

Morphological, Anatomical, and Physiological Characteristics of Heteroblastic Acacia melanoxylon Grown under Weak Light
Abstract
:1. Introduction
2. Results
2.1. Heteroblastic Phenotype and Growth of A. melanoxylon Leaves Induced by Weak Light Treatment
2.2. Differences in Anatomical Structure between Heteroblastic Leaves
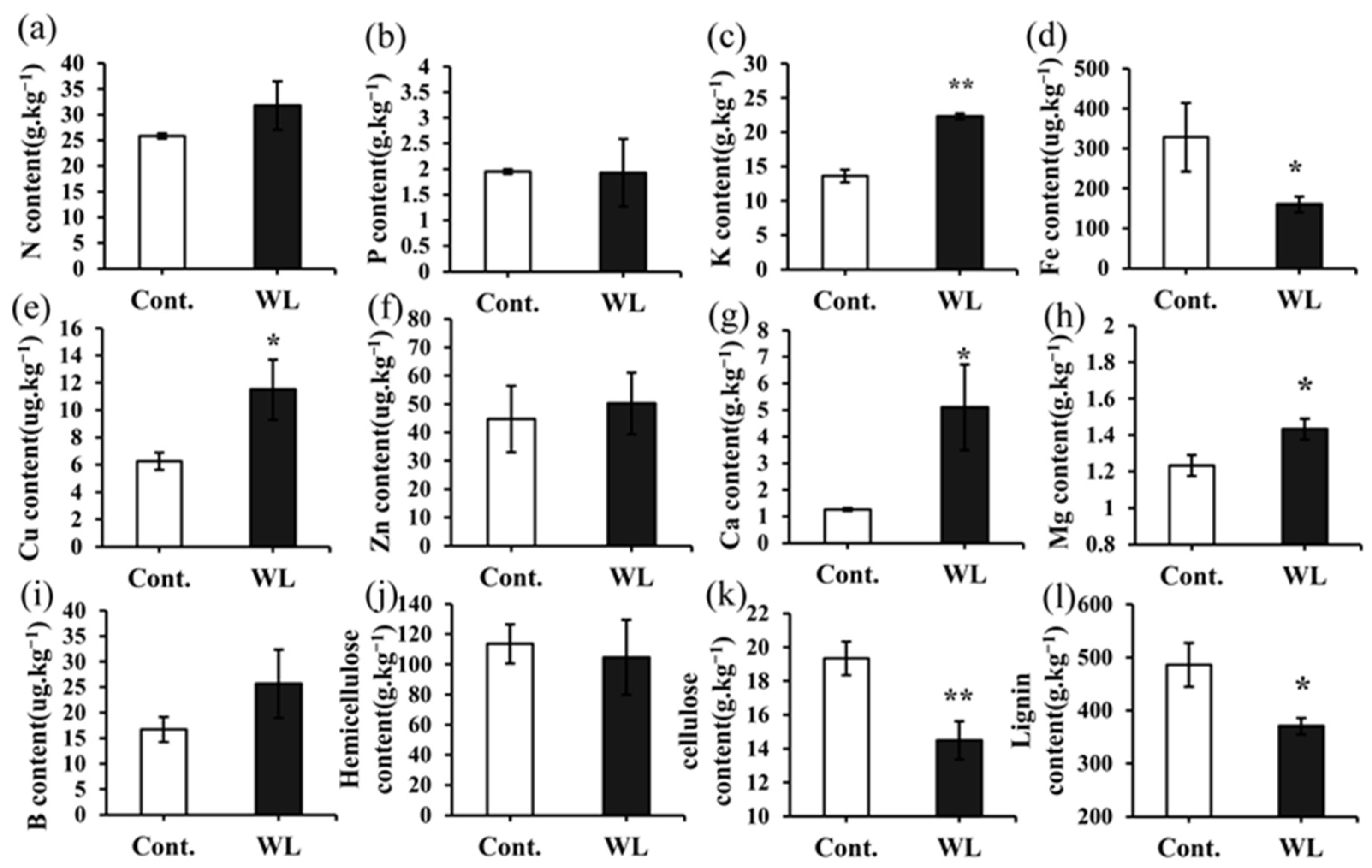
2.3. Differences in the Levels of Mineral Elements and Cell Wall Components between Heteroblastic Leaves
2.4. Differences in the Levels of Antioxidant Enzyme Activities and Osmotic Regulatory Substances between Heteroblastic Leaves
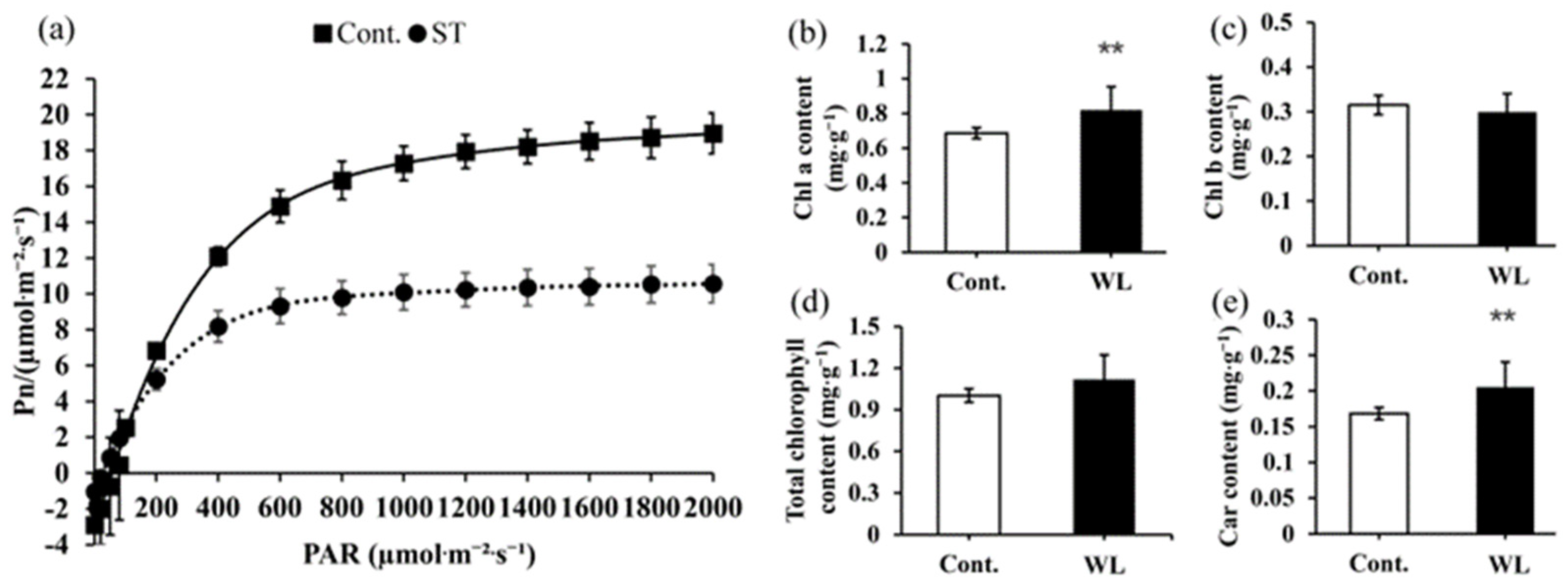
2.5. Differences in Light Utilization Capacity between Heteroblastic Leaves
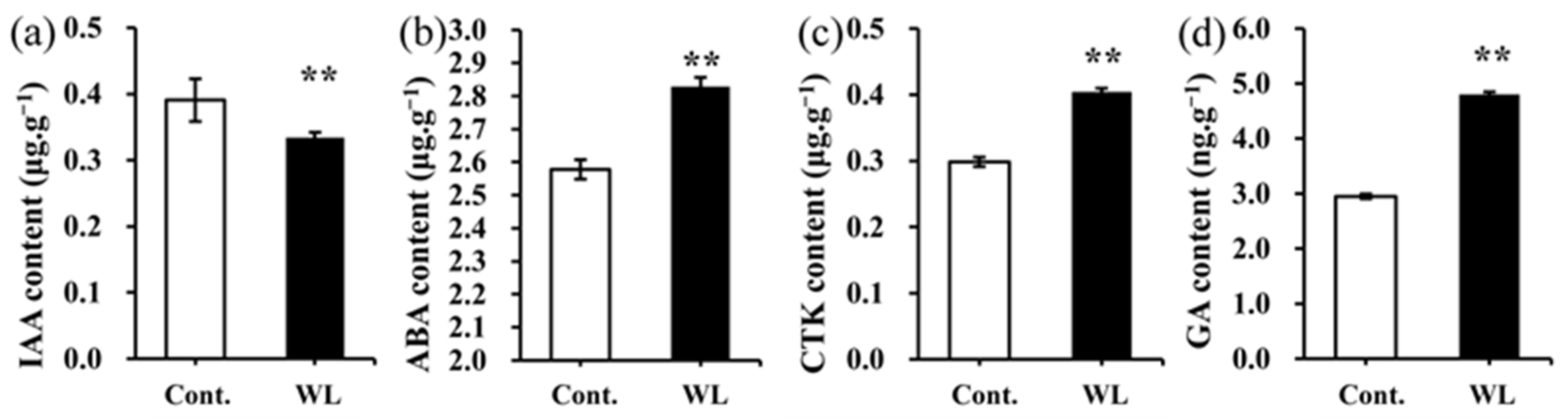
2.6. Differences in the Levels of Endogenous Hormone between Heteroblastic Leaves
2.7. Correlation Analysis of Growth and Development-Related Indicators in Heteroblastic Leaves
3. Discussion
3.1. Heteroblasty Is Highly Sensitive to the Induction of Weak Light in A. melanoxylon Leaves
3.2. Anatomical Structural Adaptive Characteristics of A. melanoxylon Heteroblastic Leaves
3.3. Variances in Anatomical Structure and Photosynthetic Pigment Levels Impact the Light Utilization Capacity of A. melanoxylon Heteroblastic Leaves
3.4. The Changes in Endogenous Hormones, the Oxidation–Reduction System, and Elemental Content Were Closely Linked throughout the Development of Weak Light-Induced Heteroblastic Leaves in A. melanoxylon
4. Materials and Methods
4.1. Plant Materials and Weak Light Treatments
4.2. Plant Growth Measurement
4.3. Leaf Anatomy and Stomatal Traits
4.4. Determination of Mineral Element Content
4.5. Determination of Cell Wall Component Contents
4.6. Determination of Antioxidant Enzyme Activity and Pro Content
4.7. Measurement of the Light Response Curve
4.8. Determination of Photosynthetic Pigment Contents
4.9. Determination of Endogenous Hormone Levels
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fritz, M.A.; Rosa, S.; Sicard, A. Mechanisms underlying the environmentally induced plasticity of leaf morphology. Front. Genet. 2018, 9, 478. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Li, Y.; Han, Z.; Li, Z. Morphological, structural and physiological differences in heteromorphic leaves of Euphrates poplar during development stages and at crown scales. Plant Biol. 2020, 22, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Oda, A.; Kenzo, T.; Kashimura, S.; Ninomiya, I.; Wang, L.; Yoshikawa, K.; Fukuda, K. Physiological and morphological differences in the heterophylly of Sabina vulgaris Ant. in the semi-arid environment of Mu Us Desert, Inner Mongolia, China. J. Arid Environ. 2010, 74, 43–48. [Google Scholar] [CrossRef]
- Hamilton, M.; Tilyard, P.; Williams, D.; Vaillancourt, R.; Wardlaw, T.; Potts, B. The genetic variation in the timing of heteroblastic transition in Eucalyptus globulus is stable across environments. Aust. J. Bot. 2011, 59, 170–175. [Google Scholar] [CrossRef]
- Deschamp, P.A.; Cooke, T.J. Causal mechanisms of leaf dimorphism in the aquatic angiosperm Callitriche heterophylla. Am. J. Bot. 1984, 71, 319–329. [Google Scholar] [CrossRef]
- Nakayama, H.; Nakayama, N.; Seiki, S.; Kojima, M.; Sakakibara, H.; Sinha, N.; Kimura, S. Regulation of the KNOX-GA gene module induces heterophyllic alteration in North American lake cress. Plant Cell 2014, 26, 4733–4748. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, S.; Yang, J.; Schultz, E.A.; Clarke, K.; Hou, H. Water-Wisteria as an ideal plant to study heterophylly in higher aquatic plants. Plant Cell Rep. 2017, 36, 1225–1236. [Google Scholar] [CrossRef]
- Iqbal, A.; Wang, T.; Wu, G.; Tang, W.; Zhu, C.; Wang, D.; Li, Y.; Wang, H. Physiological and transcriptome analysis of heteromorphic leaves and hydrophilic roots in response to soil drying in desert Populus euphratica. Sci. Rep. 2017, 7, 12188. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, W.; Li, Y.; Zhang, X.; Bai, X.; Niu, Z.; Zhang, X.; Li, Z.; Wan, D. Transcriptomic analysis of seed germination under salt stress in two desert sister species (Populus euphratica and P. pruinosa). Front. Genet. 2019, 10, 231. [Google Scholar] [CrossRef]
- Webster, C.; Figueroa-Corona, L.; Méndez-González, I.D.; Álvarez-Soto, L.; Neale, D.B.; Jaramillo-Correa, J.P.; Wegrzyn, J.L.; Vázquez-Lobo, A. Comparative analysis of differential gene expression indicates divergence in ontogenetic strategies of leaves in two conifer genera. Ecol. Evol. 2022, 12, e8611. [Google Scholar] [CrossRef]
- He, D.; Guo, P.; Gugger, P.F.; Guo, Y.; Liu, X.; Chen, J. Investigating the molecular basis for heterophylly in the aquatic plant Potamogeton octandrus (Potamogetonaceae) with comparative transcriptomics. PeerJ 2018, 6, e4448. [Google Scholar] [CrossRef]
- Wu, Z.; Jiang, Z.; Li, Z.; Jiao, P.; Zhai, J.; Liu, S.; Han, X.; Zhang, S.; Sun, J.; Gai, Z. Multi-omics analysis reveals spatiotemporal regulation and function of heteromorphic leaves in Populus. Plant Physiol. 2023, 192, 188–204. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Chen, G.; Li, M.; Liu, M.; Liu, D. Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages. PLoS ONE 2015, 10, e0137701. [Google Scholar] [CrossRef]
- Zhai, J.; Li, Z.; Si, J.; Zhang, S.; Han, X.; Chen, X. Structural and functional responses of the heteromorphic leaves of different tree heights on Populus euphratica oliv. to different soil moisture conditions. Plants 2022, 11, 2376. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Pang, M.; Zhu, C.; Bi, Z. Comparison of drought resistance of anatomical structure in the heterophylly mechanism of Sabina vulgaris. Acta Bot. Boreal.-Occident. Sin. 2017, 37, 1756–1763. [Google Scholar] [CrossRef]
- Wells, C.L.; Pigliucci, M. Adaptive phenotypic plasticity: The case of heterophylly in aquatic plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 1–18. [Google Scholar] [CrossRef]
- Mommer, L.; Visser, E.J. Underwater photosynthesis in flooded terrestrial plants: A matter of leaf plasticity. Ann. Bot. 2005, 96, 581–589. [Google Scholar] [CrossRef]
- Brodribb, T.; Hill, R. A Physiological Comparison of Leaves and Phyllodes in Acacia melanoxylon. Aust. J. Bot. 1993, 41, 293–305. [Google Scholar] [CrossRef]
- Searle, S. Acacia melanoxylon—A review of variation among planted trees. Aust. For. 2000, 63, 79–85. [Google Scholar] [CrossRef]
- Jennings, S.; Tasmania, F. Managing native forest for Blackwood (Acacia melanoxylon) production in north western Tasmania. Aust. For. 1998, 61, 141–146. [Google Scholar] [CrossRef]
- Costermans, L.F. Native trees and shrubs of south-eastern Australia. In Native Trees and Shrubs of South-Eastern Australia; Rigby Publishers Ltd.: London, UK, 1981. [Google Scholar]
- Forster, M.A.; Bonser, S.P. Heteroblastic development and the optimal partitioning of traits among contrasting environments in Acacia implexa. Ann. Bot. 2009, 103, 95–105. [Google Scholar] [CrossRef]
- Dong, Z.; He, H. Phyllode anatomy and histochemistry of four Acacia species (Leguminosae: Mimosoideae) in the Great Sandy Desert, north-western Australia. J. Arid Environ. 2017, 139, 110–120. [Google Scholar] [CrossRef]
- Boughton, V.H. Phyllode structure, taxonomy and distribution in some Australian Acacias. Aust. J. Bot. 1986, 34, 663–674. [Google Scholar] [CrossRef]
- Pedley, L. Derivation and dispersal of Acacia (Leguminosae), with particular reference to Australia, and the recognition of Senegalia and Racosperma. Bot. J. Linn. Soc. 1986, 92, 219–254. [Google Scholar] [CrossRef]
- Forster, M.A.; Ladd, B.; Bonser, S.P. Optimal allocation of resources in response to shading and neighbours in the heteroblastic species, Acacia implexa. Ann. Bot. 2011, 107, 219–228. [Google Scholar] [CrossRef]
- Rose, K.M.; Mickelbart, M.V.; Jacobs, D.F. Plasticity of phenotype and heteroblasty in contrasting populations of Acacia koa. Ann. Bot. 2019, 124, 399–409. [Google Scholar] [CrossRef]
- Walters, G.A.; Bartholomew, D.P. Adaptation of Acacia koa leaves and phyllodes to changes in photosynthetic photon flux density. For. Sci. 1990, 36, 1050–1060. [Google Scholar] [CrossRef]
- Freschet, G.T.; Violle, C.; Bourget, M.Y.; Scherer-Lorenzen, M.; Fort, F. Allocation, morphology, physiology, architecture: The multiple facets of plant above-and below-ground responses to resource stress. New Phytol. 2018, 219, 1338–1352. [Google Scholar] [CrossRef]
- Walters, G.A.; Bartholomew, D.P. Acacia koa leaves and phyllodes gas exchange, morphological, anatomical, and biochemical characteristics. Bot Gaz 1984, 145, 351–357. [Google Scholar] [CrossRef]
- Fahmy, G.M. Leaf anatomy and its relation to the ecophysiology of some non-succulent desert plants from Egypt. J. Arid Environ. 1997, 36, 499–525. [Google Scholar] [CrossRef]
- Jenks, M.A.; Ashworth, E.N. Plant epiculticular waxes: Function, production, and genetics. Hortic. Rev. 1999, 23, 1–68. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef]
- Jordan, G.J.; Dillon, R.A.; Weston, P.H. Solar radiation as a factor in the evolution of scleromorphic leaf anatomy in Proteaceae. Am. J. Bot. 2005, 92, 789–796. [Google Scholar] [CrossRef]
- Zeng, M.; He, S.; Hao, L.; Li, Y.; Zheng, C.; Zhao, Y. Conjoint analysis of genome-wide lncRNA and mRNA expression of heteromorphic leavesin response to environmental heterogeneityin Populus euphratica. Int. J. Mol. Sci. 2019, 20, 5148. [Google Scholar] [CrossRef]
- Wang, H.; Wu, F.; Li, M.; Zhu, X.; Shi, C.; Shao, C.; Ding, G. Structure and chlorophyll fluorescence of heteroblastic foliage affect first-year growth in Pinus massoniana Lamb. seedlings. Plant Physiol. Biochem. 2022, 170, 206–217. [Google Scholar] [CrossRef]
- Wang, H.; Wu, F.; Wu, C.; Yu, S.; Zhu, X.; Xie, W. Growth and physiological responses of seedlings with different leaf shapes to drought and re-watering in Pinus massoniana. J. Northeast For. Univ. 2018, 46, 1–6. [Google Scholar] [CrossRef]
- Rüggeberg, M.; Speck, T.; Paris, O.; Lapierre, C.; Burgert, I. Stiffness gradients in vascular bundles of the palm Washingtonia robusta. Proc. R. Soc. B Biol. Sci. 2008, 275, 2221–2229. [Google Scholar] [CrossRef]
- Ye, Z.-P.; Suggett, D.J.; Robakowski, P.; Kang, H.-J. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Buckley, T.N.; Diaz-Espejo, A. Partitioning changes in photosynthetic rate into contributions from different variables. Plant Cell Environ. 2015, 38, 1200–1211. [Google Scholar] [CrossRef]
- Zhang, J.; De-oliveira-Ceciliato, P.; Takahashi, Y.; Schulze, S.; Dubeaux, G.; Hauser, F.; Azoulay-Shemer, T.; Tõldsepp, K.; Kollist, H.; Rappel, W.-J. Insights into the molecular mechanisms of CO2-mediated regulation of stomatal movements. Curr. Biol. 2018, 28, R1356–R1363. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhao, F.; Zhang, H.; Zhu, Z. Responses of the growth, photosynthetic characteristics, endogenous hormones and antioxidant activity of Carpinus betulus seedlings to different light intensities. Front. Plant Sci. 2022, 13, 1055984. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Fukamachi, H.; Hidaka, T. Photosynthetic performance of Vigna radiata leaves developed at different temperature and irradiance levels. Plant Sci. 2003, 164, 451–458. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Vetoshkina, D.V.; Marenkova, T.V.; Borisova-Mubarakshina, M.M. Antioxidants of Non-Enzymatic Nature: Their function in higher plant cells and the ways of boosting their Biosynthesis. Antioxidants 2023, 12, 2014. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.M.; Ji, J.; Yue, J.Y.; Shi, S.Q.; Chang, E.M. Exogenous Abscisic Acid Modulates Reactive Oxygen Metabolism and Related Gene Expression in Platycladus orientalis under H2O2-Induced Stress. Russ. J. Plant Physiol. 2020, 67, 85–93. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Zhang, G.; Chen, Y.; Gao, J.; Sun, Y.-R.; Sun, M.-F.; Chen, J.-P. Exogenous application of gibberellic acid and ascorbic acid improved tolerance of okra seedlings to NaCl stress. Acta Physiol. Plant. 2019, 41, 93. [Google Scholar] [CrossRef]
- Chang, Z.; Liu, Y.; Dong, H.; Teng, K.; Han, L.; Zhang, X. Effects of cytokinin and nitrogen on drought tolerance of creeping bentgrass. PLoS ONE 2016, 11, e0154005. [Google Scholar] [CrossRef]
- Islam, M.R.; Rahman, M.M.; Mohi-Ud-Din, M.; Akter, M.; Zaman, E.; Keya, S.S.; Hasan, M.; Hasanuzzaman, M. Cytokinin and gibberellic acid-mediated waterlogging tolerance of mungbean (Vigna radiata L. Wilczek). PeerJ 2022, 10, e12862. [Google Scholar] [CrossRef]
- Ooi, A.; Lemtiri-Chlieh, F.; Wong, A.; Gehring, C. Direct modulation of the guard cell outward-rectifying potassium channel (GORK) by Abscisic Acid. Mol. Plant 2017, 10, 1469–1472. [Google Scholar] [CrossRef]
- Schulze, S.; Dubeaux, G.; Ceciliato, P.H.O.; Munemasa, S.; Nuhkat, M.; Yarmolinsky, D.; Aguilar, J.; Diaz, R.; Azoulay-Shemer, T.; Steinhorst, L.; et al. A role for calcium-dependent protein kinases in differential CO2 and ABA controlled stomatal closing and low CO2 induced stomatal opening in Arabidopsis. New Phytol. 2021, 229, 2765–2779. [Google Scholar] [CrossRef]
- Yruela, I. Transition metals in plant photosynthesis. Metallomics 2013, 5, 1090–1109. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Physiological and molecular responses to magnesium nutritional imbalance in plants. Plant Soil 2013, 368, 87–99. [Google Scholar] [CrossRef]
- Carrio-Segui, N.; Romero, P.; Sanz, A.; Penarrubia, L. Interaction Between ABA Signaling and Copper Homeostasis in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 1568–1582. [Google Scholar] [CrossRef]
- Wild, M.; Daviere, J.-M.; Regnault, T.; Sakvarelidze-Achard, L.; Carrera, E.; Diaz, I.L.; Cayrel, A.; Dubeaux, G.; Vert, G.; Achard, P. Tissue-Specific Regulation of Gibberellin Signaling Fine-Tunes Arabidopsis Iron-Deficiency Responses. Dev. Cell 2016, 37, 190–200. [Google Scholar] [CrossRef]
- Guo, W.; Cong, Y.; Hussain, N.; Wang, Y.; Liu, Z.; Jiang, L.; Liang, Z.; Chen, K. The Remodeling of Seedling Development in Response to Long-Term Magnesium Toxicity and Regulation by ABA-DELLA Signaling in Arabidopsis. Plant Cell Physiol. 2014, 55, 1713–1726. [Google Scholar] [CrossRef] [PubMed]
- Koenig, D.; Bayer, E.; Kang, J.; Kuhlemeier, C.; Sinha, N. Auxin patterns Solanum lycopersicum leaf morphogenesis. Development 2009, 136, 2997–3006. [Google Scholar] [CrossRef] [PubMed]
- Shani, E.; Ben-Gera, H.; Shleizer-Burko, S.; Burko, Y.; Weiss, D.; Ori, N. Cytokinin Regulates Compound Leaf Development in Tomato. Plant Cell 2010, 22, 3206–3217. [Google Scholar] [CrossRef] [PubMed]
- Borchert, R. Gibberellic acid and rejuvenation of apical meristems in acacia melanoxylon. Naturwissenschaften 1965, 52, 65–66. [Google Scholar] [CrossRef]
- Savvides, A.; Fanourakis, D.; van Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.L. Kjeldahl method for total nitrogen. Anal. Chem. 1950, 22, 354–358. [Google Scholar] [CrossRef]
- Yuan, G.; Lavkulich, L. Colorimetric determination of phosphorus in citrate-bicarbonate-dithionite extracts of soils. Commun. Soil Sci. Plant Anal. 1995, 26, 1979–1988. [Google Scholar] [CrossRef]
- Havre, G.N. The flame photometric determination of sodium, potassium and calcium in plant extracts with special reference to interference effects. Anal. Chim. Acta 1961, 25, 557–566. [Google Scholar] [CrossRef]
- Kiliç, Z.; Acar, O.; Ulaşan, M.; Ilim, M. Determination of lead, copper, zinc, magnesium, calcium and iron in fresh eggs by atomic absorption spectrometry. Food Chem. 2002, 76, 107–116. [Google Scholar] [CrossRef]
- Dible, W.; Truog, E.; Berger, K. Boron determination in soils and plants. Anal. Chem. 1954, 26, 418–421. [Google Scholar] [CrossRef]
- Soest, P.V.; Wine, R. Use of detergents in the analysis of fibrous feeds. IV. Determination of plant cell-wall constituents. J. Assoc. Off. Anal. Chem. 1967, 50, 50–55. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B- and ozone-induced biochemical changes in antioxidant enzymes of Arab. Thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of chlorophyll a, chlorophyll b, chlorophyll a+ b, and total carotenoid content in eucalyptus leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Biomass (g) | Stem Biomass (g) | Leaf Biomass (g) | Total Biomass (g) | R/S | CL/T | P/T |
|---|---|---|---|---|---|---|---|
| Cont. | 0.94 ± 0.05 a | 0.35 ± 0.04 a | 1.04 ± 0.18 a | 2.34 ± 0.15 a | 0.67 ± 0.08 a | 0.24 ± 0.05 b | 0.21 ± 0.02 a |
| WL | 0.16 ± 0.005 b | 0.11 ± 0.03 b | 0.76 ± 0.14 b | 1.02 ± 0.14 b | 0.19 ± 0.04 b | 0.74 ± 0.04 a | 0.00 ± 0.00 b |
| Leaf Type | Stomatal Length (μm) | Stomatal Width (μm) | Stomatal Aperture Length (μm) | Stomatal Aperture Width (μm) | Stomatal Density (number·mm−2) |
|---|---|---|---|---|---|
| Cont. | 14.75 ± 1.10 a | 9.07 ± 0.53 a | 2.78 ± 1.09 b | 0.91 ± 0.36 b | 660 ± 70.25 a |
| WL | 15.46 ± 2.54 a | 6.87 ± 1.16 b | 6.0 ± 1.03 a | 1.41 ± 0.27 a | 272 ± 35.12 b |
| Leaf Type | P′max (μmol·m−2·s−1) | LSP (μmol·m−2·s−1) | LCP (μmol·m−2·s−1) | Rd (μmol·m−2·s−1) |
|---|---|---|---|---|
| Cont. | 18.76 ± 0.79 a | 1748.21 ± 241.59 a | 51.48 ± 27.3 a | 3.54 ± 2.63 a |
| WL | 10.56 ± 1.14 b | 1509.91 ± 144.77 a | 25.68 ± 6.04 b | 1.28 ± 0.32 b |
| Leaf Type | Ci (µmol·mol−1) | Tr (mol·m−2·s−1) | Gs (mol·m−2·s−1) | WUE (μmol·mol−1) |
| Cont. | 174.95 ± 46.66 b | 0.0042 ± 0.0006 a | 0.159 ± 0.03 a | 4488.87 ± 519.35 a |
| WL | 245.86 ± 15.89 a | 0.0038 ± 0.0005 a | 0.138 ± 0.02 a | 2762.18 ± 502.68 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Chen, Z.; Chen, M.; Zeng, B.; Li, X.; Tu, P.; Hu, B. Morphological, Anatomical, and Physiological Characteristics of Heteroblastic Acacia melanoxylon Grown under Weak Light. Plants 2024, 13, 870. https://doi.org/10.3390/plants13060870
Bai X, Chen Z, Chen M, Zeng B, Li X, Tu P, Hu B. Morphological, Anatomical, and Physiological Characteristics of Heteroblastic Acacia melanoxylon Grown under Weak Light. Plants. 2024; 13(6):870. https://doi.org/10.3390/plants13060870
Chicago/Turabian StyleBai, Xiaogang, Zhaoli Chen, Mengjiao Chen, Bingshan Zeng, Xiangyang Li, Panfeng Tu, and Bing Hu. 2024. "Morphological, Anatomical, and Physiological Characteristics of Heteroblastic Acacia melanoxylon Grown under Weak Light" Plants 13, no. 6: 870. https://doi.org/10.3390/plants13060870