shen, Z.; Hu, Z. Ectopic Expression of Vicia Sativa Caffeoyl-CoA O-Methyltransferase (VsCCoAOMT) Increases the Uptake and Tolerance of Cadmium in Arabidopsis. Environ. Exp. Bot. 2018, 145, 47–53. [Google Scholar] [CrossRef]
- Gao, W.; Wang, X.J.; Yu, C.C.; Feng, W.J.; Hua, D.L.; Kang, G.Z.; Zhao, P. Comparative Morpho-Physiological Analyses Revealed H2O2-Induced Different Cadmium Accumulation in Two Wheat Cultivars (Triticum aestivum L.). Environ. Exp. Bot. 2021, 185, 104395. [Google Scholar] [CrossRef]
- MacFarlane, G.R.; Burchett, M.D. Cellular Distribution of Copper, Lead and Zinc in the Grey Mangrove, Avicennia Marina (Forsk.) Vierh. Aquat. Bot. 2000, 68, 45–59. [Google Scholar] [CrossRef]
- Lewis, N.G.; Davin, L.B. Evolution of Lignan and Neolignan Biochemical Pathways. In Isopentenoids and Other Natural Products; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1994; Volume 562, pp. 202–246. ISBN 978-0-8412-2934-1. [Google Scholar]
- Wei, M.; Yu, C.; Ge, B.; Liu, Y.; Zhang, H.; Duan, C.; Zhang, J.; Mao, T.; Huang, H.; Xie, Y.; et al. The Phenylcoumaran Benzylic Ether Reductase Gene PtPCBER Improves the Salt Tolerance of Transgenic Poplar through Lignan-Mediated Reactive Oxygen Species Scavenging. Environ. Exp. Bot. 2022, 201, 104946. [Google Scholar] [CrossRef]
- Li, L.; Sun, W.; Wang, P.; Li, H.; Rehman, S.; Li, D.; Zhuge, Q. Characterization, Expression, and Functional Analysis of the Pathogenesis-Related Gene PtDIR11 in Transgenic Poplar. Int. J. Biol. Macromol. 2022, 210, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Niculaes, C.; Morreel, K.; Kim, H.; Lu, F.; McKee, L.S.; Ivens, B.; Haustraete, J.; Vanholme, B.; Rycke, R.D.; Hertzberg, M.; et al. Phenylcoumaran Benzylic Ether Reductase Prevents Accumulation of Compounds Formed under Oxidative Conditions in Poplar Xylem. Plant Cell 2014, 26, 3775–3791. [Google Scholar] [CrossRef] [PubMed]
- Bollella, P.; Medici, L.; Tessema, M.; Poloznikov, A.A.; Hushpulian, D.M.; Tishkov, V.I.; Andreu, R.; Leech, D.; Megersa, N.; Marcaccio, M.; et al. Highly Sensitive, Stable and Selective Hydrogen Peroxide Amperometric Biosensors Based on Peroxidases from Different Sources Wired by Os-Polymer: A Comparative Study. Solid State Ion. 2018, 314, 178–186. [Google Scholar] [CrossRef]
- Ji, Y.; Wu, P.; Zhang, J.; Zhang, J.; Zhou, Y.; Peng, Y.; Zhang, S.; Cai, G.; Gao, G. Heavy Metal Accumulation, Risk Assessment and Integrated Biomarker Responses of Local Vegetables: A Case Study along the Le’an River. Chemosphere 2018, 199, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Kawano, T. Roles of the Reactive Oxygen Species-Generating Peroxidase Reactions in Plant Defense and Growth Induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Tugbaeva, A.; Wuriyanghan, H.; Maleva, M. Copper Stress Enhances the Lignification of Axial Organs in Zinnia Elegans. Horticulturae 2022, 8, 558. [Google Scholar] [CrossRef]
- Zhang, A.; Lu, F.; Sun, R.; Ralph, J. Ferulate–Coniferyl Alcohol Cross-Coupled Products Formed by Radical Coupling Reactions. Planta 2009, 229, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, T.; Morreel, K.; Delhomme, N.; Gauthier, A.; Schiffthaler, B.; Nickolov, K.; Brader, G.; Lim, K.-J.; Teeri, T.H.; Street, N.R.; et al. A Key Role for Apoplastic H2O2 in Norway Spruce Phenolic Metabolism. Plant Physiol. 2017, 174, 1449–1475. [Google Scholar] [CrossRef] [PubMed]
- Kidwai, M.; Dhar, Y.V.; Gautam, N.; Tiwari, M.; Ahmad, I.Z.; Asif, M.H.; Chakrabarty, D. Oryza Sativa Class III Peroxidase (OsPRX38) Overexpression in Arabidopsis Thaliana Reduces Arsenic Accumulation Due to Apoplastic Lignification. J. Hazard. Mater. 2019, 362, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wu, J.; Yuan, D.; Zhang, D.; Huang, Z.; Xiao, L.; Yang, C. Perturbation of Auxin Homeostasis Caused by Mitochondrial FtSH4 Gene-Mediated Peroxidase Accumulation Regulates Arabidopsis Architecture. Mol. Plant 2014, 7, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.-M.; Xu, H.-H.; Liu, W.-C.; Lu, Y.-T. Copper Regulates Primary Root Elongation Through PIN1-Mediated Auxin Redistribution. Plant Cell Physiol. 2013, 54, 766–778. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Q.; Xuan, W.; Huang, X.-Y.; Mao, C.; Zhao, F.-J. Cadmium Inhibits Lateral Root Emergence in Rice by Disrupting OsPIN-Mediated Auxin Distribution and the Protective Effect of OsHMA3. Plant Cell Physiol. 2020, 62, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Khan, N.A.; D’Ippolito, I.; Reyes, F. Subtoxic Levels of Some Heavy Metals Cause Differential Root-Shoot Structure, Morphology and Auxins Levels in Arabidopsis thaliana. Plant Physiol. Biochem. 2022, 173, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Mustafiz, A.; Singh, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Genome-Wide Analysis of Rice and Arabidopsis Identifies Two Glyoxalase Genes That Are Highly Expressed in Abiotic Stresses. Funct. Integr. Genomics 2011, 11, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Mustafiz, A.; Ghosh, A.; Tripathi, A.K.; Kaur, C.; Ganguly, A.K.; Bhavesh, N.S.; Tripathi, J.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. A Unique Ni2+ -Dependent and Methylglyoxal-Inducible Rice Glyoxalase I Possesses a Single Active Site and Functions in Abiotic Stress Response. Plant J. 2014, 78, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Welchen, E.; Schmitz, J.; Fuchs, P.; García, L.; Wagner, S.; Wienstroer, J.; Schertl, P.; Braun, H.-P.; Schwarzländer, M.; Gonzalez, D.H.; et al. D-Lactate Dehydrogenase Links Methylglyoxal Degradation and Electron Transport through Cytochrome c. Plant Physiol. 2016, 172, 901–912. [Google Scholar] [PubMed]
- Wang, Y.; Ye, X.-Y.; Qiu, X.-M.; Li, Z.-G. Methylglyoxal Triggers the Heat Tolerance in Maize Seedlings by Driving AsA-GSH Cycle and Reactive Oxygen Species-/Methylglyoxal-Scavenging System. Plant Physiol. Biochem. 2019, 138, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Hoque, T.S.; Uraji, M.; Ye, W.; Hossain, M.A.; Nakamura, Y.; Murata, Y. Methylglyoxal-Induced Stomatal Closure Accompanied by Peroxidase-Mediated ROS Production in Arabidopsis. J. Plant Physiol. 2012, 169, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Mohsin, S.M.; Bhuyan, M.H.M.B.; Parvin, K.; Hawrylak-Nowak, B.; Fujita, M. Silicon-Induced Antioxidant Defense and Methylglyoxal Detoxification Works Coordinately in Alleviating Nickel Toxicity in Oryza sativa L. Ecotoxicology 2019, 28, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Mano, J.; Miyatake, F.; Hiraoka, E.; Tamoi, M. Evaluation of the Toxicity of Stress-Related Aldehydes to Photosynthesis in Chloroplasts. Planta 2009, 230, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Hoque, T.S.; Uraji, M.; Tuya, A.; Nakamura, Y.; Murata, Y. Methylglyoxal Inhibits Seed Germination and Root Elongation and Up-Regulates Transcription of Stress-Responsive Genes in ABA-Dependent Pathway in Arabidopsis. Plant Biol. 2012, 14, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Yiğit, İ.; Atici, Ö. Seed Priming with Nitric Oxide Mitigates Exogenous Methylglyoxal Toxicity by Restoring Glyoxalase and Antioxidant Systems in Germinating Maize (Zea mays L.) Seeds. Cereal Res. Commun. 2022, 50, 811–820. [Google Scholar] [CrossRef]
- Gambhir, P.; Raghuvanshi, U.; Parida, A.P.; Kujur, S.; Sharma, S.; Sopory, S.K.; Kumar, R.; Sharma, A.K. Elevated Methylglyoxal Levels Inhibit Tomato Fruit Ripening by Preventing Ethylene Biosynthesis. Plant Physiol. 2023, 192, 2161–2184. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Thelen, J.J. The Plastid Isoform of Triose Phosphate Isomerase Is Required for the Postgerminative Transition from Heterotrophic to Autotrophic Growth in Arabidopsis. Plant Cell 2010, 22, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Hossain, M.A.; Siddiqui, M.N.; Fujita, M.; Tran, L.-S.P. Phenotypical, Physiological and Biochemical Analyses Provide Insight into Selenium-Induced Phytotoxicity in Rice Plants. Chemosphere 2017, 178, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, J.; Kusnetsova, A.; Tsarev, A.; Janse van Rensburg, H.C.; Medvedev, S.; Demidchik, V.; Van den Ende, W.; Frolov, A. Glycation of Plant Proteins: Regulatory Roles and Interplay with Sugar Signalling? Int. J. Mol. Sci. 2019, 20, 2366. [Google Scholar] [CrossRef]
- Adak, S.; Agarwal, T.; Das, P.; Ray, S.; Lahiri Majumder, A. Characterization of Myo-Inositol Oxygenase from Rice (OsMIOX): Influence of Salinity Stress in Different Indica Rice Cultivars. Physiol. Mol. Biol. Plants 2023, 29, 927–945. [Google Scholar] [CrossRef] [PubMed]
- Lyczakowski, J.J.; Wicher, K.B.; Terrett, O.M.; Faria-Blanc, N.; Yu, X.; Brown, D.; Krogh, K.B.R.M.; Dupree, P.; Busse-Wicher, M. Removal of Glucuronic Acid from Xylan Is a Strategy to Improve the Conversion of Plant Biomass to Sugars for Bioenergy. Biotechnol. Biofuels 2017, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Lyczakowski, J.J.; Yu, L.; Terrett, O.M.; Fleischmann, C.; Temple, H.; Thorlby, G.; Sorieul, M.; Dupree, P. Two Conifer GUX Clades Are Responsible for Distinct Glucuronic Acid Patterns on Xylan. New Phytol. 2021, 231, 1720–1733. [Google Scholar] [CrossRef]
- Silveira, R.L.; Stoyanov, S.R.; Gusarov, S.; Skaf, M.S.; Kovalenko, A. Plant Biomass Recalcitrance: Effect of Hemicellulose Composition on Nanoscale Forces That Control Cell Wall Strength. J. Am. Chem. Soc. 2013, 135, 19048–19051. [Google Scholar] [CrossRef] [PubMed]
- Kanter, U.; Usadel, B.; Guerineau, F.; Li, Y.; Pauly, M.; Tenhaken, R. The Inositol Oxygenase Gene Family of Arabidopsis Is Involved in the Biosynthesis of Nucleotide Sugar Precursors for Cell-Wall Matrix Polysaccharides. Planta 2005, 221, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Wan, J.X.; Wu, Q.; Zhao, X.S.; Zheng, S.J.; Shen, R.F. PARVUS Affects Aluminium Sensitivity by Modulating the Structure of Glucuronoxylan in Arabidopsis thaliana. Plant Cell Environ. 2017, 40, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, J.; Zhao, L.; Gu, L.; Wu, F.; Tian, W.; Sun, Y.; Zhang, S.; Su, H.; Wang, L. Ectopic Expression of a Malus Hupehensis Rehd. Myo- Inositol Oxygenase Gene (MhMIOX2) Enhances Tolerance to Salt Stress. Sci. Hortic. 2021, 281, 109898. [Google Scholar] [CrossRef]
- Duan, J.; Zhang, M.; Zhang, H.; Xiong, H.; Liu, P.; Ali, J.; Li, J.; Li, Z. OsMIOX, a Myo-Inositol Oxygenase Gene, Improves Drought Tolerance through Scavenging of Reactive Oxygen Species in Rice (Oryza sativa L.). Plant Sci. 2012, 196, 143–151. [Google Scholar] [CrossRef]
- Toyofuku, K.; Kasahara, M.; Yamaguchi, J. Characterization and Expression of Monosaccharide Transporters (OsMSTs) in Rice. Plant Cell Physiol. 2000, 41, 940–947. [Google Scholar] [CrossRef]
- Sherson, S.M.; Alford, H.L.; Forbes, S.M.; Wallace, G.; Smith, S.M. Roles of Cell-wall Invertases and Monosaccharide Transporters in the Growth and Development of Arabidopsis. J. Exp. Bot. 2003, 54, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Greaves, J.G.; Jakada, B.H.; Fakher, B.; Wang, X.; Qin, Y. AcCIPK5, a Pineapple CBL-Interacting Protein Kinase, Confers Salt, Osmotic and Cold Stress Tolerance in Transgenic Arabidopsis. Plant Sci. Int. J. Exp. Plant Biol. 2022, 320, 111284. [Google Scholar] [CrossRef] [PubMed]
- Wingenter, K.; Schulz, A.; Wormit, A.; Wic, S.; Trentmann, O.; Hoermiller, I.I.; Heyer, A.G.; Marten, I.; Hedrich, R.; Neuhaus, H.E. Increased Activity of the Vacuolar Monosaccharide Transporter TMT1 Alters Cellular Sugar Partitioning, Sugar Signaling, and Seed Yield in Arabidopsis. Plant Physiol. 2010, 154, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Chigri, F.; Flosdorff, S.; Pilz, S.; Kölle, E.; Dolze, E.; Gietl, C.; Vothknecht, U.C. The Arabidopsis Calmodulin-like Proteins AtCML30 and AtCML3 Are Targeted to Mitochondria and Peroxisomes, Respectively. Plant Mol. Biol. 2012, 78, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.S.; Vadassery, J.; Heyer, M.; Reichelt, M.; Bender, K.W.; Snedden, W.A.; Boland, W.; Mithöfer, A. Mutation of the Arabidopsis Calmodulin-Like Protein CML37 Deregulates the Jasmonate Pathway and Enhances Susceptibility to Herbivory. Mol. Plant 2014, 7, 1712–1726. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Liu, Z.; Long, Y.; Liang, Y.; Jin, Z.; Zhang, L.; Liu, D.; Li, H.; Zhai, J.; Pei, Y. The Ca2+/Calmodulin2-Binding Transcription Factor TGA3 Elevates LCD Expression and H2S Production to Bolster Cr6+ Tolerance in Arabidopsis. Plant J. 2017, 91, 1038–1050. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Zarinkamar, F. Nitric Oxide, Calmodulin and Calcium Protein Kinase Interactions in the Response of Brassica Napus to Salinity Stress. Plant Biol. 2023, 25, 411–419. [Google Scholar] [CrossRef]
- Tian, W.; Hou, C.; Ren, Z.; Wang, C.; Zhao, F.; Dahlbeck, D.; Hu, S.; Zhang, L.; Niu, Q.; Li, L.; et al. A Calmodulin-Gated Calcium Channel Links Pathogen Patterns to Plant Immunity. Nature 2019, 572, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.M.; Huang, L.F.; Zhang, X.; Wang, M.L.; Xu, G.Y.; Xia, X.J. OsCML4 Improves Drought Tolerance through Scavenging of Reactive Oxygen Species in Rice. J. Plant Biol. 2015, 58, 68–73. [Google Scholar] [CrossRef]
- Yoo, J.H.; Cheong, M.S.; Park, C.Y.; Moon, B.C.; Kim, M.C.; Kang, Y.H.; Park, H.C.; Choi, M.S.; Lee, J.H.; Jung, W.Y.; et al. Regulation of the Dual Specificity Protein Phosphatase, DsPTP1, through Interactions with Calmodulin*. J. Biol. Chem. 2004, 279, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Delumeau, O.; Paven, M.-C.M.-L.; Montrichard, F.; Laval-Martin, D.L. Effects of Short-Term NaCl Stress on Calmodulin Transcript Levels and Calmodulin-Dependent NAD Kinase Activity in Two Species of Tomato. Plant Cell Environ. 2000, 23, 329–336. [Google Scholar] [CrossRef]
- Ghorbel, M.; Zaidi, I.; Robe, E.; Ranty, B.; Mazars, C.; Galaud, J.-P.; Hanin, M. The Activity of the Wheat MAP Kinase Phosphatase 1 Is Regulated by Manganese and by Calmodulin. Biochimie 2015, 108, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Borsics, T.; Webb, D.; Andeme-Ondzighi, C.; Staehelin, L.A.; Christopher, D.A. The Cyclic Nucleotide-Gated Calmodulin-Binding Channel AtCNGC10 Localizes to the Plasma Membrane and Influences Numerous Growth Responses and Starch Accumulation in Arabidopsis Thaliana. Planta 2007, 225, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ji, L.; Liu, S.; Jing, P.; Hu, J.; Jin, D.; Wang, L.; Xie, G. The CaM1-Associated CCaMK–MKK1/6 Cascade Positively Affects Lateral Root Growth via Auxin Signaling under Salt Stress in Rice. J. Exp. Bot. 2021, 72, 6611–6627. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Koo, Y.D.; Lee, J.; Lee, H.; Baek, D.; Lee, S.; Cheon, C.-I.; Kwak, S.-S.; Lee, S.Y.; Yun, D.-J. Athb-12, a Homeobox-Leucine Zipper Domain Protein from Arabidopsis Thaliana, Increases Salt Tolerance in Yeast by Regulating Sodium Exclusion. Biochem. Biophys. Res. Commun. 2004, 323, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Arazi, T.; Sunkar, R.; Kaplan, B.; Fromm, H. A Tobacco Plasma Membrane Calmodulin-Binding Transporter Confers Ni2+ Tolerance and Pb2+ Hypersensitivity in Transgenic Plants. Plant J. 1999, 20, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Moy, A.; Czajka, K.; Michael, P.; Nkongolo, K. Transcriptome Analysis Reveals Changes in Whole Gene Expression, Biological Process, and Molecular Functions Induced by Nickel in Jack Pine (Pinus banksiana). Plants 2023, 12, 2889. [Google Scholar] [CrossRef]








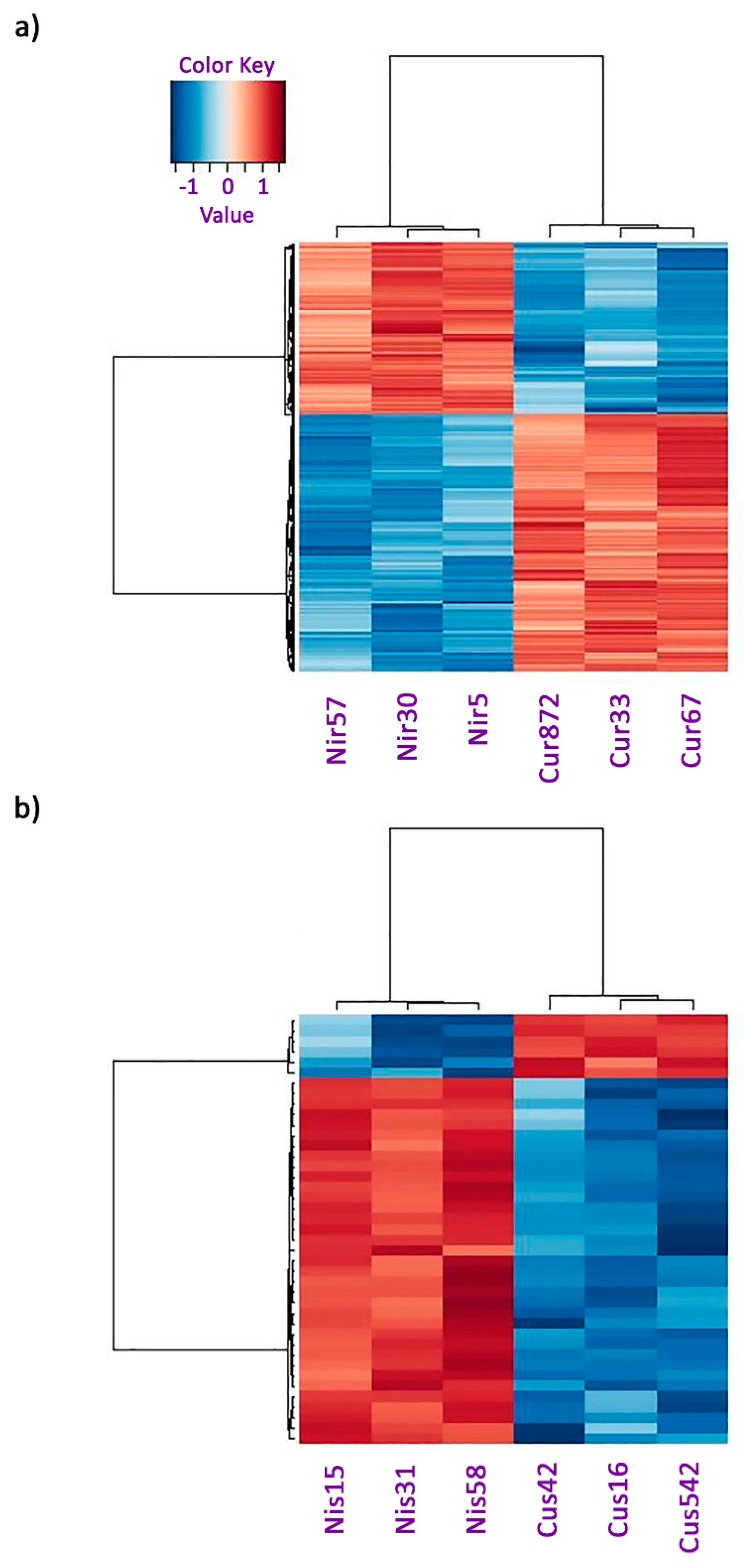{kind=link}
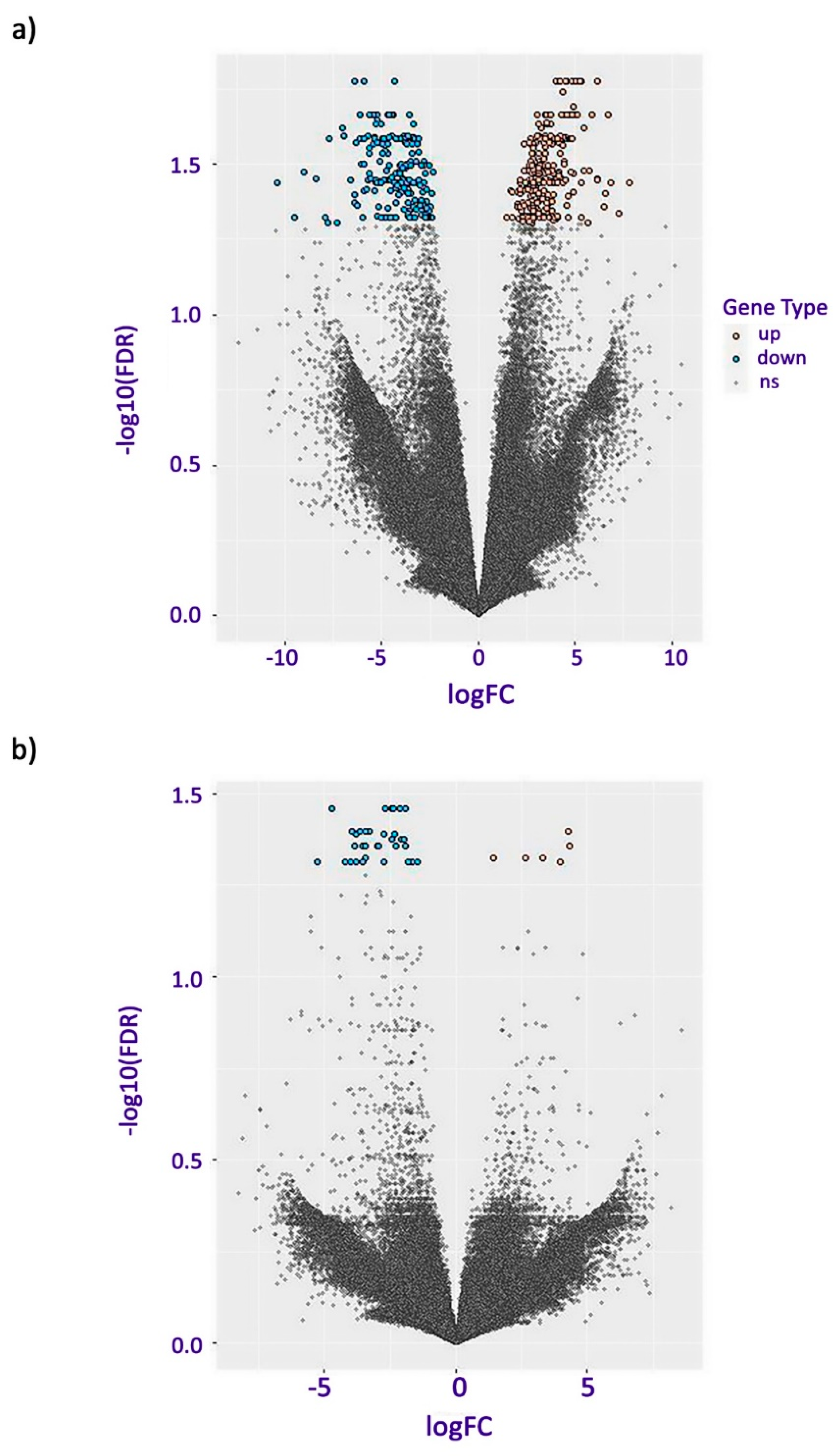{kind=link}
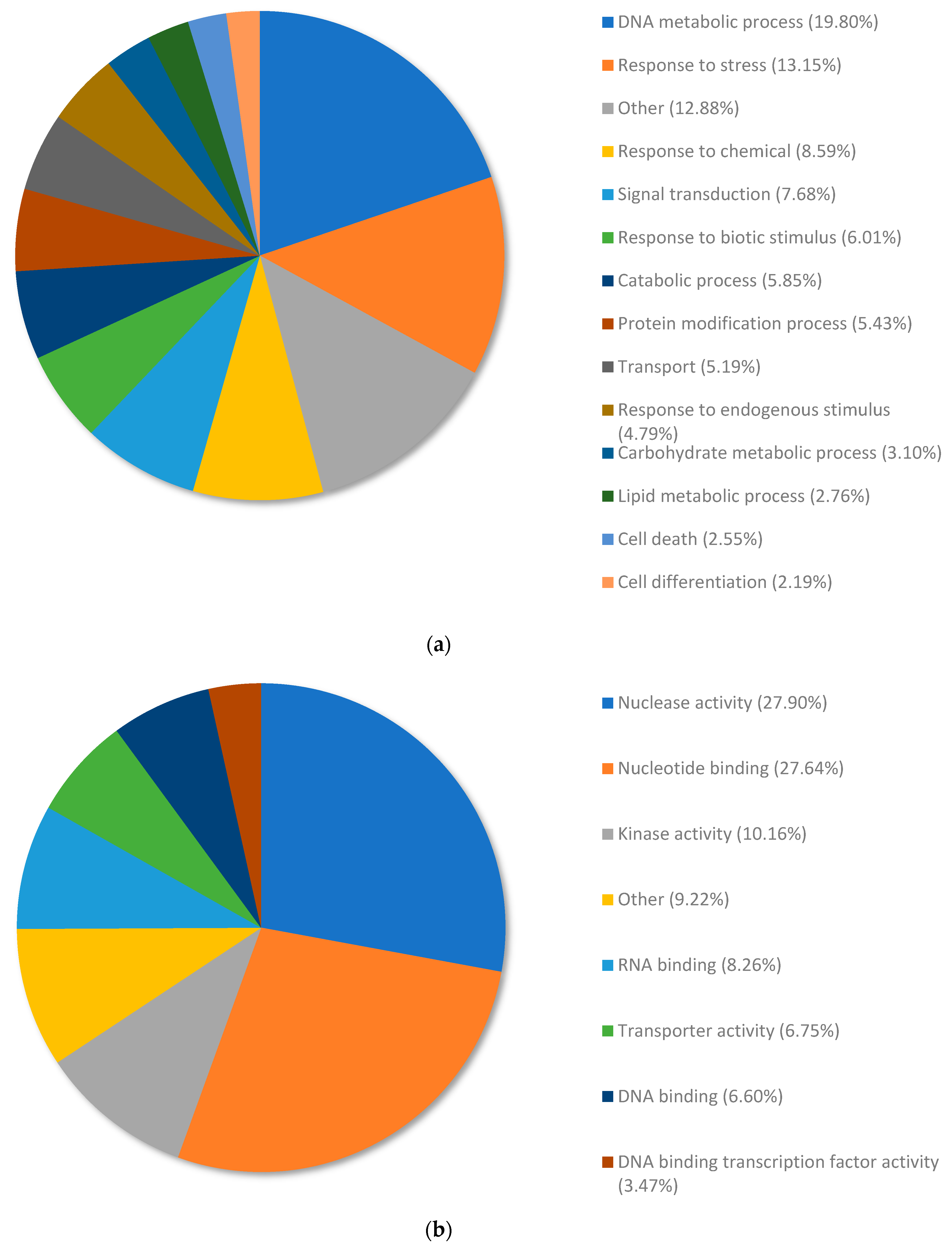{kind=link}
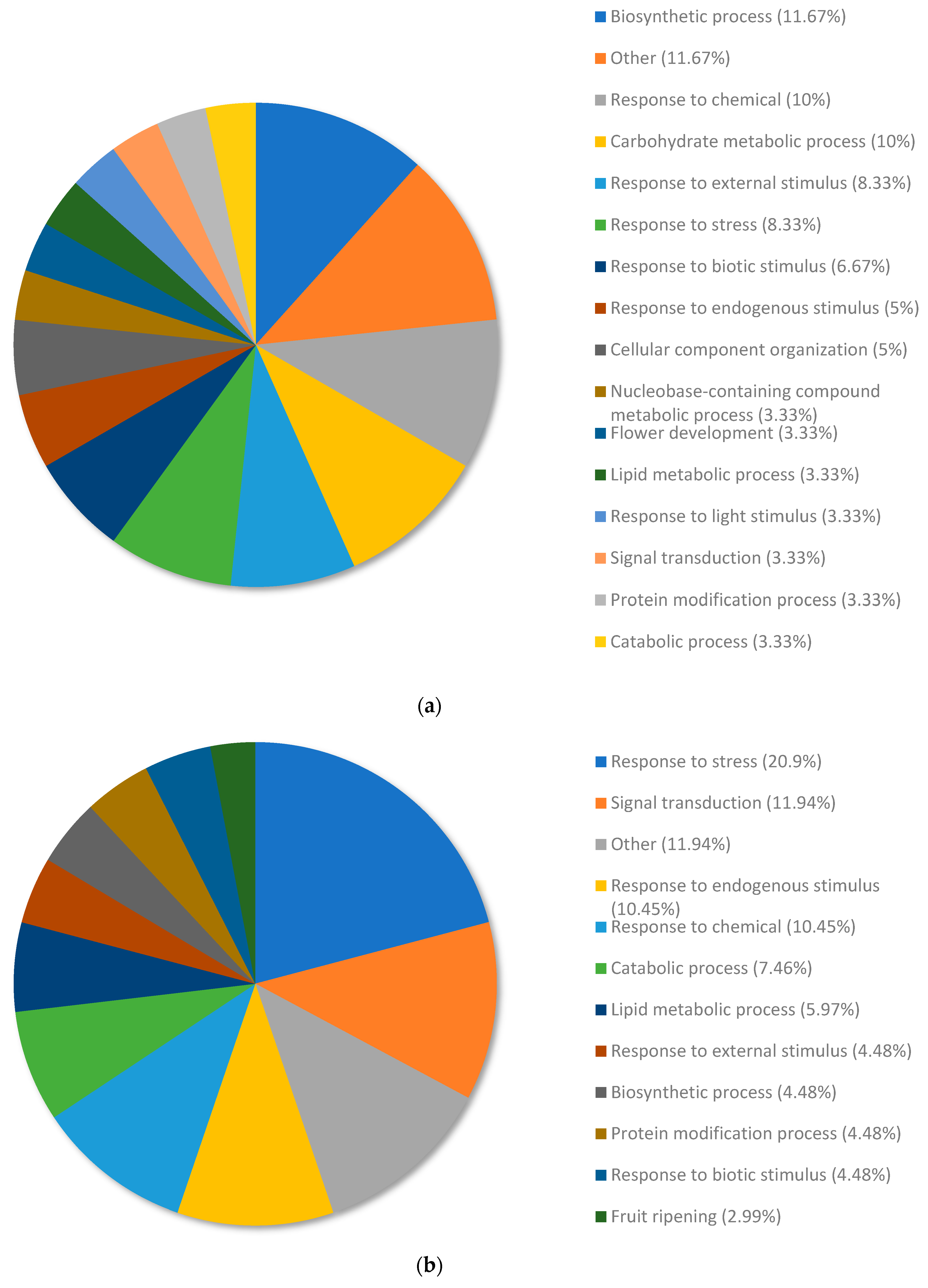{kind=link}
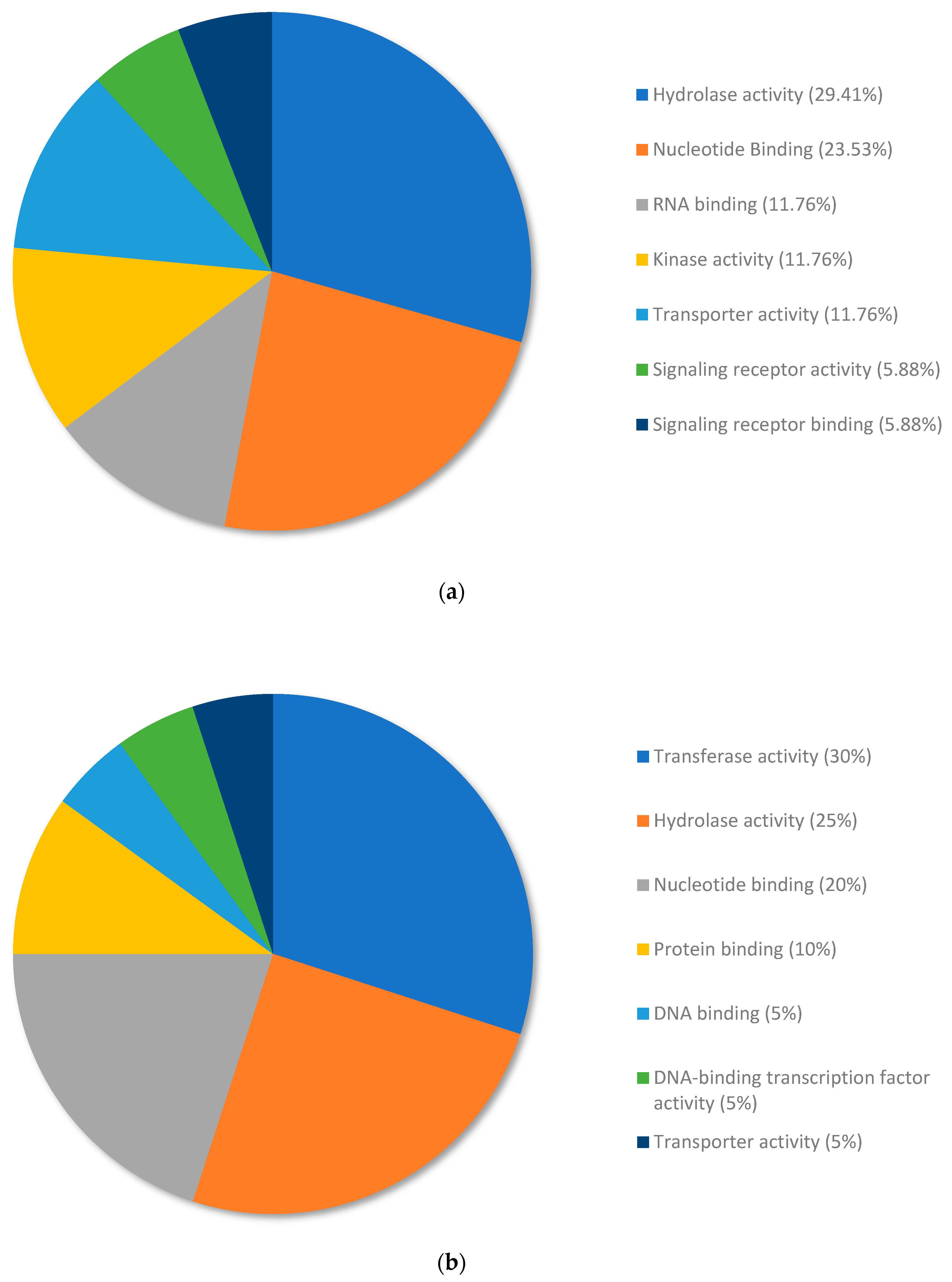{kind=link}
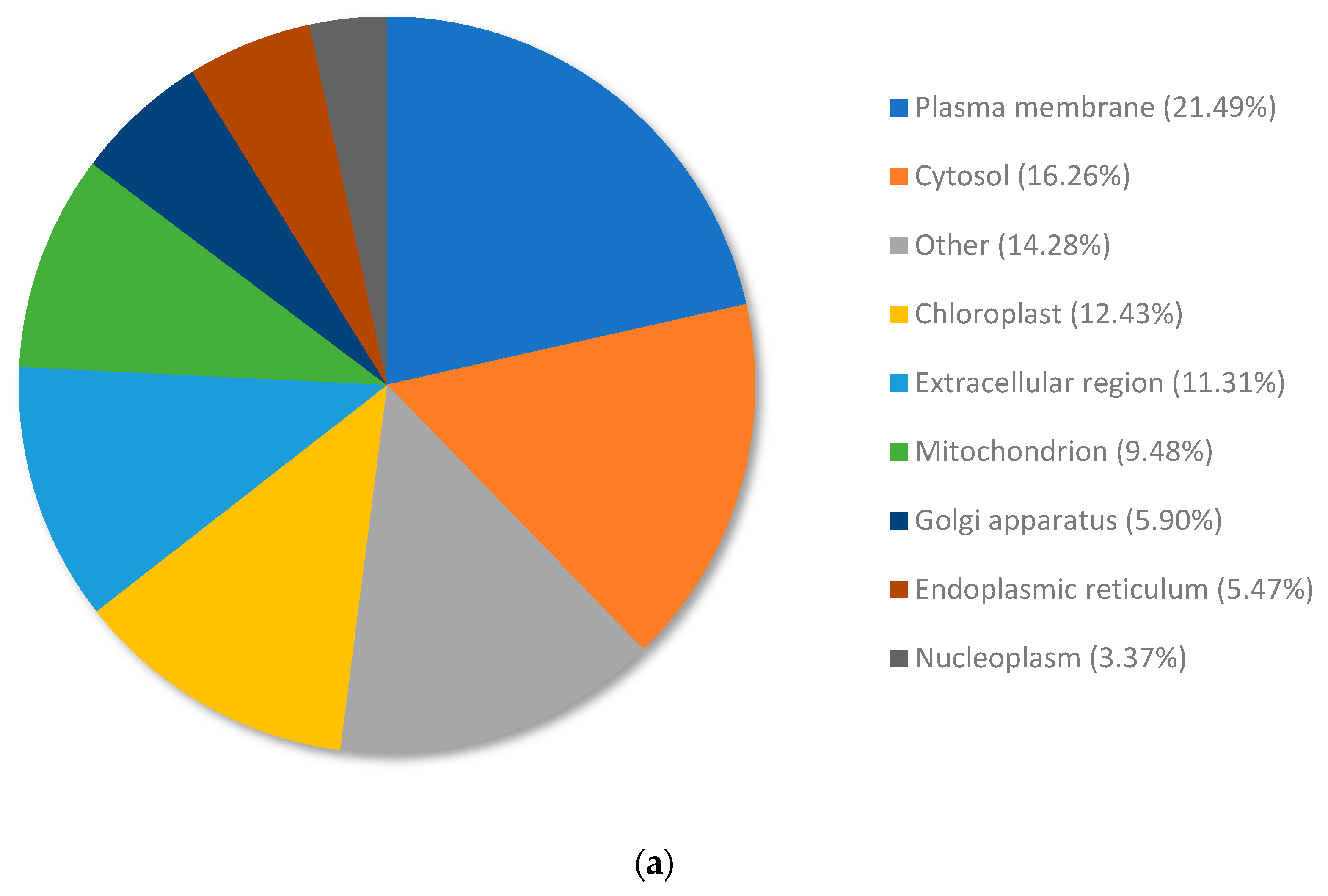{kind=link}
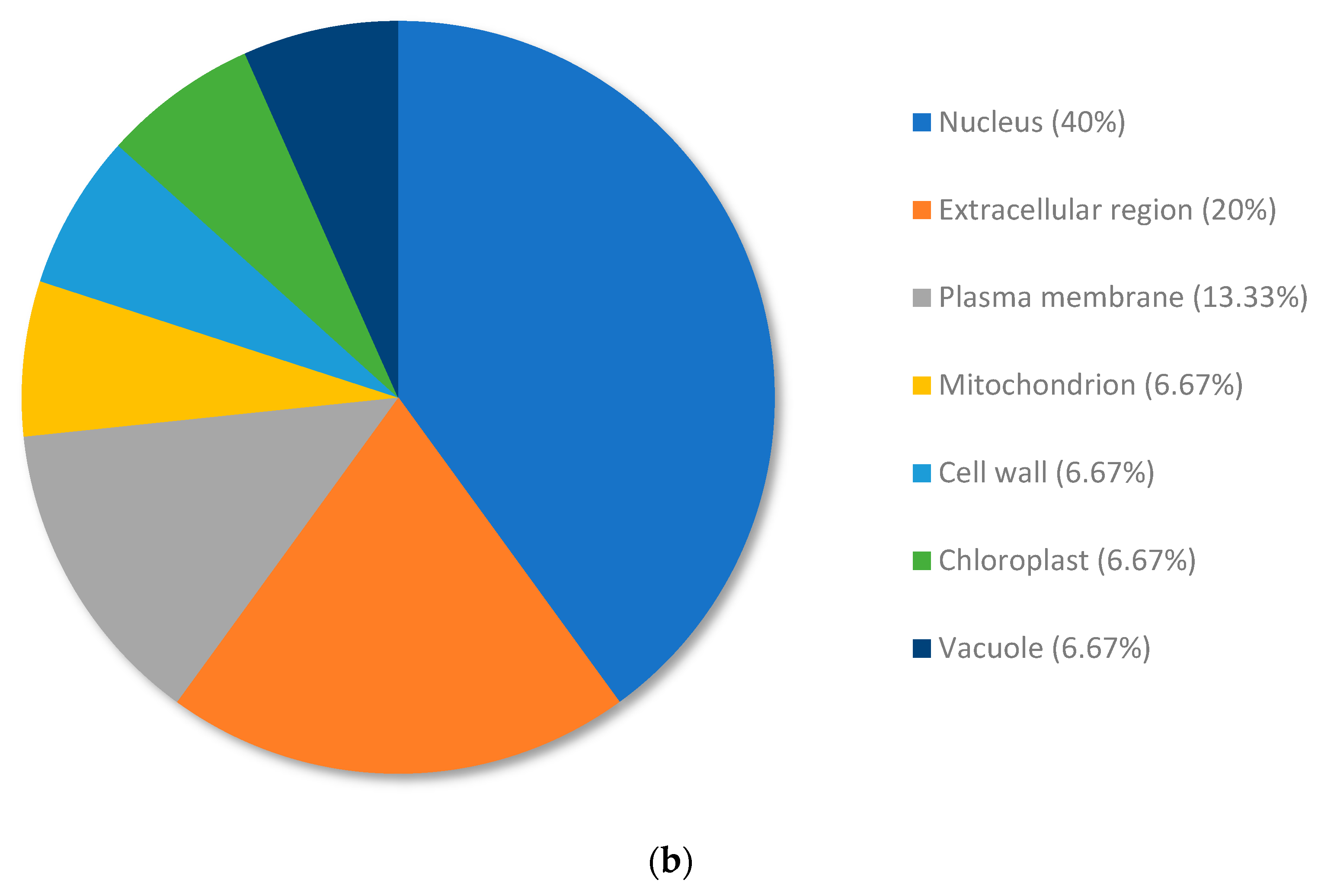{kind=link}
| Cut-Off | Standard (Two Fold and FDR 0.05) | Low Stringency (Two Fold and p Value 0.01) |
|---|---|---|
| Upregulated genes | 269 | 3387 |
| Downregulated genes | 180 | 3815 |
| Total genes | 449 | 7202 |
| Cut-Off | Standard (Two Fold and FDR 0.05) | Low Stringency (Two Fold and p Value 0.01) |
|---|---|---|
| Upregulated genes | 6 | 2494 |
| Downregulated genes | 35 | 2626 |
| Total genes | 41 | 5120 |
| Rank | Gene ID | Cu Res1 | Cu Res2 | Cu Res3 | Ni Res1 | Ni Res2 | Ni Res3 | logFC | Adj. p Value | UniProt Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TRINITY_DN2085_c0_g1 | 29.21 | 82.28 | 46.17 | 0.34 | 0.57 | 0.14 | 7.793 | 7.10E-05 | Predicted Protein |
| 1 | TRINITY_DN1891_c0_g2 | 51.01 | 52.45 | 17.61 | 0.04 | 2.06 | 0.7 | 7.247 | 0.000114 | Predicted Protein |
| 2 | TRINITY_DN8469_c0_g1 | 43.02 | 275.4 | 201.9 | 1.33 | 8.3 | 0.46 | 6.863 | 6.98E-05 | Acid phosphatase 1, EC 3.1.3.2 (Apase-1(1)) |
| 3 | TRINITY_DN1979_c0_g1 | 32.13 | 43.51 | 47.01 | 0.21 | 3.16 | 0.33 | 6.702 | 5.45E-06 | Predicted Protein |
| 4 | TRINITY_DN4373_c0_g1 | 35.55 | 56.98 | 13.12 | 0.54 | 0.5 | 0.36 | 6.595 | 8.44E-05 | Predicted Protein |
| 5 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106363 | Pleiotropic drug resistance protein 1 (NtPDR1) |
| 6 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | ABC transporter G family member 32, ABC transporter ABCG.32, AtABCG32 (Pleiotropic drug resistance protein 4) (Protein PERMEABLE CUTICLE 1) |
| 7 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | ABC transporter G family member 36, OsABCG36 (Pleiotropic drug resistance protein 9, OsPDR9) |
| 8 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | Pleiotropic drug resistance protein 1 (NpPDR1) |
| 9 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | ABC transporter G family member 44, OsABCG44 (Pleiotropic drug resistance protein 17, OsPDR17) |
| 10 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | ABC transporter G family member 31, OsABCG31 (Pleiotropic drug resistance protein 6, OsPDR6) |
| 11 | TRINITY_DN5_c0_g1 | 18.7 | 16.84 | 12.27 | 0.1 | 0.8 | 0.23 | 6.538 | 0.000106 | ABC transporter G family member 34, OsABCG34 (Pleiotropic drug resistance protein 10, OsPDR10) |
| 12 | TRINITY_DN5742_c0_g1 | 25.47 | 76.4 | 14.17 | 0.6 | 1.07 | 0.41 | 6.162 | 5.17E-05 | Predicted Protein |
| 13 | TRINITY_DN59895_c0_g1 | 107.34 | 136.22 | 54.47 | 0.8 | 4.56 | 2.36 | 6.155 | 1.45E-06 | Predicted Protein |
| 14 | TRINITY_DN51306_c0_g1 | 32.05 | 61.43 | 35.69 | 0.53 | 0.7 | 1.91 | 6.153 | 1.48E-06 | Predicted Protein |
| 15 | TRINITY_DN8170_c0_g1 | 50.21 | 123.33 | 300.25 | 1.81 | 1.13 | 8.48 | 6.150 | 5.72E-05 | Predicted Protein |
| 16 | TRINITY_DN19606_c0_g1 | 31.77 | 21.11 | 20.22 | 0.5 | 1.66 | 0.29 | 5.866 | 4.22E-06 | Granule-bound starch synthase 1, chloroplastic/amyloplastic, EC 2.4.1.242 (Granule-bound starch synthase I, GBSS-I) |
| 17 | TRINITY_DN6541_c0_g1 | 251.91 | 62.08 | 39.98 | 3.22 | 2.8 | 1.68 | 5.744 | 4.07E-05 | Heavy metal-associated isoprenylated plant protein 20, AtHIP20, AtHIPP20 |
| 18 | TRINITY_DN6541_c0_g1 | 251.91 | 62.08 | 39.98 | 3.22 | 2.8 | 1.68 | 5.744 | 4.07E-05 | Heavy metal-associated isoprenylated plant protein 26, AtHIP26, AtHIPP26 (Farnesylated protein 6, AtFP6) |
| 19 | TRINITY_DN47729_c0_g2 | 7.89 | 24.87 | 20.63 | 0.46 | 0.23 | 0.87 | 5.668 | 0.000146 | Predicted Protein |
| 20 | TRINITY_DN430_c0_g1 | 123.72 | 65.04 | 52.06 | 1.02 | 8.95 | 2.15 | 5.441 | 8.58E-06 | Cytochrome P450 86A22, EC 1.14.14.129 (Long-chain acyl-CoA omega-monooxygenase) |
| 21 | TRINITY_DN1790_c0_g1 | 9.55 | 18.4 | 18.66 | 0.23 | 2.08 | 0.31 | 5.440 | 4.00E-05 | Receptor-like protein kinase HSL1, EC 2.7.11.1 (Protein HAESA-LIKE1) |
| 22 | TRINITY_DN1072_c0_g1 | 97.21 | 115.92 | 41.8 | 4.4 | 9.29 | 0.62 | 5.334 | 6.56E-05 | Predicted Protein |
| 23 | TRINITY_DN6055_c0_g1 | 18.79 | 22.98 | 21.77 | 0.56 | 1.18 | 0.81 | 5.330 | 5.78E-07 | Predicted Protein |
| 24 | TRINITY_DN6813_c0_g1 | 235.35 | 117.07 | 35.45 | 1.7 | 10.19 | 3.65 | 5.309 | 6.74E-05 | Predicted Protein |
| 25 | TRINITY_DN11710_c0_g2 | 150.88 | 92.2 | 20.39 | 1.17 | 7.66 | 2.13 | 5.304 | 0.000137 | Predicted Protein |
| 26 | TRINITY_DN63319_c0_g1 | 28.39 | 21.29 | 22.62 | 0.58 | 1.05 | 1.41 | 5.255 | 8.92E-07 | Predicted Protein |
| 27 | TRINITY_DN82353_c0_g1 | 13.94 | 23.06 | 17.36 | 0.87 | 0.98 | 0.45 | 5.156 | 2.89E-06 | Predicted Protein |
| 28 | TRINITY_DN48473_c0_g1 | 56.5 | 107.55 | 80.8 | 1.68 | 5.59 | 4.91 | 5.085 | 9.27E-07 | Predicted Protein |
| 29 | TRINITY_DN2130_c0_g1 | 22.32 | 44.82 | 44.69 | 2.2 | 1.09 | 1.68 | 5.050 | 4.23E-06 | Predicted Protein |
| 30 | TRINITY_DN1441_c0_g2 | 340.37 | 453.06 | 121.8 | 13.03 | 19.89 | 9.5 | 4.943 | 5.01E-06 | Predicted Protein |
| Rank | Gene ID | Cu Res1 | Cu Res2 | Cu Res3 | Ni Res1 | Ni Res2 | Ni Res3 | logFC | Adj. p Value | UniProt Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TRINITY_DN43547_c0_g1 | 0 | 0 | 0 | 24.15 | 39.01 | 23.93 | −10.382 | 6.42E-05 | Predicted Protein |
| 1 | TRINITY_DN1456_c0_g1 | 1.37 | 0 | 1.34 | 306.48 | 253.1 | 155.47 | −9.527 | 0.000128719 | Predicted Protein |
| 2 | TRINITY_DN2126_c0_g1 | 1.31 | 0.24 | 0.4 | 954.55 | 193.03 | 448.85 | −9.030 | 4.26E-05 | UDP-glycosyltransferase 75C1, Abscisic acid beta-glucosyltransferase, Indole-3-acetate beta-glucosyltransferase, SlUGT75C1, EC 2.4.1.121, EC 2.4.1.263 |
| 3 | TRINITY_DN5965_c0_g1 | 0.28 | 0.12 | 1.02 | 159.21 | 70.92 | 203.21 | −8.429 | 5.08E-05 | Predicted Protein |
| 4 | TRINITY_DN5240_c1_g1 | 0.06 | 0.2 | 0.38 | 75.64 | 57.88 | 47.63 | −7.894 | 0.000131675 | Predicted Protein |
| 5 | TRINITY_DN2786_c0_g1 | 7.97 | 1.28 | 0.09 | 767.81 | 197.57 | 545.86 | −7.817 | 0.000147002 | Predicted Protein |
| 6 | TRINITY_DN7685_c0_g1 | 0.35 | 0.17 | 0.18 | 69.15 | 75.78 | 88.19 | −7.718 | 1.68E-05 | Predicted Protein |
| 7 | TRINITY_DN3685_c0_g2 | 7.54 | 1.2 | 0.09 | 524.13 | 169.45 | 298.36 | −7.315 | 0.000146393 | Copia protein (Gag-int-pol protein) [Cleaved into: Copia VLP protein; Copia protease, EC 3.4.23.-] |
| 8 | TRINITY_DN2463_c0_g1 | 5.21 | 0.97 | 0.68 | 301.76 | 196.16 | 568.86 | −7.021 | 8.44E-06 | Predicted Protein |
| 9 | TRINITY_DN2040_c0_g1 | 28.51 | 6 | 12.87 | 3943.91 | 1043.09 | 3644.25 | −6.950 | 1.04E-05 | Predicted Protein |
| 10 | TRINITY_DN5795_c0_g1 | 13.6 | 5.09 | 0.4 | 753.52 | 420.03 | 412.9 | −6.580 | 5.36E-05 | Predicted Protein |
| 11 | TRINITY_DN735_c0_g1 | 5.88 | 1.92 | 0.26 | 401.38 | 124.53 | 198.21 | −6.422 | 9.90E-05 | Predicted Protein |
| 12 | TRINITY_DN41085_c0_g1 | 1.46 | 0.55 | 8.2 | 230.93 | 100.32 | 461.71 | −6.396 | 8.67E-05 | Predicted Protein |
| 13 | TRINITY_DN5240_c0_g1 | 0.4 | 0.48 | 1.48 | 86.62 | 78.96 | 59.64 | −6.375 | 1.11E-06 | Protein TIFY 10b, OsTIFY10b (Jasmonate ZIM domain-containing protein 7, OsJAZ7) (OsJAZ6) |
| 14 | TRINITY_DN17_c0_g2 | 0.65 | 0.15 | 0.58 | 50.63 | 58.32 | 26.97 | −6.307 | 1.89E-05 | RING-H2 finger protein ATL60, EC 2.3.2.27 (RING-type E3 ubiquitin transferase ATL60) |
| 15 | TRINITY_DN800_c1_g1 | 2.53 | 0.34 | 0.33 | 127.84 | 44.55 | 121.22 | −6.264 | 0.000103509 | Predicted Protein |
| 16 | TRINITY_DN1755_c0_g1 | 0.73 | 0.19 | 0.22 | 37.36 | 42.04 | 37.53 | −6.143 | 1.54E-05 | Predicted Protein |
| 17 | TRINITY_DN1067_c0_g1 | 0.21 | 0.63 | 2.66 | 74.75 | 51.13 | 62.5 | −6.138 | 1.50E-05 | Predicted Protein |
| 18 | TRINITY_DN4963_c0_g1 | 1.2 | 1.08 | 0.26 | 107.91 | 63.25 | 101.25 | −6.114 | 6.05E-06 | Predicted Protein |
| 19 | TRINITY_DN3889_c0_g1 | 8.77 | 0.85 | 2.51 | 414.11 | 167.54 | 300.23 | −6.071 | 3.22E-05 | Predicted Protein |
| 20 | TRINITY_DN6289_c0_g1 | 0.34 | 0.08 | 0.38 | 23.79 | 21.01 | 21.41 | −5.988 | 0.000135072 | Predicted Protein |
| 21 | TRINITY_DN6111_c0_g1 | 0.47 | 0.11 | 0.98 | 29.31 | 29.12 | 29.02 | −5.932 | 1.49E-05 | Predicted Protein |
| 22 | TRINITY_DN4477_c0_g1 | 1.32 | 1.08 | 0.77 | 67.92 | 163.48 | 92.08 | −5.924 | 1.74E-07 | Predicted Protein |
| 23 | TRINITY_DN71807_c0_g1 | 0.75 | 0.09 | 1.03 | 29.61 | 23.8 | 49.04 | −5.903 | 8.02E-05 | Predicted Protein |
| 24 | TRINITY_DN971_c0_g1 | 0.32 | 0.18 | 3.12 | 40.71 | 30.9 | 35.98 | −5.891 | 3.15E-05 | 1-aminocyclopropane-1-carboxylate synthase 7, ACC synthase 7, EC 4.4.1.14 (S-adenosyl-L-methionine methylthioadenosine-lyase 7) |
| 25 | TRINITY_DN971_c0_g1 | 0.32 | 0.18 | 3.12 | 40.71 | 30.9 | 35.98 | −5.891 | 3.15E-05 | 1-aminocyclopropane-1-carboxylate synthase, ACC synthase, EC 1.4.-.-, EC 4.4.1.14 (S-adenosyl-L-methionine methylthioadenosine-lyase) |
| 26 | TRINITY_DN971_c0_g1 | 0.32 | 0.18 | 3.12 | 40.71 | 30.9 | 35.98 | −5.891 | 3.15E-05 | 1-aminocyclopropane-1-carboxylate synthase 6, ACC synthase 6, EC 4.4.1.14 (S-adenosyl-L-methionine methylthioadenosine-lyase 6) |
| 27 | TRINITY_DN971_c0_g1 | 0.32 | 0.18 | 3.12 | 40.71 | 30.9 | 35.98 | −5.891 | 3.15E-05 | 1-aminocyclopropane-1-carboxylate synthase 3, ACC synthase 3, EC 4.4.1.14 (Le-ACS3, ACS-3) (S-adenosyl-L-methionine methylthioadenosine-lyase 3) |
| 28 | TRINITY_DN971_c0_g1 | 0.32 | 0.18 | 3.12 | 40.71 | 30.9 | 35.98 | −5.891 | 3.15E-05 | 1-aminocyclopropane-1-carboxylate synthase 1, ACC synthase 1, EC 4.4.1.14 (S-adenosyl-L-methionine methylthioadenosine-lyase) |
| 29 | TRINITY_DN756_c0_g1 | 7.06 | 0.88 | 8.25 | 578.57 | 195.91 | 235.86 | −5.769 | 7.82E-05 | Dirigent protein 21, AtDIR21 |
| 30 | TRINITY_DN6061_c0_g1 | 3.75 | 2.9 | 7.44 | 355.22 | 178.84 | 637.66 | −5.730 | 1.01E-05 | Predicted Protein |
| Rank | Gene ID | Cu Sus 1 | Cu Sus 2 | Cu Sus 3 | Ni Sus 1 | Ni Sus 2 | Ni Sus 3 | logFC | Adj. p Value | UniProt Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TRINITY_DN3441_c0_g1 | 161.51 | 141.19 | 99.42 | 14.73 | 4.54 | 4.16 | 4.327038 | 4.74E-06 | Predicted Protein |
| 1 | TRINITY_DN29629_c0_g1 | 130.48 | 102.59 | 87.52 | 8.98 | 4.39 | 3.79 | 4.292084 | 1.72E-06 | Predicted Protein |
| 2 | TRINITY_DN25822_c0_g1 | 52.29 | 67.97 | 55.74 | 7.15 | 2.61 | 2.46 | 3.959541 | 7.45E-06 | Sugar transport protein MST3 (Monosaccharide transporter 3, OsMST3) (Sugar:proton symporter MST3) |
| 3 | TRINITY_DN324_c0_g1 | 122.98 | 261.98 | 186.02 | 18.74 | 13.27 | 19.44 | 3.330385 | 5.86E-06 | Inositol oxygenase 1, EC 1.13.99.1 (Myo-inositol oxygenase 1, AtMIOX1, MI oxygenase 1) |
| 4 | TRINITY_DN324_c0_g1 | 122.98 | 261.98 | 186.02 | 18.74 | 13.27 | 19.44 | 3.330385 | 5.86E-06 | Inositol oxygenase 5, EC 1.13.99.1 (Myo-inositol oxygenase 5, AtMIOX5, MI oxygenase 5) |
| 5 | TRINITY_DN324_c0_g1 | 122.98 | 261.98 | 186.02 | 18.74 | 13.27 | 19.44 | 3.330385 | 5.86E-06 | Probable inositol oxygenase, EC 1.13.99.1 (Myo-inositol oxygenase, MI oxygenase) |
| 6 | TRINITY_DN324_c0_g1 | 122.98 | 261.98 | 186.02 | 18.74 | 13.27 | 19.44 | 3.330385 | 5.86E-06 | Inositol oxygenase 4, EC 1.13.99.1 (Myo-inositol oxygenase 4, AtMIOX4, MI oxygenase 4) |
| 7 | TRINITY_DN13749_c0_g1 | 87.25 | 112.1 | 72.4 | 18.01 | 11.02 | 12.71 | 2.669987 | 5.62E-06 | Predicted Protein |
| 8 | TRINITY_DN1743_c0_g1 | 398.17 | 530.25 | 356.13 | 126.44 | 202.71 | 141.62 | 1.416768 | 5.38E-06 | G-type lectin S-receptor-like serine/threonine-protein kinase At5g24080, EC 2.7.11.1 |
| Rank | Gene ID | Cu Sus1 | Cu Sus2 | Cu Sus3 | Ni Sus1 | Ni Sus2 | Ni Sus3 | logFC | Adj. p Value | UniProt Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TRINITY_DN343_c0_g1 | 1.96 | 1.42 | 0.92 | 54.19 | 45.61 | 75.54 | −5.26455 | 7.48E-06 | Probable disease resistance protein At4g33300 |
| 1 | TRINITY_DN1105_c0_g1 | 3.27 | 4.87 | 2.07 | 76.5 | 72.45 | 109.99 | −4.74021 | 8.13E-07 | Probable calcium-binding protein CML24 (Calmodulin-like protein 24) |
| 2 | TRINITY_DN1140_c1_g1 | 4.64 | 8.22 | 3.13 | 95.64 | 64.13 | 116.13 | −4.21256 | 7.05E-06 | Heavy metal-associated isoprenylated plant protein 39, AtHIP39 |
| 3 | TRINITY_DN9069_c0_g1 | 3.08 | 3.94 | 1.43 | 39 | 35.11 | 55.72 | −4.0072 | 7.49E-06 | Predicted Protein |
| 4 | TRINITY_DN26455_c0_g1 | 5.55 | 5.56 | 3.67 | 47.82 | 81.22 | 105.73 | −3.96197 | 1.86E-06 | Predicted Protein |
| 5 | TRINITY_DN709_c0_g1 | 3.48 | 4.83 | 1.97 | 36.9 | 41.46 | 63.36 | −3.84112 | 4.28E-06 | Predicted Protein |
| 6 | TRINITY_DN24318_c0_g1 | 4.75 | 2.79 | 4.67 | 40.3 | 50.05 | 80.73 | −3.79817 | 8.01E-06 | Predicted Protein |
| 7 | TRINITY_DN2118_c0_g1 | 5.31 | 19.73 | 5.94 | 91.04 | 111.37 | 142.16 | −3.79689 | 2.20E-06 | Predicted Protein |
| 8 | TRINITY_DN66111_c1_g1 | 14.78 | 24.19 | 24.59 | 172.94 | 234.1 | 371.3 | −3.66779 | 1.60E-06 | Predicted Protein |
| 9 | TRINITY_DN9317_c0_g1 | 9.85 | 29.45 | 5.37 | 142.89 | 125.63 | 137.64 | −3.56924 | 7.93E-06 | Predicted Protein |
| 10 | TRINITY_DN25789_c0_g1 | 5.35 | 15.72 | 3.93 | 67.56 | 76.93 | 92.73 | −3.54421 | 3.52E-06 | Predicted Protein |
| 11 | TRINITY_DN2751_c0_g1 | 12.73 | 8.43 | 4.46 | 77.66 | 76.97 | 116.69 | −3.4682 | 4.60E-06 | Predicted Protein |
| 12 | TRINITY_DN1481_c0_g1 | 31.61 | 46.43 | 12.53 | 248.24 | 432.11 | 226.95 | −3.44461 | 5.91E-06 | Predicted Protein |
| 13 | TRINITY_DN37281_c0_g1 | 6.17 | 13.07 | 6.21 | 74.35 | 73.03 | 101.82 | −3.42972 | 1.93E-06 | Predicted Protein |
| 14 | TRINITY_DN11462_c0_g1 | 15.98 | 31.57 | 16.82 | 122.24 | 278.4 | 208.28 | −3.29128 | 1.68E-06 | Predicted Protein |
| 15 | TRINITY_DN3306_c0_g2 | 18.25 | 34.56 | 13.19 | 154.79 | 169.1 | 263.27 | −3.27608 | 1.59E-06 | Calcium-binding protein KIC (KCBP-interacting calcium-binding protein) |
| 16 | TRINITY_DN4522_c0_g1 | 12.58 | 21.21 | 9.5 | 81.86 | 98.59 | 152.54 | −3.01301 | 3.71E-06 | Predicted Protein |
| 17 | TRINITY_DN729_c0_g1 | 85.61 | 37.04 | 35.87 | 381.71 | 394.83 | 348.76 | −2.92258 | 4.38E-06 | Alcohol dehydrogenase 2, EC 1.1.1.1 |
| 18 | TRINITY_DN729_c0_g1 | 85.61 | 37.04 | 35.87 | 381.71 | 394.83 | 348.76 | −2.92258 | 4.38E-06 | Alcohol dehydrogenase 1, EC 1.1.1.1 (ADH slow-allele) |
| 19 | TRINITY_DN41085_c0_g1 | 50.47 | 50.02 | 52.9 | 257.45 | 297.21 | 485.74 | −2.76064 | 2.45E-06 | Predicted Protein |
| 20 | TRINITY_DN3066_c1_g1 | 13.73 | 15.11 | 13.46 | 55.99 | 116.42 | 118.98 | −2.74976 | 6.97E-06 | Predicted Protein |
| 21 | TRINITY_DN2515_c0_g1 | 42.66 | 64 | 22.61 | 203.34 | 269.87 | 276.58 | −2.66249 | 6.45E-07 | Probable disease resistance protein At5g04720 |
| 22 | TRINITY_DN1690_c0_g1 | 82.49 | 125.29 | 74.33 | 334.26 | 557.98 | 584.67 | −2.43231 | 2.73E-07 | Predicted Protein |
| 23 | TRINITY_DN1454_c0_g1 | 36.11 | 69.11 | 23.97 | 190.96 | 205.3 | 220.09 | −2.42405 | 3.20E-06 | Probable disease resistance protein At4g33300 |
| 24 | TRINITY_DN1454_c0_g1 | 36.11 | 69.11 | 23.97 | 190.96 | 205.3 | 220.09 | −2.42405 | 3.20E-06 | Disease resistance protein ADR1 (Activated disease resistance protein 1) |
| 25 | TRINITY_DN187_c0_g2 | 34.07 | 59.55 | 27.12 | 157.12 | 214.46 | 204.87 | −2.36432 | 8.35E-07 | Probable disease resistance protein At4g33300 |
| 26 | TRINITY_DN187_c0_g2 | 34.07 | 59.55 | 27.12 | 157.12 | 214.46 | 204.87 | −2.36432 | 8.35E-07 | Probable RNA-binding protein 19 (RNA-binding motif protein 19) |
| 27 | TRINITY_DN2581_c0_g1 | 73.03 | 104.45 | 74.62 | 307.26 | 360.65 | 549.82 | −2.30639 | 2.39E-06 | Ubiquitin-40S ribosomal protein S27a-1 [Cleaved into: Ubiquitin; 40S ribosomal protein S27a-1 ] |
| 28 | TRINITY_DN829_c0_g1 | 36.79 | 52.72 | 34.25 | 136.33 | 187.57 | 265.96 | −2.27764 | 4.73E-06 | Predicted Protein |
| 29 | TRINITY_DN871_c0_g1 | 77.17 | 92.43 | 46.57 | 254.71 | 317.44 | 342.34 | −2.14567 | 5.09E-07 | Lipase-like PAD4, EC 2.3.1.- (Protein ENHANCED DISEASE SUSCEPTIBILITY 9) (Protein PHYTOALEXIN DEFICIENT 4, AtPAD4) |
| 30 | TRINITY_DN2157_c0_g1 | 68.38 | 76.54 | 36.2 | 215.2 | 233.04 | 270.04 | −2.06397 | 3.08E-06 | Predicted Protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moy, A.; Nkongolo, K. Decrypting Molecular Mechanisms Involved in Counteracting Copper and Nickel Toxicity in Jack Pine (Pinus banksiana) Based on Transcriptomic Analysis. Plants 2024, 13, 1042. https://doi.org/10.3390/plants13071042
Moy A, Nkongolo K. Decrypting Molecular Mechanisms Involved in Counteracting Copper and Nickel Toxicity in Jack Pine (Pinus banksiana) Based on Transcriptomic Analysis. Plants. 2024; 13(7):1042. https://doi.org/10.3390/plants13071042
Chicago/Turabian StyleMoy, Alistar, and Kabwe Nkongolo. 2024. "Decrypting Molecular Mechanisms Involved in Counteracting Copper and Nickel Toxicity in Jack Pine (Pinus banksiana) Based on Transcriptomic Analysis" Plants 13, no. 7: 1042. https://doi.org/10.3390/plants13071042