
Re-Evaluation of Vascular Histogenesis in the Root Tips of Selected Species in the Poaceae Using New Methods: Analysis of the Plerome, Vascular Initials, Pericycle and Late-Maturing Metaxylem Vessels


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Preparation for Light Microscopy (LM)
2.3. Image Processing
2.4. Tracing Cell Files Derived from Vascular Initials
2.5. Counting the Numbers of Pericycle Cells, Vascular Cells, and Vascular Initials
3. Results
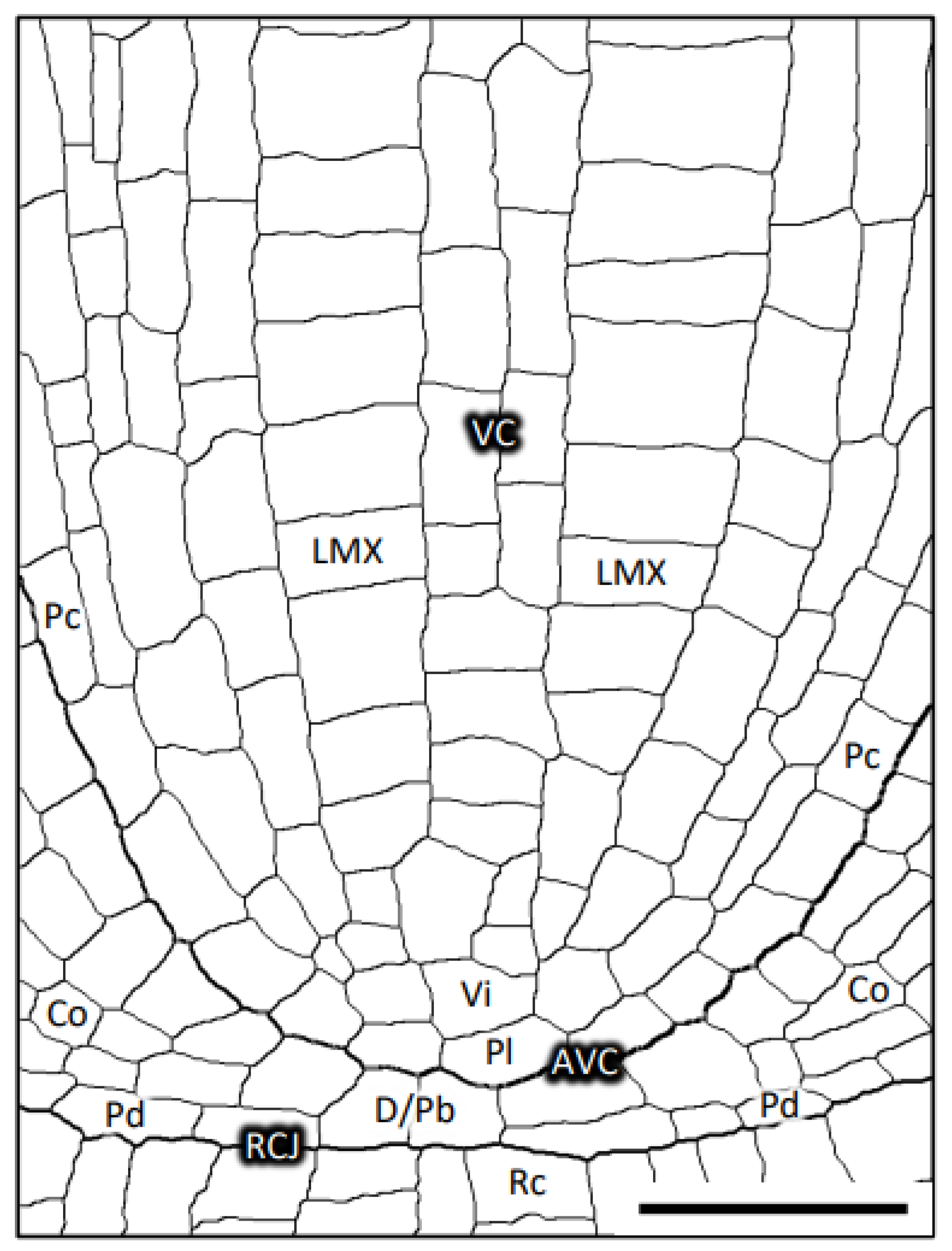
3.1. Plerome, Vascular Initials and LMXs of Teosinte
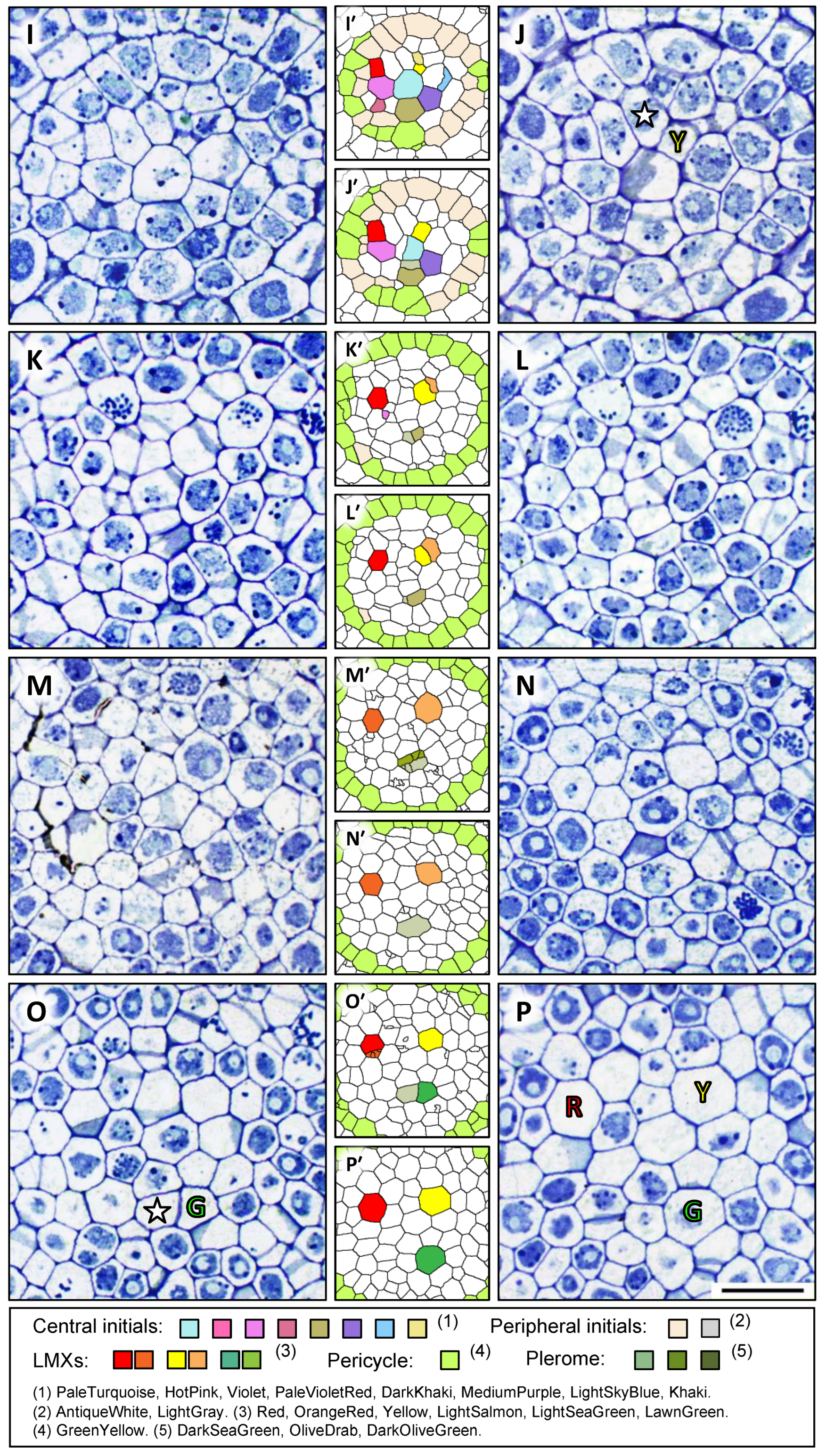
 ) and a collar cell (☆), shown in Figure 2H. The differentiation of the second LMX (
) and a collar cell (☆), shown in Figure 2H. The differentiation of the second LMX ( ) could be seen in the section 42 µm from the RCJ and the corresponding initial cell (Khaki in Figure 2I′) divided into an LMX () and a collar cell (☆ in Figure 2J). The differentiation of the third LMX (
) could be seen in the section 42 µm from the RCJ and the corresponding initial cell (Khaki in Figure 2I′) divided into an LMX () and a collar cell (☆ in Figure 2J). The differentiation of the third LMX ( ) followed a slightly more complicated process. The daughter cell produced by one of the central initials (DarkKhaki, Figure 2I′) underwent a subsequent cell division once at 58 µm from the RCJ in the axial plane (Figure 2M). These derivative cells differentiated into an LMX element () and a collar cell (☆) at the section 69 μm from the RCJ (Figure 2O). All the LMXs (, , and ) could be detected at 80 µm from the RCJ (Figure 2P,P′).
) followed a slightly more complicated process. The daughter cell produced by one of the central initials (DarkKhaki, Figure 2I′) underwent a subsequent cell division once at 58 µm from the RCJ in the axial plane (Figure 2M). These derivative cells differentiated into an LMX element () and a collar cell (☆) at the section 69 μm from the RCJ (Figure 2O). All the LMXs (, , and ) could be detected at 80 µm from the RCJ (Figure 2P,P′).3.2. Cells Detected as Vascular Initials in Teosinte Promeristems
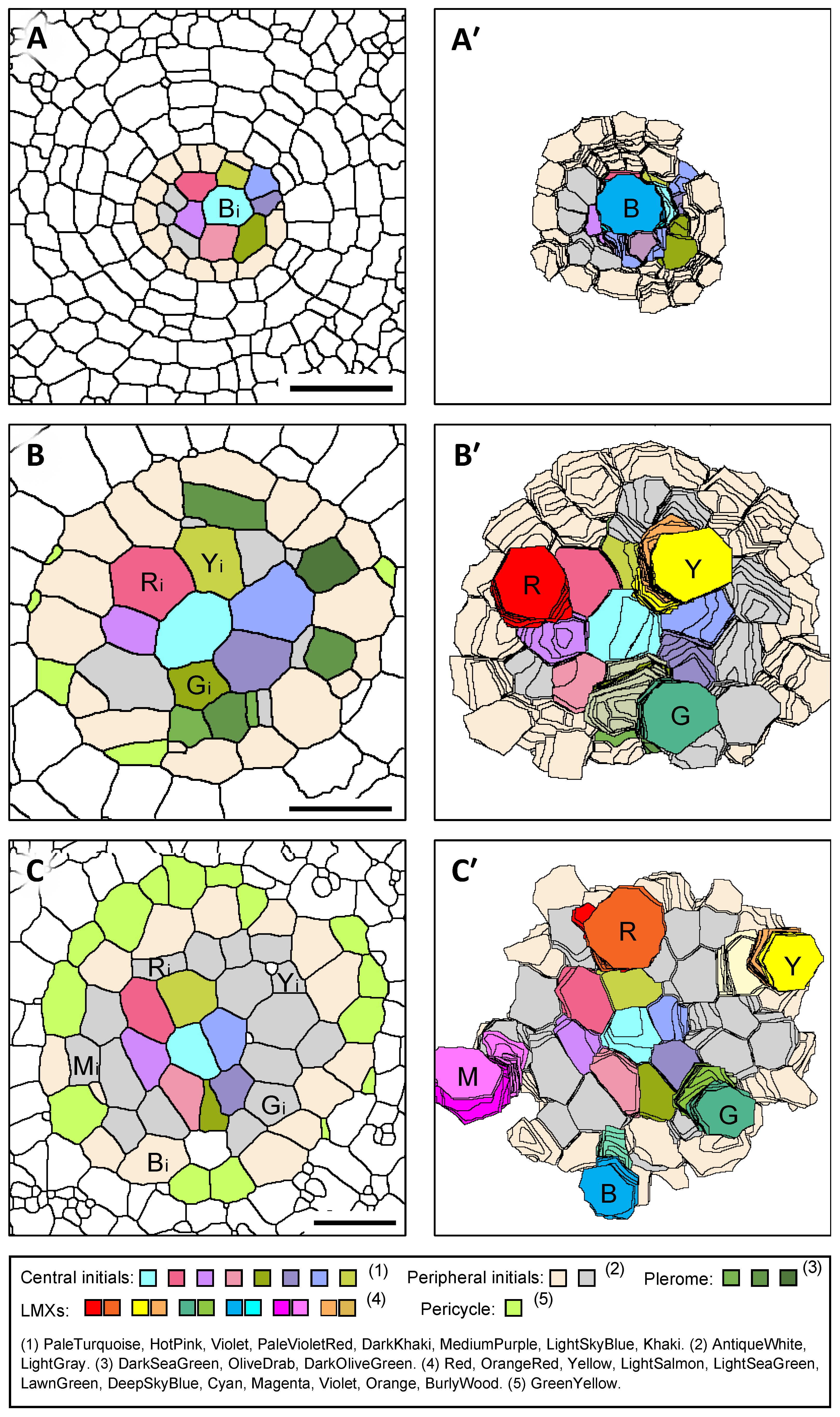
3.3. 3D Structure of the Plerome and Vascular Initials of Teosinte
3.4. 3D Structure of LMXs of Teosinte
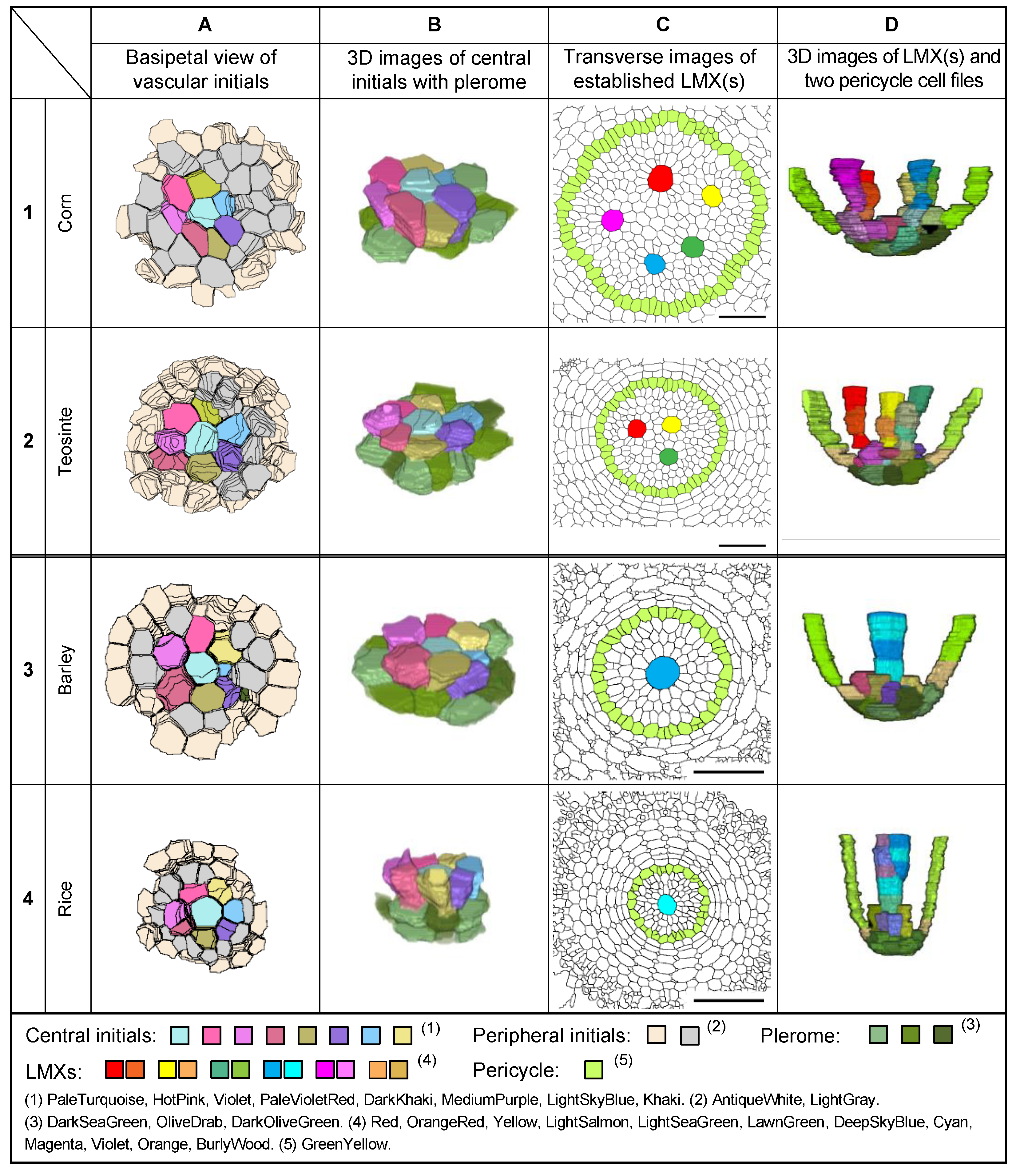
3.5. Plerome, Vascular Initials, and LMXs of Rice
3.6. Arrangement of Initials That Generate LMXs
3.7. Plerome, Vascular Initials, and LMXs of Sweet Corn and Barley
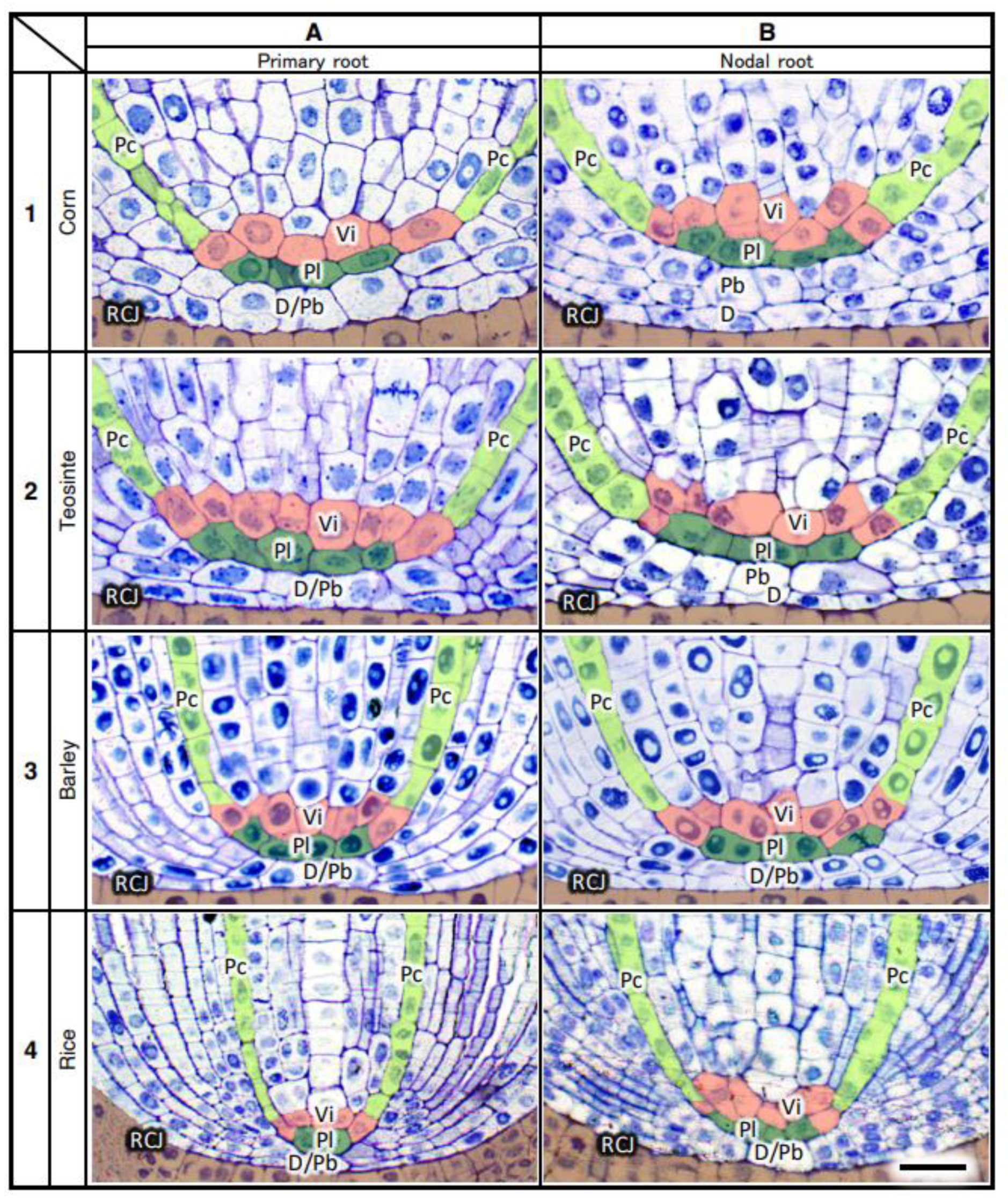
3.8. Structure of the Root Promeristems of Four Taxa in the Poaceae
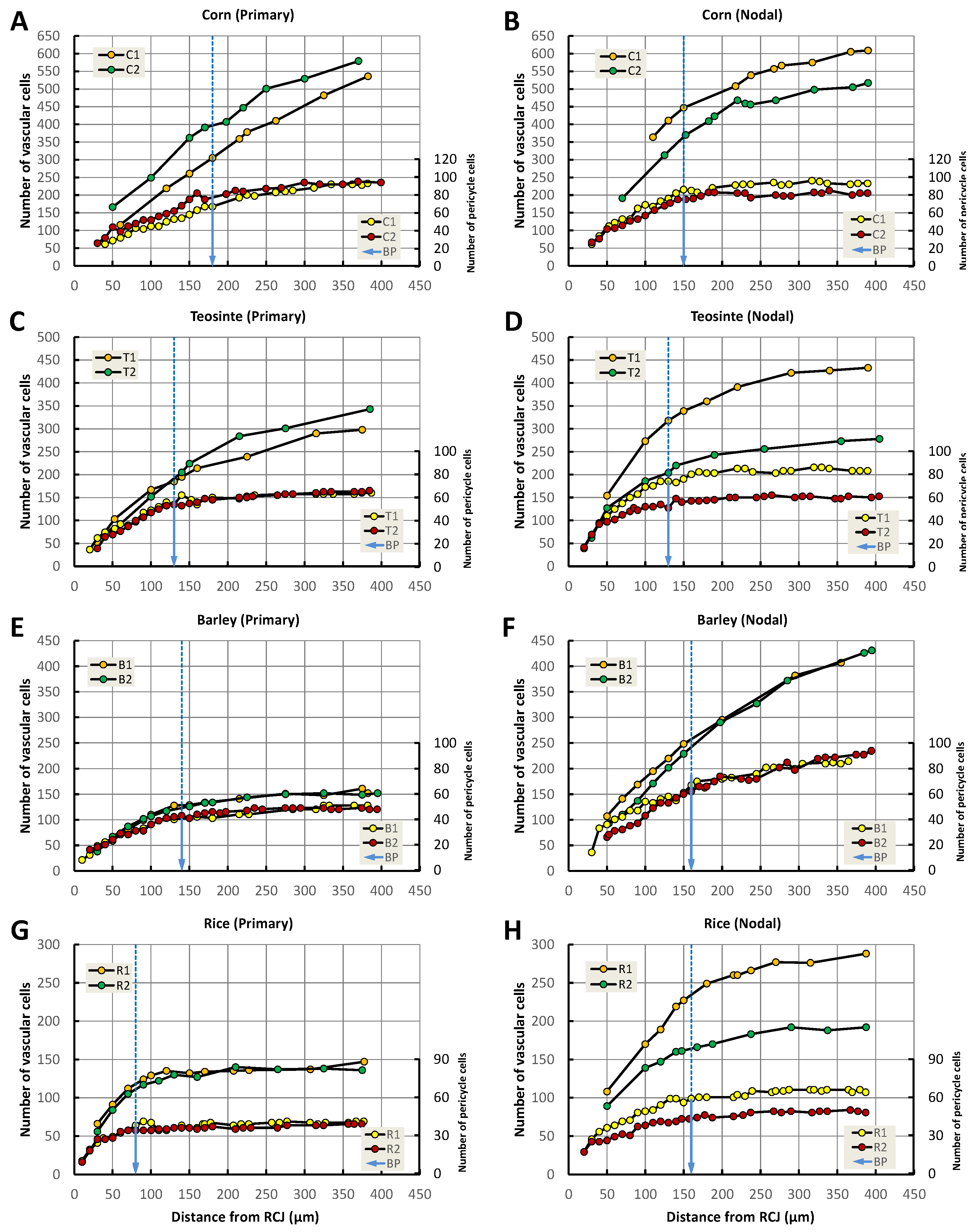
3.9. Increase in Pericycle Cell and Vascular Cell Numbers
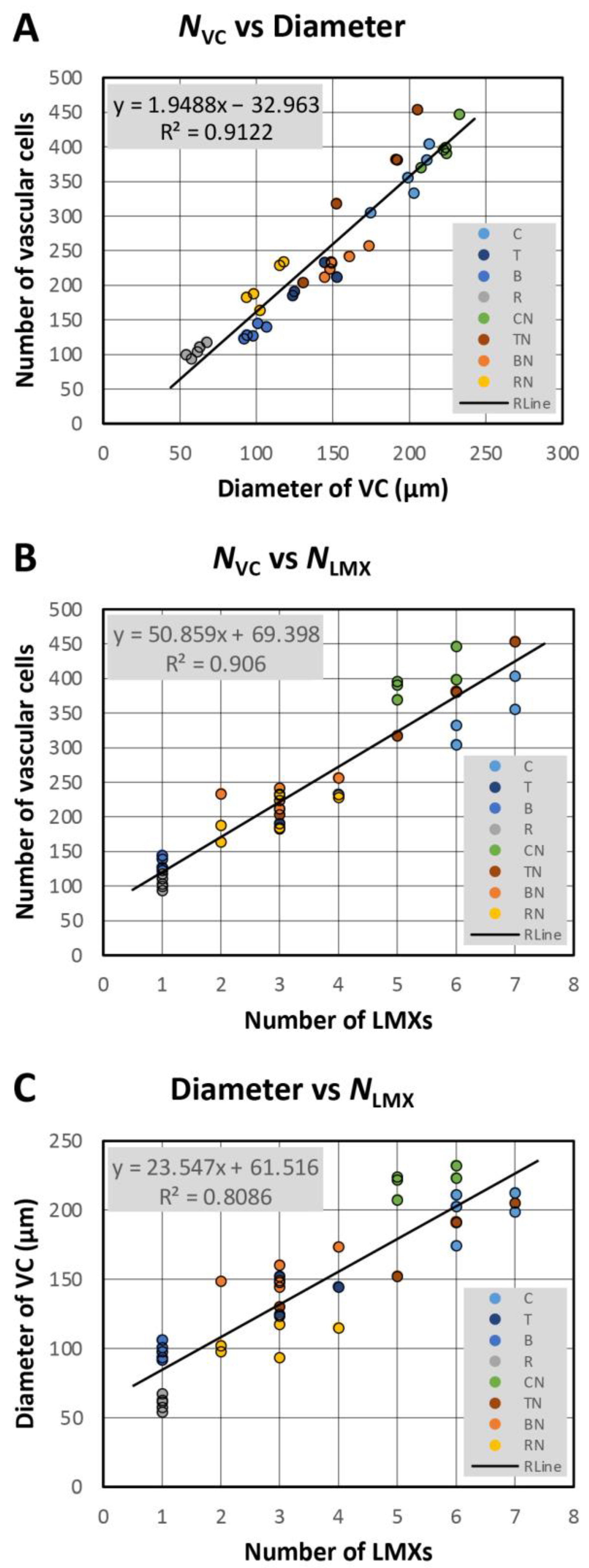
3.10. Number of Vascular Cells vs. Number of Vascular Initials
3.11. Diameter of the VC, Average Size of Vascular Cells, and the Number of LMXs
4. Discussion
4.1. Description of the Root VC Apical Structure of Four Taxa in the Poaceae
4.2. LMXs Generated from Vascular Initials
4.3. Boundary Point for the Increase in Pericycle Cell and Vascular Cell Numbers
4.4. Number of Vascular Initials vs. Vascular Cells, Cell Size and LMX
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Hanstein, J. Die Entwicklung des Keimes der Monokotylen und der Dikotylen. Bot. Abhandl. 1870, 1, 1–112. [Google Scholar]
- Schüepp, O. Meristeme. In Handbuch der Pflanzenanatomie; Borntraeger: Berlin, Germany, 1926. [Google Scholar]
- Clowes, F.A.L. Root apical meristems of Fagus sylvatica. New Phytol. 1950, 49, 248–268. [Google Scholar] [CrossRef]
- Heimsch, C.; Seago, J.L., Jr. Organization of the root apical meristem in angiosperms. Am. J. Bot. 2008, 95, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Clowes, F.A.L. Development of quiescent centres in root meristems. New Phytol. 1958, 57, 85–88. [Google Scholar] [CrossRef]
- von Guttenberg, H. Grundzüge der Histogenese Höherer Pflanzen. I. Die Angiospermen. Handbuch der Pflanzenanatomie; Band 8, Teil 3; Gebrüder Borntraeger: Berlin, Germany, 1960; pp. 72–103. [Google Scholar]
- Baum, S.E.; Dubrovsky, J.G.; Rost, T.L. Apical organization and maturation of the cortex and vascular cylinder in Arabidopsis thaliana (Brassicaceae) root. Am. J. Bot. 2002, 89, 908–920. [Google Scholar] [CrossRef]
- Saito, S.; Niki, T.; Gladish, D.K. Comparison of promeristem structure and ontogeny of procambium in primary roots of Zea mays ssp. mexicana and Z. mays ‘Honey Bantam’ with emphasis on metaxylem vessel histogenesis. Plants 2019, 8, 162. [Google Scholar] [CrossRef]
- Clowes, F.A.L. The promeristem and the minimal constructional centre in grass root apices. New Phytol. 1954, 53, 108–116. [Google Scholar] [CrossRef]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organization of the Arabidopsis root. Development 1993, 119, 71–84. [Google Scholar] [CrossRef]
- Heimsch, C. Development of vascular tissues in barley roots. Am. J. Bot. 1951, 38, 523–537. [Google Scholar] [CrossRef]
- Kawata, S.; Nishimaki, K.; Yamazaki, K. The apical structure of crown roots in rice plants. Jpn. J. Crop Sci. 1977, 46, 393–402, With summary in English. [Google Scholar] [CrossRef]
- Kawata, S.; Morita, S.; Yamazaki, K. On the differentiation of vessels and sieve tubes at the root tips of rice plants. Jpn. J. Crop Sci. 1978, 47, 101–110, With summary in English. [Google Scholar] [CrossRef]
- Kawata, S.; Morita, S.; Yamazaki, K. On the number of vessels and sieve tubes in crown roots of rice plants. Jpn. J. Crop Sci. 1979, 48, 502–509, With summary in English. [Google Scholar] [CrossRef]
- Liu, Z.; Giehl, R.; Hartmann, A.; Hajirezaei, M.; Carpentier, S.; von Wiren, N. Seminal and nodal roots of barley differ in anatomy, proteome, and nitrate uptake capacity. Plant Cell Physiol. 2020, 61, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Niki, T.; Saito, S.; Gladish, D.K. A novel thin section preparation and staining protocol to increase contrast and resolution of cell details for light microscopy. Biotech. Histochem. 2019, 94, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Saito, S.; Niki, T.; Gladish, D.K. Three-dimensional digital image construction of metaxylem vessels in root tips of Zea mays subsp. mexicana from thin transverse sections. Appl. Plant Sci. 2020, 8, e11347. [Google Scholar] [CrossRef]
- Miki, Y.; Saito, S.; Niki, T.; Gladish, D.K. Improved image processing for 3D virtual object construction from serial sections reveals tissue patterns in root tips of Zea mays. Appl. Plant Sci. 2023, 11, e11531. [Google Scholar] [CrossRef] [PubMed]
- Gladish, D.K.; Niki, T. Factors inducing cavity formation in the vascular cylinders of pea roots (Pisum sativum L., cv. ‘Alaska’). Environ. Exp. Bot. 2000, 43, 1–9. [Google Scholar] [CrossRef]
- Niki, T.; Gladish, D.K.; Lu, P.; Rost, T.L. Cellular changes precede cavity formation in the vascular cylinder of pea roots (Pisum sativum L. cv. ‘Alaska’). Int. J. Plant Sci. 1995, 156, 290–302. [Google Scholar] [CrossRef]
- Sugiura, M.; Takeda, Y. Nucleic acids. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; Chapter 6; p. 260. [Google Scholar]
- Morita, S.; Nemoto, K. Morphology and anatomy of rice roots with special reference to coordination in organo- and histogenesis. In Structure and Function of Roots: Proceedings of the Fourth International Symposium on Structure and Function of Roots; Baluska, F., Ciamporova, M., Gasparikova, O., Barlow, P., Eds.; Kluwer: Dordrecht, The Netherlands, 1995; pp. 75–86. [Google Scholar]
- Jiang, K.; Feldman, L.J. Regulation of root apical meristem development. Annu. Rev. Cell Dev. Biol. 2005, 21, 485–509. [Google Scholar] [CrossRef]
- Avery, G.S. Comparative anatomy and morphology of embryo and seedling of maize, oats and wheat. Bot. Gaz. 1930, 81, 1–39. [Google Scholar] [CrossRef]
- Saito, S.; Takahashi, M.; Niki, T. The differentiation of xylem on embryogenesis of Zea mays ‘Honey Bantam’, JSRR 45th Annual Meeting 2016, Okayama. Root Res. 2016, 25, 94. (In Japanese) [Google Scholar]
- Barlow, P. Regeneration of the cap of primary roots of Zea mays. New Phytol. 1974, 73, 937–954. [Google Scholar] [CrossRef]
- Torrey, J.G. Auxin control of vascular pattern formation in regenerating pea root meristems grown in vitro. Am. J. Bot. 1957, 44, 859–870. [Google Scholar] [CrossRef]
- Torrey, J.G.; Fosket, D.E.; Hepler, P.K. Xylem formation: A paradigm of cytodifferentiation in higher plants. Am. Sci. 1971, 59, 338–352. [Google Scholar]
- Alarcon, M.V.; Lloret, P.G.; Salguero, J. The development of the maize root system: Role of auxin and ethylene. In Root Engineering: Basic and Applied Concepts; Morte, A., Varma, A., Eds.; Springer: Heidelberg, Germany, 2014; pp. 75–103. [Google Scholar]
- Mironova, V.V.; Omelyanchuk, N.A.; Novoselova, E.S.; Doroshkov, A.V.; Kazantsev, F.V.; Kochetov, A.V.; Kolchanov, N.A.; Mjolsness, E.; Likhoshvai, V.A. Combined in silico/in vivo analysis of mechanisms providing for root apical meristem self-organization and maintenance. Ann. Bot. 2012, 110, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Baluska, F.; Volkman, D.; Barlow, P. A polarity crossroad in the transition growth zone of maize root apices: Cytoskeletal and developmental implications. J. Plant Growth Regul. 2001, 20, 170–181. [Google Scholar] [CrossRef]
- Schiefelbein, J.W.; Masucci, J.D.; Wang, H. Building a root: The control of patterning and morphogenesis during root development. Plant Cell 1997, 9, 1089–1098. [Google Scholar] [CrossRef]
- Scheres, B.; Benfey, P.; Dolan, L. Root development. Arab. Book/Am. Soc. Plant Biol. 2002, 1, e0101. [Google Scholar] [CrossRef]
 ) and a collar cell (☆) that differentiated from the initial in (G′) (HotPink) can be recognized in (H′). Scale bar = 20 µm. Selected transverse sections of a root apex of teosinte from 41 to 80 μm from the RCJ. The sections shown in (I–P) were 41, 42, 49, 50, 58, 63, 69 and 80 µm from the RCJ, respectively. Pericycle cells (GreenYellow) produced by peripheral initials (AntiqueWhite) can be seen in (I′,J′). An LMX () and a collar cell (☆) can be recognized in (J). Two LMXs ( and ) can be seen in (K–L). The daughter cell derived from a peripheral initial (LightGray) in (L′–O′) divided to form the third xylem initial (a secondary LMX initial, ) and a collar cell (☆) as shown in (O). All three LMXs (, and ) are shown in (P). Scale bar = 20 μm. Three LMXs that we labeled red, yellow, and green in (P′) were derived from the corresponding original points with the same colors (H′,J′,O′), and the progression of the development of the LMXs were observed.
) and a collar cell (☆) that differentiated from the initial in (G′) (HotPink) can be recognized in (H′). Scale bar = 20 µm. Selected transverse sections of a root apex of teosinte from 41 to 80 μm from the RCJ. The sections shown in (I–P) were 41, 42, 49, 50, 58, 63, 69 and 80 µm from the RCJ, respectively. Pericycle cells (GreenYellow) produced by peripheral initials (AntiqueWhite) can be seen in (I′,J′). An LMX () and a collar cell (☆) can be recognized in (J). Two LMXs ( and ) can be seen in (K–L). The daughter cell derived from a peripheral initial (LightGray) in (L′–O′) divided to form the third xylem initial (a secondary LMX initial, ) and a collar cell (☆) as shown in (O). All three LMXs (, and ) are shown in (P). Scale bar = 20 μm. Three LMXs that we labeled red, yellow, and green in (P′) were derived from the corresponding original points with the same colors (H′,J′,O′), and the progression of the development of the LMXs were observed.
) and a collar cell (☆) that differentiated from the initial in (G′) (HotPink) can be recognized in (H′). Scale bar = 20 µm. Selected transverse sections of a root apex of teosinte from 41 to 80 μm from the RCJ. The sections shown in (I–P) were 41, 42, 49, 50, 58, 63, 69 and 80 µm from the RCJ, respectively. Pericycle cells (GreenYellow) produced by peripheral initials (AntiqueWhite) can be seen in (I′,J′). An LMX () and a collar cell (☆) can be recognized in (J). Two LMXs ( and ) can be seen in (K–L). The daughter cell derived from a peripheral initial (LightGray) in (L′–O′) divided to form the third xylem initial (a secondary LMX initial, ) and a collar cell (☆) as shown in (O). All three LMXs (, and ) are shown in (P). Scale bar = 20 μm. Three LMXs that we labeled red, yellow, and green in (P′) were derived from the corresponding original points with the same colors (H′,J′,O′), and the progression of the development of the LMXs were observed.
) and a collar cell (☆) that differentiated from the initial in (G′) (HotPink) can be recognized in (H′). Scale bar = 20 µm. Selected transverse sections of a root apex of teosinte from 41 to 80 μm from the RCJ. The sections shown in (I–P) were 41, 42, 49, 50, 58, 63, 69 and 80 µm from the RCJ, respectively. Pericycle cells (GreenYellow) produced by peripheral initials (AntiqueWhite) can be seen in (I′,J′). An LMX () and a collar cell (☆) can be recognized in (J). Two LMXs ( and ) can be seen in (K–L). The daughter cell derived from a peripheral initial (LightGray) in (L′–O′) divided to form the third xylem initial (a secondary LMX initial, ) and a collar cell (☆) as shown in (O). All three LMXs (, and ) are shown in (P). Scale bar = 20 μm. Three LMXs that we labeled red, yellow, and green in (P′) were derived from the corresponding original points with the same colors (H′,J′,O′), and the progression of the development of the LMXs were observed.
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miki, Y.; Saito, S.; Niki, T.; Gladish, D.K. Re-Evaluation of Vascular Histogenesis in the Root Tips of Selected Species in the Poaceae Using New Methods: Analysis of the Plerome, Vascular Initials, Pericycle and Late-Maturing Metaxylem Vessels. Plants 2024, 13, 910. https://doi.org/10.3390/plants13060910
Miki Y, Saito S, Niki T, Gladish DK. Re-Evaluation of Vascular Histogenesis in the Root Tips of Selected Species in the Poaceae Using New Methods: Analysis of the Plerome, Vascular Initials, Pericycle and Late-Maturing Metaxylem Vessels. Plants. 2024; 13(6):910. https://doi.org/10.3390/plants13060910
Chicago/Turabian StyleMiki, Yasushi, Susumu Saito, Teruo Niki, and Daniel K. Gladish. 2024. "Re-Evaluation of Vascular Histogenesis in the Root Tips of Selected Species in the Poaceae Using New Methods: Analysis of the Plerome, Vascular Initials, Pericycle and Late-Maturing Metaxylem Vessels" Plants 13, no. 6: 910. https://doi.org/10.3390/plants13060910