

Salt Water Exposure Exacerbates the Negative Response of Phragmites australis Haplotypes to Sea-Level Rise

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Site Differences and Correlations across Abiotic Factors
2.2. Plant Survival
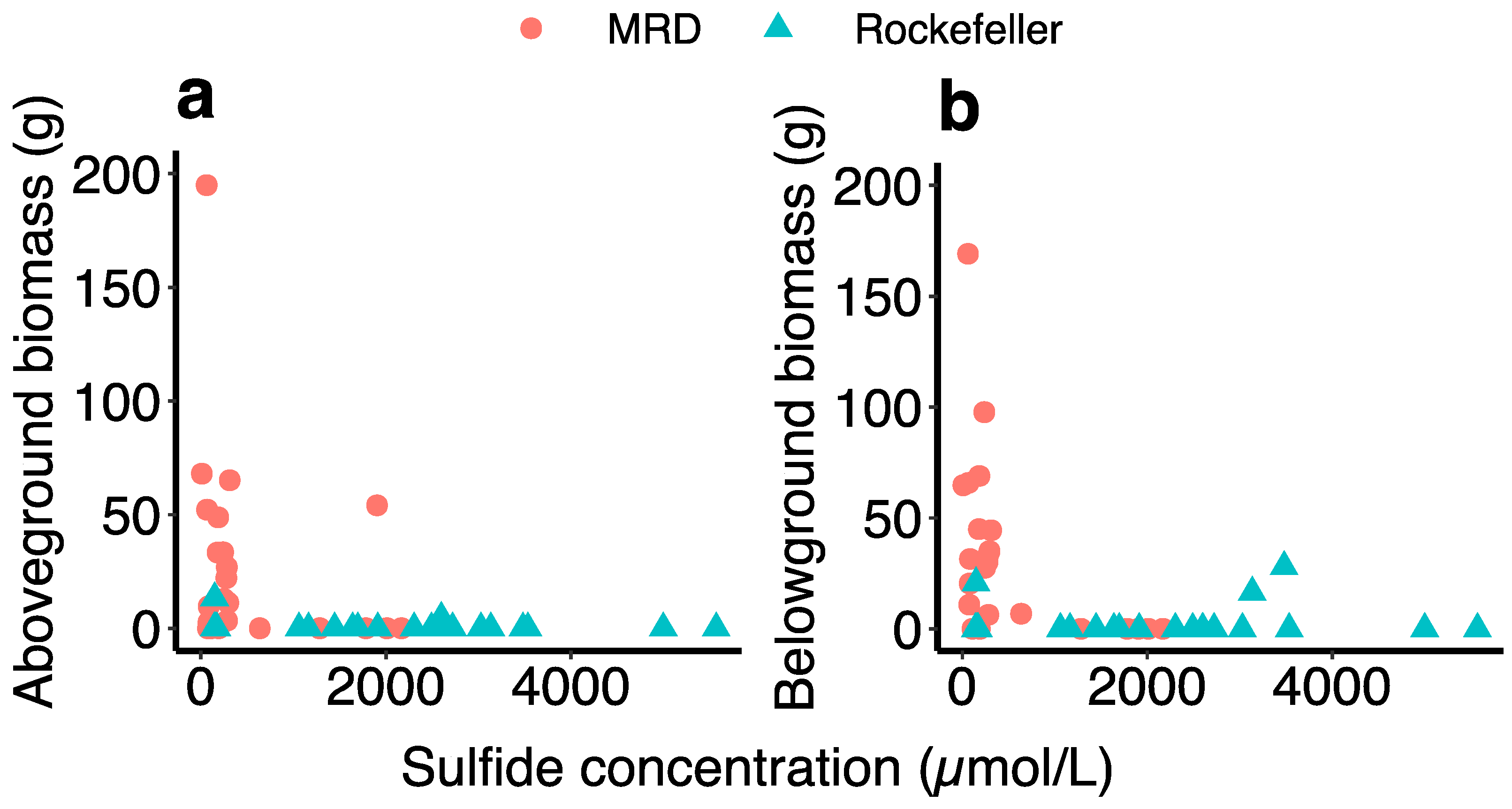
2.3. Aboveground Biomass
2.4. Belowground Biomass
3. Discussion
3.1. Phragmites australis Lineages
3.2. Response to Abiotic Stress
3.3. Management Implications
4. Materials and Methods
4.1. Study Locations
4.2. Plant Collection
4.3. Experimental Design and Marsh Organs
4.4. Measurements
4.4.1. Flooding Dynamics
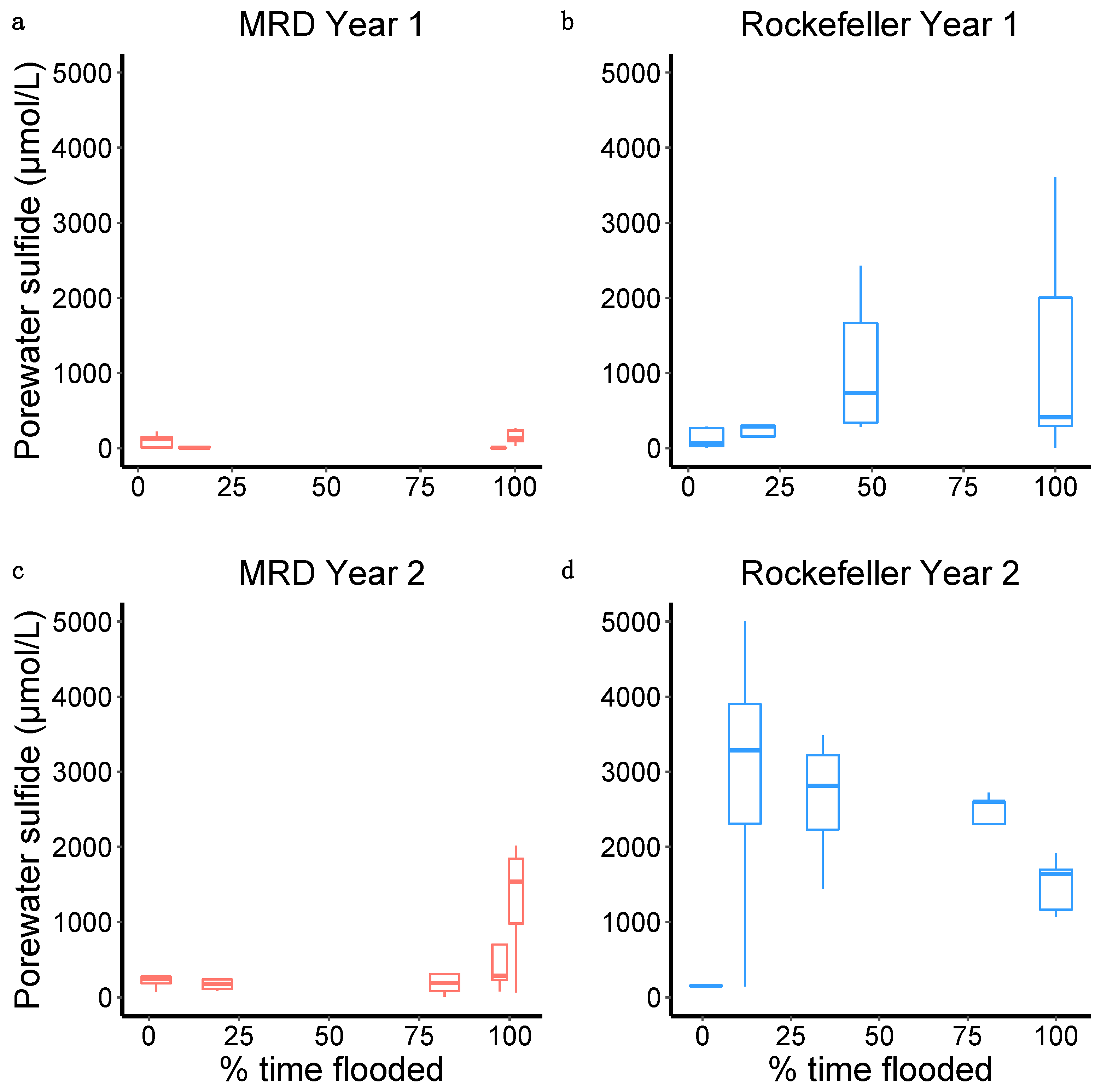
4.4.2. Porewater Chemistry and Redox Potential
4.4.3. Survivorship and Productivity
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goberville, E.; Hautekèete, N.C.; Kirby, R.R.; Piquot, Y.; Luczak, C.; Beaugrand, G. Climate Change and the Ash Dieback Crisis. Sci. Rep. 2016, 6, 35303. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Ruiz-Benito, P.; Greenwood, S.; Allen, C.D.; Kitzberger, T.; Fensham, R.; Martínez-Vilalta, J.; Lloret, F. Structural Overshoot of Tree Growth with Climate Variability and the Global Spectrum of Drought-Induced Forest Dieback. Glob. Change Biol. 2017, 23, 3742–3757. [Google Scholar] [CrossRef]
- McKee, K.L.; Mendelssohn, I.A.; Materne, M.D. Acute Salt Marsh Dieback in the Mississippi River Deltaic Plain: A Drought-Induced Phenomenon? Glob. Ecol. Biogeogr. 2004, 13, 65–73. [Google Scholar] [CrossRef]
- Strzepek, K.; Yohe, G.; Neumann, J.; Boehlert, B. Characterizing Changes in Drought Risk for the United States from Climate Change. Environ. Res. Lett. 2010, 5, 3742–3757. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Hernandez, M.E. Landscape and Climate Change Threats to Wetlands of North and Central America. Aquat. Sci. 2013, 75, 133–149. [Google Scholar] [CrossRef]
- Cahoon, D.R.; McKee, K.L.; Morris, J.T. How Plants Influence Resilience of Salt Marsh and Mangrove Wetlands to Sea-Level Rise. Estuaries Coasts 2021, 44, 883–898. [Google Scholar] [CrossRef]
- Nyman, J.A.; Delaune, R.D.; Roberts, H.H.; Patrick, W.H. Relationship between Vegetation and Soil Formation in a Rapidly Submerging Coastal Marsh. Mar. Ecol. Prog. Ser. 1993, 96, 269–279. [Google Scholar] [CrossRef]
- Nyman, J.A.; Walters, R.J.; Delaune, R.D.; Patrick, W.H. Marsh Vertical Accretion via Vegetative Growth. Estuar. Coast. Shelf Sci. 2006, 69, 370–380. [Google Scholar] [CrossRef]
- Clevering, O.A.; Lissner, J. Taxonomy, Chromosome Numbers, Clonal Diversity and Population Dynamics of Phragmites australis. Aquat. Bot. 1999, 64, 185–208. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Saltonstall, K.; Windham, L.; Kiviat, E.; Findlay, S. A Comparison of Phragmites australis in Freshwater and Brackish Marsh Environments in North America. Wetl. Ecol. Manag. 2000, 8, 89–103. [Google Scholar] [CrossRef]
- Saltonstall, K. The Naming of Phragmites Haplotypes. Biol. Invasions 2016, 18, 2433–2441. [Google Scholar] [CrossRef]
- League, M.T.; Colbert, E.P.; Seliskar, D.M.; Gallagher, J.L. Rhizome Growth Dynamics of Native and Exotic Haplotypes of Phragmites australis (Common Reed). Estuaries Coasts 2006, 29, 269–276. [Google Scholar] [CrossRef]
- Benoit, L.K.; Askins, R.A. Impact of the Spread of Phragmites on the Distribution of Birds in Connecticut Tidal Marshes. Wetlands 1999, 19, 194–208. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Cronin, J.T. Evidence for Multiple Introductions of Phragmites australis to North America: Detection of a New Non-Native Haplotype. Biol. Invasions 2013, 15, 2605–2608. [Google Scholar] [CrossRef]
- Nguyen, L.X.; Lambertini, C.; Sorrell, B.K.; Eller, F.; Achenbach, L.; Brix, H. Photosynthesis of Co-Existing Phragmites Haplotypes in Their Non-Native Range: Are Characteristics Determined by Adaptations Derived from Their Native Origin? AoB Plants 2013, 5, plt016. [Google Scholar] [CrossRef]
- Lambertini, C.; Mendelssohn, I.A.; Gustafsson, M.H.G.; Olesen, B. Tracing the Origin of Gulf Coast Phragmites (Poaceae): A Story of Long-Distance Dispersal and Hybridization. Am. J. Bot. 2012, 99, 538–551. [Google Scholar] [CrossRef]
- Achenbach, L.; Eller, F.; Nguyen, L.X.; Brix, H. Differences in Salinity Tolerance of Genetically Distinct Phragmites australis Clones. AoB Plants 2013, 5, plt019. [Google Scholar] [CrossRef]
- Achenbach, L.; Brix, H. Can Differences in Salinity Tolerance Explain the Distribution of Four Genetically Distinct Lineages of Phragmites australis in the Mississippi River Delta? Hydrobiologia 2014, 737, 5–23. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W.; Van Der Putten, W.H. Phragmites Die-Back: Bud and Root Death, Blockages within the Aeration and Vascular Systems and the Possible Role of Phytotoxins. New Phytol. 1996, 133, 399–414. [Google Scholar] [CrossRef]
- Simkin, S.M.; Bedford, B.L.; Weathers, K.C. Phytotoxic Sulfide More Important than Nutrients for Plants Within a Groundwater-Fed Wetland. Ecosystems 2013, 16, 1118–1129. [Google Scholar] [CrossRef]
- Kotula, L.; Colmer, T.D.; Nakazono, M. Effects of Organic Acids on the Formation of the Barrier to Radial Oxygen Loss in Roots of Hordeum Marinum. Funct. Plant Biol. 2014, 41, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Snedden, G.A.; Cretini, K.; Patton, B. Inundation and Salinity Impacts to Above- and Belowground Productivity in Spartina Patens and Spartina alterniflora in the Mississippi River Deltaic Plain: Implications for Using River Diversions as Restoration Tools. Ecol. Eng. 2015, 81, 133–139. [Google Scholar] [CrossRef]
- Morris, J. Estimating Net Primary Production of Salt Marsh Macrophytes. In Principles and Standards for Measuring Primary Production; Fahey, T., Knapp, A., Eds.; Oxford University Press Inc.: New York, NY, USA, 2007; pp. 106–119. [Google Scholar]
- Kirwan, M.L.; Guntenspergen, G.R. Response of Plant Productivity to Experimental Flooding in a Stable and a Submerging Marsh. Ecosystems 2015, 18, 903–913. [Google Scholar] [CrossRef]
- Morris, J.T.; Sundberg, K.; Hopkinson, C.S. Salt Marsh Primary Production and Its Responses to Relative Sea Level and Nutrients in Estuaries at Plum Island, Massachusetts, and North Inlet, South Carolina, USA. Oceanogr. Soc. 2013, 26, 78–84. [Google Scholar] [CrossRef]
- Smith, J.A.M. The Role of Phragmites australis in Mediating Inland Salt Marsh Migration in a Mid-Atlantic Estuary. PLoS ONE 2013, 8, e0065091. [Google Scholar] [CrossRef] [PubMed]
- Kiviat, E. Ecosystem Services of Phragmites in North America with Emphasis on Habitat Functions. AoB Plants 2013, 5, plt008. [Google Scholar] [CrossRef]
- Rooth, J.E.; Stevenson, J.C.; Cornwell, J.C. Increased Sediment Accretion Rates Following Invasion by Phragmites australis: The Role of Litter. Estuaries 2003, 26, 475–483. [Google Scholar] [CrossRef]
- Gigante, D.; Angiolini, C.; Landucci, F.; Maneli, F.; Nisi, B.; Vaselli, O.; Venanzoni, R.; Lastrucci, L. New Occurrence of Reed Bed Decline in Southern Europe: Do Permanent Flooding and Chemical Parameters Play a Role? Comptes Rendus-Biol. 2014, 337, 487–498. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W.; Wu, Z.; Afreen-Zobayed, F. A Role for Phytotoxins in the Phragmites Die-Back Syndrome? Folia Geobot. Phytotax. 1996, 31, 127–142. [Google Scholar] [CrossRef]
- Cronin, J.T.; Johnston, J.; Diaz, R. Multiple Potential Stressors and Dieback of Phragmites australis in the Mississippi River Delta, USA: Implications for Restoration. Wetlands 2020, 40, 2247–2261. [Google Scholar] [CrossRef]
- Pagter, M.; Bragato, C.; Brix, H. Tolerance and Physiological Responses of Phragmites australis to Water Deficit. Aquat. Bot. 2005, 81, 285–299. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Christensen, A.; McCall, A.; Jensen, D.J.; Snedden, G.A.; Morris, J.T.; Cavell, J.A. Biomass Allocation of Tidal Freshwater Marsh Species in Response to Natural and Manipulated Hydroperiod in Coastal Deltaic Floodplains. Estuar. Coast. Shelf Sci. 2022, 268, 107784. [Google Scholar] [CrossRef]
- Lane, S.L. Using Marsh Organs to Test Seed Recruitment in Tidal Freshwater Marshes. Appl. Plant Sci. 2022, 10, e11474. [Google Scholar] [CrossRef]
- Armstrong, J.; Afreen-Zobayed, F.; Armstrong, W. Phragmites Die-Back: Sulphide- and Acetic Acid-Induced Bud and Root Death, Lignifications, and Blockages within Aeration and Vascular Systems. New Phytol. 1996, 134, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Van Der Putten, W.H. Die-Back of Phragmites australis in European Wetlands: An Overview of the European Research Programme on Reed Die-Back and Progression (1993–1994). Aquat. Bot. 1997, 59, 263–275. [Google Scholar] [CrossRef]
- McKee, K.L.; Mendelssohn, I.A. Response of a Freshwater Marsh Plant Community to Increased Salinity and Increased Water Level. Aquat. Bot. 1989, 34, 301–316. [Google Scholar] [CrossRef]
- Janousek, C.N.; Buffington, K.J.; Thorne, K.M.; Guntenspergen, G.R.; Takekawa, J.Y.; Dugger, B.D. Potential Effects of Sea-Level Rise on Plant Productivity: Species-Specific Responses in Northeast Pacific Tidal Marshes. Mar. Ecol. Prog. Ser. 2016, 548, 111–125. [Google Scholar] [CrossRef]
- Buffington, K.J.; Goodman, A.C.; Freeman, C.M.; Thorne, K.M. Testing the Interactive Effects of Flooding and Salinity on Tidal Marsh Plant Productivity. Aquat. Bot. 2020, 164, 103231. [Google Scholar] [CrossRef]
- Amsberry, L.; Baker, M.A.; Ewanchuk, P.J.; Bertness, M.D. Clonal Integration and the Expansion of Phragmites australis. Ecol. Appl. 2000, 10, 1110–1118. [Google Scholar] [CrossRef]
- Lee, H.; Diaz, R.; Johnston, J.; Knight, I.A.; Nyman, J.A.; Cronin, J.T. Vegetation Restoration Following Dieback of Phragmites australis in the Mississippi River Delta, USA. Wetlands 2023, 43, 98. [Google Scholar] [CrossRef]
- Hauber, D.P.; Saltonstall, K. Genetic Variation in the Common Reed, Phragmites australis, in the Mississippi River Delta Marshes: Evidence for Multiple Introductions. Estuaries Coasts 2011, 34, 851–862. [Google Scholar] [CrossRef]
- Visser, J.M.; Sandy, E.R. The Effects of Flooding on Four Common Louisiana Marsh Plants. Gulf Mex. Sci. 2009, 27, 21–29. [Google Scholar] [CrossRef]
- Kovacs, M.; Turcsanyi, G.; Tuba, Z.; Wolcsanszky, S.; Vasarhelyi, T.; Dely-Draskovits, A.; Toth, S.; Koltay, A.; Kaszab, L.; Szoke, P.; et al. The Decay of Reed in Hungarian Lakes. Symp. Biol. Hungarica 1989, 38, 461–471. [Google Scholar]
- Hellings, S.E.; Gallagher, J.L. The Effects of Salinity and Flooding on Phragmites australis. J. Appl. Ecol. 1992, 29, 41–49. [Google Scholar] [CrossRef]
- Chambers, R.M.; Mozdzer, T.J.; Ambrose, J.C. Effects of Salinity and Sulfide on the Distribution of Phragmites australis and Spartina alterniflora in a Tidal Saltmarsh. Aquat. Bot. 1998, 62, 161–169. [Google Scholar] [CrossRef]
- Knight, I.A.; Cronin, J.T.; Gill, M.; Nyman, J.A.; Wilson, B.E.; Diaz, R. Investigating Plant Phenotype, Salinity, and Infestation by the Roseau Cane Scale as Factors in the Die-Back of Phragmites australis in the Mississippi River Delta, USA. Wetlands 2020, 40, 1327–1337. [Google Scholar] [CrossRef]
- Lynch, J.M. Production and Phytotoxicity of Acetic Acid in Anaerobic Soils Containing Plant Residues. Soil Biol. Biochem. 1978, 10, 131–135. [Google Scholar] [CrossRef]
- Čížková, H.; Brix, H.; Kopecký, J.; Lukavská, J. Organic Acids in the Sediments of Wetlands Dominated by Phragmites australis: Evidence of Phytotoxic Concentrations. Aquat. Bot. 1999, 64, 303–315. [Google Scholar] [CrossRef]
- Mendelssohn, I.A.; Sorrell, B.K.; Brix, H.; Schierup, H.H.; Lorenzen, B.; Maltby, E. Controls on Soil Cellulose Decomposition along a Salinity Gradient in a Phragmites australis Wetland in Denmark. Aquat. Bot. 1999, 64, 381–398. [Google Scholar] [CrossRef]
- Himanen, M.; Prochazka, P.; Hänninen, K.; Oikari, A. Phytotoxicity of Low-Weight Carboxylic Acids. Chemosphere 2012, 88, 426–431. [Google Scholar] [CrossRef]
- Čížková, H.; Strand, J.A.; Lukavská, J. Factors Associated with Reed Decline in a Eutrophic Fishpond, Rožmberk (South Bohemia, Czech Republic). Folia Geobot. 1996, 31, 73–84. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W. An Overview of the Effects of Phytotoxins on Phragmites australis in Relation to Die-Back. Aquat. Bot. 2001, 69, 251–268. [Google Scholar] [CrossRef]
- Tully, K.; Gedan, K.; Epanchin-Niell, R.; Strong, A.; Bernhardt, E.S.; Bendor, T.; Mitchell, M.; Kominoski, J.; Jordan, T.E.; Neubauer, S.C.; et al. The Invisible Flood: The Chemistry, Ecology, and Social Implications of Coastal Saltwater Intrusion. Bioscience 2019, 69, 368–378. [Google Scholar] [CrossRef]
- Jacobs, M.D. Phragmites australis Haplotypic Variation and Response to Abiotic Stressors; Louisiana State University: Baton Rouge, LA, USA, 2022. [Google Scholar]
- Lamers, M.; Govers, L.L.; Janssen, I.C.J.M.; Geurts, J.J.M.; Van Der Welle, M.E.W.; Van Katwijk, M.M.; Van Der Heide, T.; Roelofs, J.G.M.; Alfons, J.P. Sulfide as a Soil Phytotoxin—A Review. Front. Plant Sci. 2013, 4, 268. [Google Scholar] [CrossRef] [PubMed]
- Smolders, A.J.P.; Nijober, R.C.; Roelofs, J.G.M. Prevention of Sulphide Accumulation and Phosphate Mobilization by the Addition of Iron(II) Chloride to a Reduced Sediment: An Enclosure Experiment. Freshw. Biol. 1995, 34, 559–568. [Google Scholar] [CrossRef]
- Wu, S.; Kuschk, P.; Wiessner, A.; Müller, J.; Saad, R.A.B.; Dong, R. Sulphur Transformations in Constructed Wetlands for Wastewater Treatment: A Review. Ecol. Eng. 2013, 52, 278–289. [Google Scholar] [CrossRef]
- Chambers, R.M. Porewater Chemistry Associated with Phragmites and Spartina in a Connecticut Tidal Marsh. Wetlands 1997, 17, 360–367. [Google Scholar] [CrossRef]
- Swarzenski, C.M.; Doyle, T.W.; Fry, B.; Hargis, T.G. Biogeochemical Response of Organic-Rich Freshwater Marshes in the Louisiana Delta Plain to Chronic River Water Influx. Biogeochemistry 2008, 90, 49–63. [Google Scholar] [CrossRef]
- Hazelton, E.L.G.; Mozdzer, T.J.; Burdick, D.M.; Kettenring, K.M.; Whigham, D.F. Phragmites australis Management in the United States: 40 Years of Methods and Outcomes. AoB Plants 2014, 6, plu001. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.H.; Zhao, B.; Ma, Z.J.; Luo, Y.Q.; Chen, J.K.; Li, B. Effects of Growing Conditions on the Growth of and Interactions between Salt Marsh Plants: Implications for Invasibility of Habitats. Biol. Invasions 2006, 8, 1547–1560. [Google Scholar] [CrossRef]
- Annett, R.; Habibi, H.R.; Hontela, A. Impact of Glyphosate and Glyphosate-Based Herbicides on the Freshwater Environment. J. Appl. Toxicol. 2014, 34, 458–479. [Google Scholar] [CrossRef] [PubMed]
- Rohal, C.B.; Hazelton, E.L.G.; McFarland, E.K.; Downard, R.; McCormick, M.K.; Whigham, D.F.; Kettenring, K.M. Landscape and Site Factors Drive Invasive Phragmites Management and Native Plant Recovery across Chesapeake Bay Wetlands. Ecosphere 2023, 14, e4392. [Google Scholar] [CrossRef]
- Reddy, R.K.; D’Angelo, E.M.; Harris, W.G. Biogeochemistry of Wetlands. In Handbook of Soil Science; Sumner, M.E., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 89–119. [Google Scholar]
- Bhushan, A.; Yemane, D.; Trudell, D.; Overton, E.B.; Goettert, J. Fabrication of Micro-Gas Chromatograph Columns for Fast Chromatography. Microsyst. Technol. 2007, 13, 361–368. [Google Scholar] [CrossRef]
- Cassella, R.J.; de Oliveira, L.G.; Santelli, R.E. On Line Dissolution of ZnS For Sulfide Determination in Stabilized Water Samples with Zinc Acetate, Using Spectrophotometry by Methylene Blue Method. Spectrosc. Lett. 1999, 32, 469–484. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric Determination of Hydrogen Sulfide in Natural Waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Mylon, S.E.; Benoit, G. Subnanomolar Detection of Acid-Labile Sulfides by the Classical Methylene Blue Method Coupled to HPLC. Environ. Sci. Technol. 2001, 35, 4544–4548. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 December 2020).








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynn, A.; Elsey-Quirk, T. Salt Water Exposure Exacerbates the Negative Response of Phragmites australis Haplotypes to Sea-Level Rise. Plants 2024, 13, 906. https://doi.org/10.3390/plants13060906
Lynn A, Elsey-Quirk T. Salt Water Exposure Exacerbates the Negative Response of Phragmites australis Haplotypes to Sea-Level Rise. Plants. 2024; 13(6):906. https://doi.org/10.3390/plants13060906
Chicago/Turabian StyleLynn, Austin, and Tracy Elsey-Quirk. 2024. "Salt Water Exposure Exacerbates the Negative Response of Phragmites australis Haplotypes to Sea-Level Rise" Plants 13, no. 6: 906. https://doi.org/10.3390/plants13060906