
The Role of Soil Microbial Consortia in Sustainable Cereal Crop Residue Management



Abstract
:1. Introduction
2. Estimates of Cereal Crop Residue Production in Lithuania and Northern Europe
3. Decomposition Dynamics of Cereal Crop Residues
4. Agricultural Practices Enhance the Development of Microbial Communities Associated with Crop Residue Decomposition
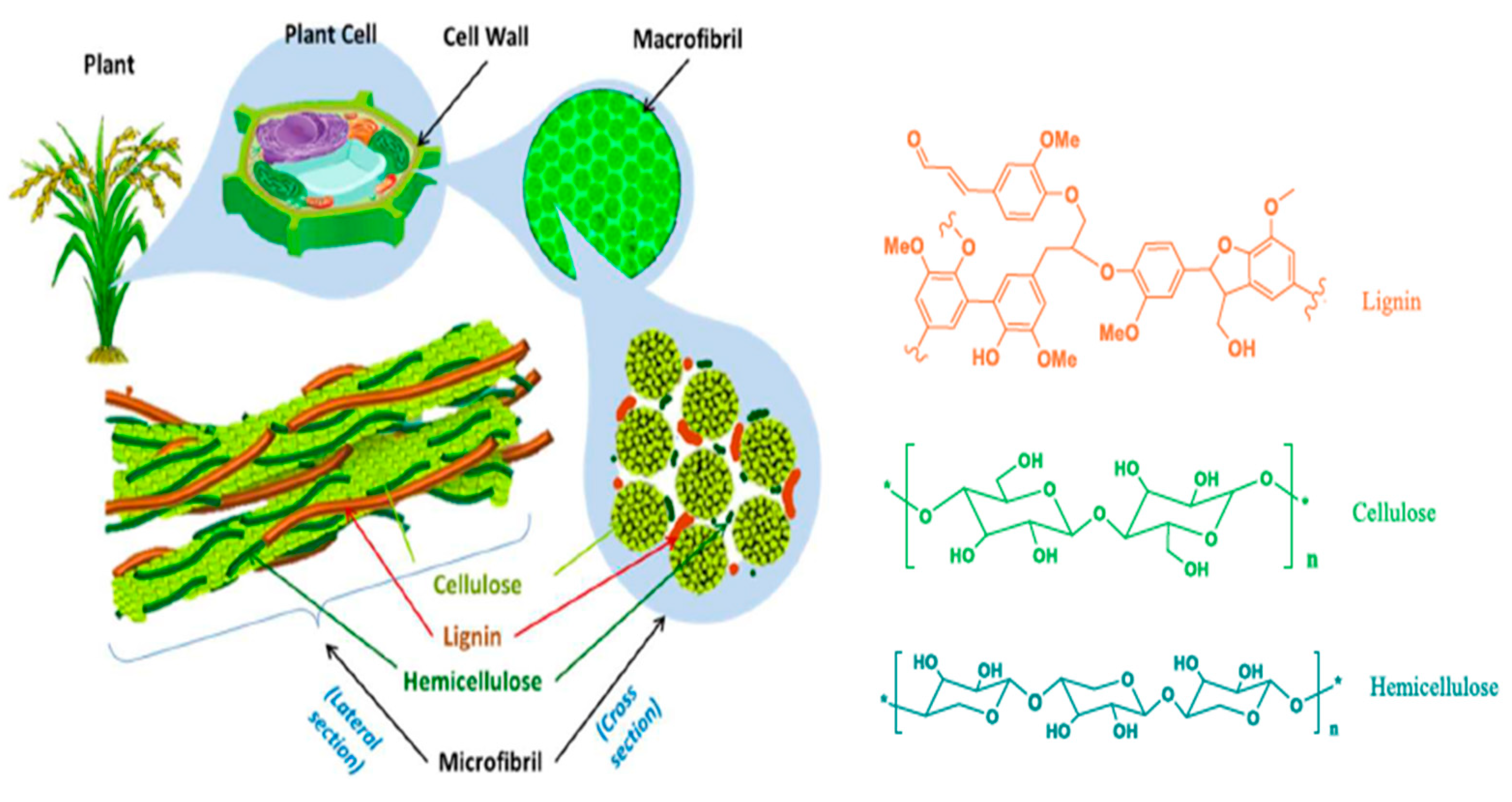
5. Potential Utilization of Lignocellulolytic Microorganisms in Cereal Crop Residue Decomposition
5.1. Lignocellulolytic Activity of Fungi
5.2. Lignocellulolytic Activity of Bacteria
5.3. Lignocellulolytic Activity of Actinobacteria
6. Research Gaps and Future Directions
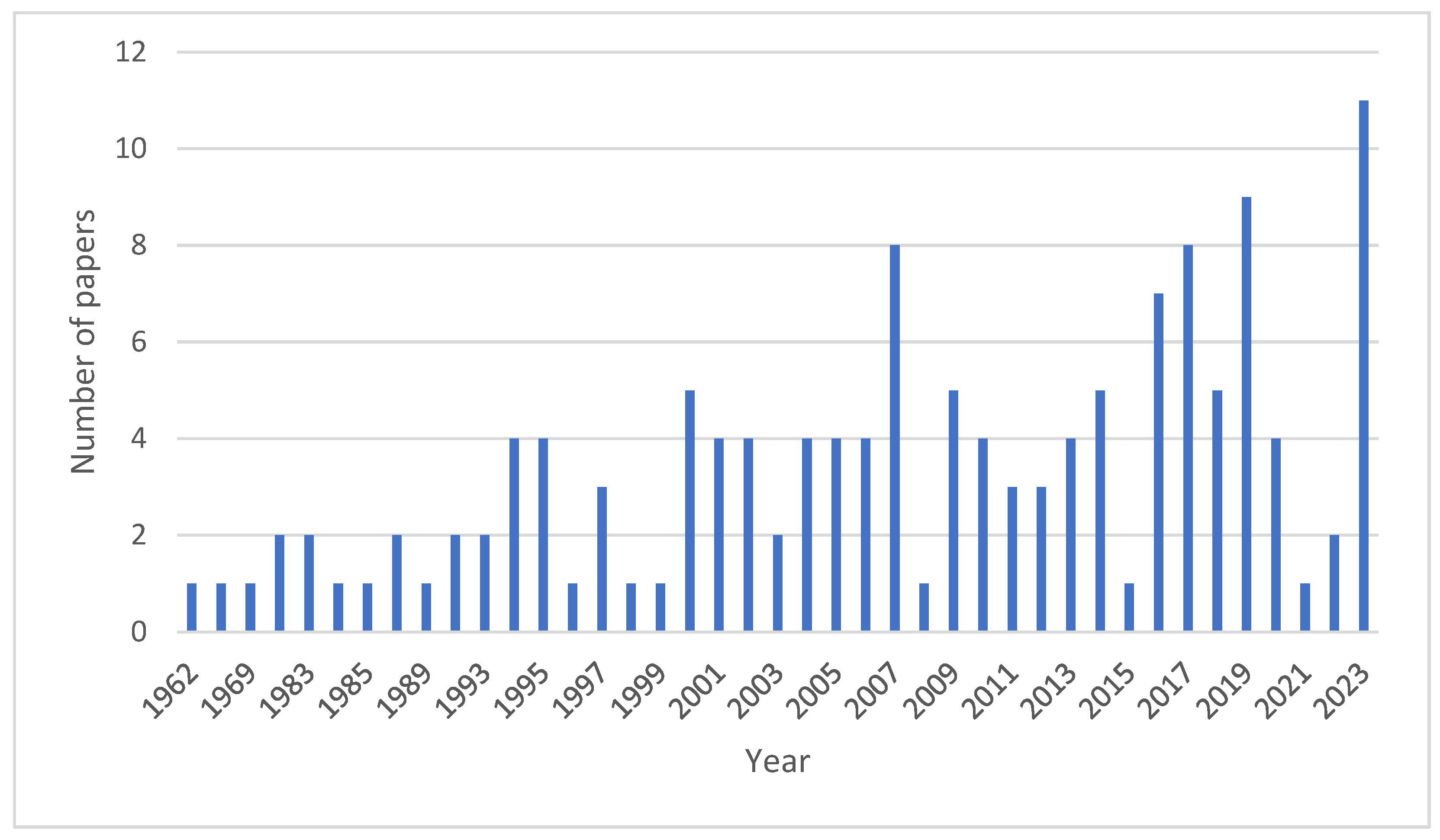
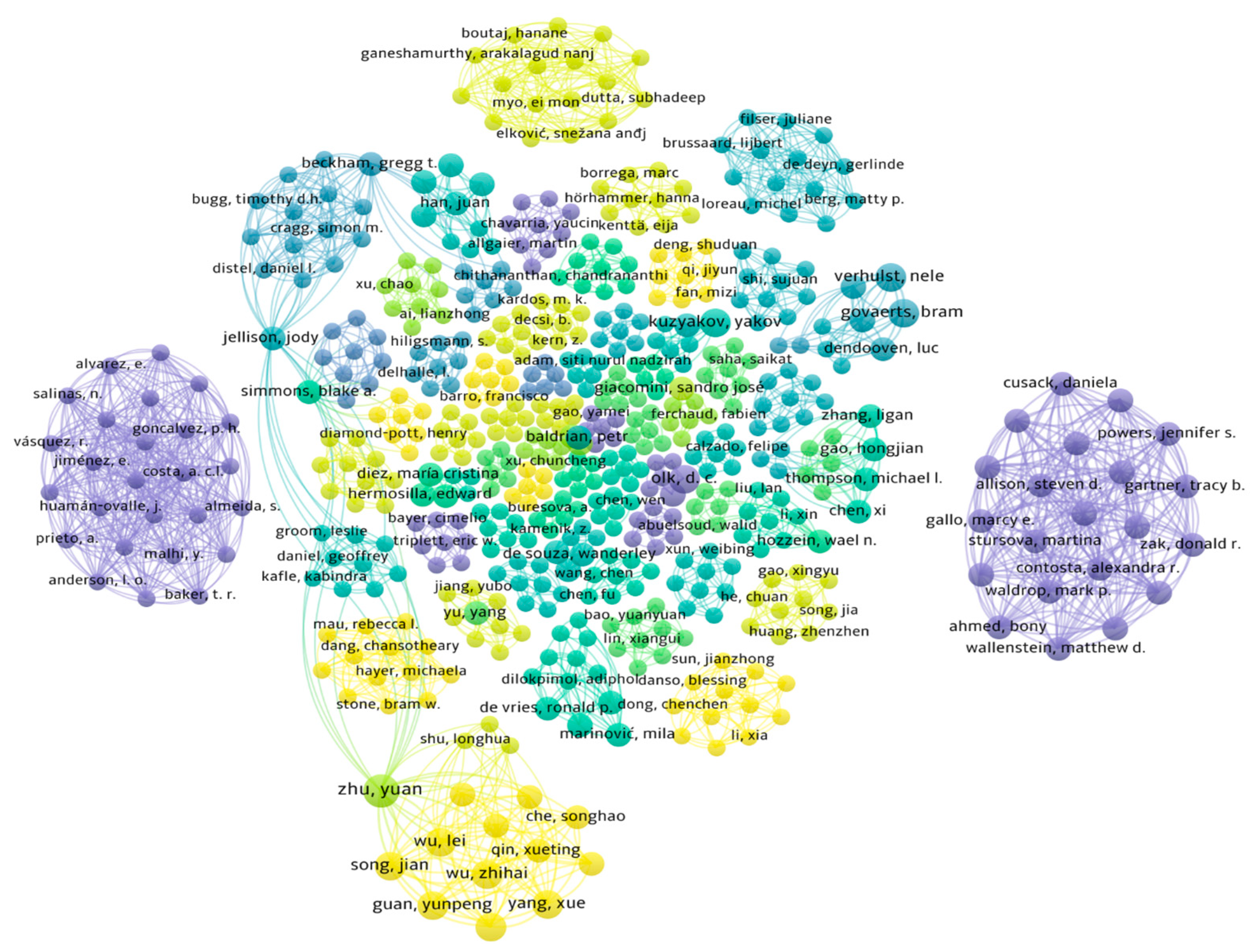
7. Materials and Methods
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lai, R. Conservation Tillage for Sustainable Agriculture: Tropics Versus Temperate Environments. Adv. Agron. 1989, 42, 85–197. [Google Scholar] [CrossRef]
- Bisen, N.; Rahangdale, C.P. Crop Residues Management Option for Sustainable Soil Health in Rice-Wheat System: A Review. Int. J. Chem. Stud. 2017, 5, 1038–1042. [Google Scholar]
- Yadvinder-Singh, B.S.; Timsina, J. Crop Residue Management for Nutrient Cycling and Improving Soil Productivity in Rice-Based Cropping Systems in the Tropics. Adv. Agron. 2005, 85, 269–407. [Google Scholar] [CrossRef]
- Triplett, G.B.; Dick, W.A. No-Tillage Crop Production: A Revolution in Agriculture! Agron. J. 2008, 100, 153–165. [Google Scholar] [CrossRef]
- The United Nations Statistics Division. Standard Country or Area Codes for Statistical Use (M49); The United Nations Statistics Division: New York, NY, USA, 2018. [Google Scholar]
- Pinke, Z.; Decsi, B.; Jámbor, A.; Kardos, M.K.; Kern, Z.; Kozma, Z.; Ács, T. Climate Change and Modernization Drive Structural Realignments in European Grain Production. Sci. Rep. 2022, 12, 7374. [Google Scholar] [CrossRef] [PubMed]
- Turmel, M.S.; Speratti, A.; Baudron, F.; Verhulst, N.; Govaerts, B. Crop Residue Management and Soil Health: A Systems Analysis. Agric. Syst. 2015, 134, 6–16. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. Crop Residue Removal Impacts on Soil Productivity and Environmental Quality. CRC Crit. Rev. Plant Sci. 2009, 28, 140–160. [Google Scholar] [CrossRef]
- Wang, J.; Sainju, U.M. Aggregate-Associated Carbon and Nitrogen Affected by Residue Placement, Crop Species, and Nitrogen Fertilization. Soil Sci. 2014, 179, 153–165. [Google Scholar] [CrossRef]
- Smith, W.N.; Grant, B.B.; Campbell, C.A.; McConkey, B.G.; Desjardins, R.L.; Kröbel, R.; Malhi, S.S. Crop Residue Removal Effects on Soil Carbon: Measured and Inter-Model Comparisons. Agric. Ecosyst. Environ. 2012, 161, 27–38. [Google Scholar] [CrossRef]
- He, Y.; Presley, D.A.R.; Tatarko, J.; Blanco-Canqui, H. Crop Residue Harvest Impacts Wind Erodibility and Simulated Soil Loss in the Central Great Plains. GCB Bioenergy 2018, 10, 213–226. [Google Scholar] [CrossRef]
- Chowdhury, S.; Farrell, M.; Butler, G.; Bolan, N. Assessing the Effect of Crop Residue Removal on Soil Organic Carbon Storage and Microbial Activity in a No-till Cropping System. Soil Use Manag. 2015, 31, 450–460. [Google Scholar] [CrossRef]
- Rasche, F.; Musyoki, M.K.; Röhl, C.; Muema, E.K.; Vanlauwe, B.; Cadisch, G. Lasting Influence of Biochemically Contrasting Organic Inputs on Abundance and Community Structure of Total and Proteolytic Bacteria in Tropical Soils. Soil Biol. Biochem. 2014, 74, 204–213. [Google Scholar] [CrossRef]
- Garg, S. Bioremediation of Agricultural, Municipal, and Industrial Wastes. In Waste Management: Concepts, Methodologies, Tools, and Applications; IGI Global: Hershey, PA, USA, 2019. [Google Scholar] [CrossRef]
- Goswami, S.B.; Mondal, R.; Mandi, S.K. Crop Residue Management Options in Rice–Rice System: A Review. Arch. Agron. Soil Sci. 2020, 66, 1218–1234. [Google Scholar] [CrossRef]
- Payne, C.M.; Knott, B.C.; Mayes, H.B.; Hansson, H.; Himmel, M.E.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T. Fungal Cellulases. Chem. Rev. 2015, 115, 1308–1448. [Google Scholar] [CrossRef]
- Madeira, J.V.; Contesini, F.J.; Calzado, F.; Rubio, M.V.; Zubieta, M.P.; Lopes, D.B.; de Melo, R.R. Agro-Industrial Residues and Microbial Enzymes. In Biotechnology of Microbial Enzymes; Academic Press: Cambridge, MA, USA, 2017; pp. 475–511. [Google Scholar] [CrossRef]
- Shamshitov, A.; Decorosi, F.; Viti, C.; Fornasier, F.; Kadžienė, G.; Supronienė, S. Characterisation of Cellulolytic Bacteria Isolated from Agricultural Soil in Central Lithuania. Sustainability 2023, 15, 598. [Google Scholar] [CrossRef]
- FAO. FAOSTAT. Available online: https://www.fao.org/faostat/en/data/QCL (accessed on 10 July 2023).
- Hakala, K.; Kontturi, M.; Pahkala, K. Field Biomass as Global Energy Source. Agric. Food Sci. 2009, 18, 347–365. [Google Scholar] [CrossRef]
- Almagro, M.; Ruiz-Navarro, A.; Díaz-Pereira, E.; Albaladejo, J.; Martínez-Mena, M. Plant Residue Chemical Quality Modulates the Soil Microbial Response Related to Decomposition and Soil Organic Carbon and Nitrogen Stabilization in a Rainfed Mediterranean Agroecosystem. Soil Biol. Biochem. 2021, 156, 108198. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Del Galdo, I.; Piermatteo, D. Litter Decomposition: Concepts, Methods and Future Perspectives. In Soil Carbon Dynamics: An Integrated Methodology; Cambridge University Press: Cambridge, UK, 2010; pp. 76–90. [Google Scholar] [CrossRef]
- Kumar, K.; Goh, K.M. Crop Residues and Management Practices: Effects on Soil Quality, Soil Nitrogen Dynamics, Crop Yield, and Nitrogen Recovery. Adv. Agron. 1999, 68, 197–319. [Google Scholar] [CrossRef]
- Liira, J.; Sepp, T.; Parrest, O. The Forest Structure and Ecosystem Quality in Conditions of Anthropogenic Disturbance along Productivity Gradient. For. Ecol. Manag. 2007, 250, 34–46. [Google Scholar] [CrossRef]
- Chapman, S.K.; Koch, G.W. What Type of Diversity Yields Synergy during Mixed Litter Decomposition in a Natural Forest Ecosystem? Plant Soil 2007, 299, 153–162. [Google Scholar] [CrossRef]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and Cellulose Degradation and Nitrogen Dynamics during Decomposition of Three Leaf Litter Species in a Mediterranean Ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Shinde, R.; Shahi, D.K.; Mahapatra, P.; Naik, S.K.; Thombare, N.; Singh, A.K. Potential of Lignocellulose Degrading Microorganisms for Agricultural Residue Decomposition in Soil: A Review. J. Environ. Manag. 2022, 320, 115843. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Sampath Kumar, N.S.; Chintagunta, A.D. Bioethanol Production from Cereal Crops and Lignocelluloses Rich Agro-Residues: Prospects and Challenges. SN Appl. Sci. 2020, 2, 1673. [Google Scholar] [CrossRef]
- Naomi, R.; Idrus, R.B.H.; Fauzi, M.B. Plant-vs. Bacterial-Derived Cellulose for Wound Healing: A Review. Int. J. Environ. Res. Public Health 2020, 18, 6803. [Google Scholar] [CrossRef]
- Kumar Gupta, P.; Sai Raghunath, S.; Venkatesh Prasanna, D.; Venkat, P.; Shree, V.; Chithananthan, C.; Choudhary, S.; Surender, K.; Geetha, K. An Update on Overview of Cellulose, Its Structure and Applications. In Cellulose; Books on Demand: Norderstedt, Germany, 2019. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Rao, J.; Lv, Z.; Chen, G.; Peng, F. Hemicellulose: Structure, Chemical Modification, and Application. Prog. Polym. Sci. 2023, 140, 101675. [Google Scholar] [CrossRef]
- Balakshin, M.Y.; Capanema, E.A.; Chang, H.M. Recent Advances in the Isolation and Analysis of Lignins and Lignin-Carbohydrate Complexes. In Characterization of Lignocellulosic Materials; Wiley Online Library: Hoboken, NJ, USA, 2009. [Google Scholar] [CrossRef]
- Chio, C.; Sain, M.; Qin, W. Lignin Utilization: A Review of Lignin Depolymerization from Various Aspects. Renew. Sustain. Energy Rev. 2019, 107, 232–249. [Google Scholar] [CrossRef]
- Di Gruttola, F.; Borello, D. Analysis of the Eu Secondary Biomass Availability and Conversion Processes to Produce Advanced Biofuels: Use of Existing Databases for Assessing a Metric Evaluation for the 2025 Perspective. Sustainability 2021, 14, 7882. [Google Scholar] [CrossRef]
- Puițel, A.C.; Suditu, G.D.; Danu, M.; Ailiesei, G.L.; Nechita, M.T. An Experimental Study on the Hot Alkali Extraction of Xylan-Based Hemicelluloses from Wheat Straw and Corn Stalks and Optimization Methods. Polymers 2022, 14, 1662. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zuo, S.; Niu, D.; Jiang, D.; Tao, Y.; Xu, C. Effect of Four Species of White Rot Fungi on the Chemical Composition and In Vitro Rumen Degradability of Naked Oat Straw. Waste Biomass Valorization 2021, 12, 435–443. [Google Scholar] [CrossRef]
- Kamusoko, R.; Jingura, R.M.; Parawira, W.; Chikwambi, Z. Characterization of Lignocellulosic Crop Residues for Potential Biogas Production in Zimbabwe. Biofuels Bioprod. Biorefin. 2022, 16, 1165–1171. [Google Scholar] [CrossRef]
- Rencoret, J.; Marques, G.; Rosado, M.J.; Benito, J.; Barro, F.; Gutiérrez, A.; del Río, J.C. Variations in the Composition and Structure of the Lignins of Oat (Avena sativa L.) Straws According to Variety and Planting Season. Int. J. Biol. Macromol. 2023, 242, 124811. [Google Scholar] [CrossRef]
- Sun, R.C.; Fang, J.M.; Tomkinson, J. Characterization and Esterification of Hemicelluloses from Rye Straw. J. Agric. Food Chem. 2000, 48, 1247–1252. [Google Scholar] [CrossRef]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic Biomass: A Sustainable Platform for the Production of Bio-Based Chemicals and Polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef]
- Hernández-Beltrán, J.U.; Hernández-De Lira, I.O.; Cruz-Santos, M.M.; Saucedo-Luevanos, A.; Hernández-Terán, F.; Balagurusamy, N. Insight into Pretreatment Methods of Lignocellulosic Biomass to Increase Biogas Yield: Current State, Challenges, and Opportunities. Appl. Sci. 2019, 18, 3721. [Google Scholar] [CrossRef]
- Borrega, M.; Hinkka, V.; Hörhammer, H.; Kataja, K.; Kenttä, E.; Ketoja, J.A.; Palmgren, R.; Salo, M.; Sundqvist-Andberg, H.; Tanaka, A. Utilizing and Valorizing Oat and Barley Straw as an Alternative Source of Lignocellulosic Fibers. Materials 2022, 15, 7826. [Google Scholar] [CrossRef]
- Weiler, D.A.; Bastos, L.M.; Schirmann, J.; Aita, C.; Giacomini, S.J. Changes in Chemical Composition of Cover Crops Residue during Decomposition. Cienc. Rural 2022, 52. [Google Scholar] [CrossRef]
- Chaves, B.; Redin, M.; Giacomini, S.J.; Schmatz, R.; Léonard, J.; Ferchaud, F.; Recous, S. The Combination of Residue Quality, Residue Placement and Soil Mineral N Content Drives C and N Dynamics by Modifying N Availability to Microbial Decomposers. Soil Biol. Biochem. 2021, 163, 108434. [Google Scholar] [CrossRef]
- Aragão, L.E.O.C.; Malhi, Y.; Metcalfe, D.B.; Silva-Espejo, J.E.; Jiménez, E.; Navarrete, D.; Almeida, S.; Costa, A.C.L.; Salinas, N.; Phillips, O.L.; et al. Above- and below-Ground Net Primary Productivity across Ten Amazonian Forests on Contrasting Soils. Biogeosciences 2009, 6, 2759–2778. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter Decomposition in Forest Ecosystems: A Review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Bao, Y.; Feng, Y.; Stegen, J.C.; Wu, M.; Chen, R.; Liu, W.; Zhang, J.; Li, Z.; Lin, X. Straw Chemistry Links the Assembly of Bacterial Communities to Decomposition in Paddy Soils. Soil Biol. Biochem. 2020, 148, 107866. [Google Scholar] [CrossRef]
- Filser, J.; Faber, J.H.; Tiunov, A.V.; Brussaard, L.; Frouz, J.; De Deyn, G.; Uvarov, A.V.; Berg, M.P.; Lavelle, P.; Loreau, M.; et al. Soil Fauna: Key to New Carbon Models. SOIL 2016, 2, 565–582. [Google Scholar] [CrossRef]
- Begum, N.; Guppy, C.; Herridge, D.; Schwenke, G. Influence of Source and Quality of Plant Residues on Emissions of N2O and CO2 from a Fertile, Acidic Black Vertisol. Biol. Fertil. Soils 2014, 50, 499–506. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Sources of CO2 Efflux from Soil and Review of Partitioning Methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric Controls on Carbon, Nitrogen, and Phosphorus Dynamics in Decomposing Litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef]
- McBride, S.G.; Choudoir, M.; Fierer, N.; Strickland, M.S. Volatile Organic Compounds from Leaf Litter Decomposition Alter Soil Microbial Communities and Carbon Dynamics. Ecology 2020, 101, e03130. [Google Scholar] [CrossRef] [PubMed]
- Grzyb, A.; Wolna-Maruwka, A.; Niewiadomska, A. Environmental Factors Affecting the Mineralization of Crop Residues. Agronomy 2020, 12, 1951. [Google Scholar] [CrossRef]
- Schoenau, J.J.; Campbell, C.A. Impact of Crop Residues on Nutrient Availability in Conservation Tillage Systems. Can. J. Plant Sci. 1996, 76, 621–626. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Studies in Ecology; Blackwell Scientific Publications: Oxford, UK, 1979; Volume 5. [Google Scholar]
- Bardgett, R.D.; Caruso, T. Soil Microbial Community Responses to Climate Extremes: Resistance, Resilience and Transitions to Alternative States. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190112. [Google Scholar] [CrossRef]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; McBratney, A.B. Soil Properties Drive Microbial Community Structure in a Large Scale Transect in South Eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef]
- Schloter, M.; Dilly, O.; Munch, J.C. Indicators for Evaluating Soil Quality. Agric. Ecosyst. Environ. 2003, 98, 255–262. [Google Scholar] [CrossRef]
- Hou, H.; Wang, C.; Ding, Z.; Zhang, S.; Yang, Y.; Ma, J.; Chen, F.; Li, J. Variation in the Soil Microbial Community of Reclaimed Land over Different Reclamation Periods. Sustainability 2018, 10, 2286. [Google Scholar] [CrossRef]
- Hartmann, M.; Six, J. Soil Structure and Microbiome Functions in Agroecosystems. Nat. Rev. Earth Environ. 2023, 4, 4–18. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, U.B.; Sahu, P.K.; Paul, S.; Kumar, A.; Malviya, D.; Singh, S.; Kuppusamy, P.; Singh, P.; Paul, D.; et al. Linking Soil Microbial Diversity to Modern Agriculture Practices: A Review. Int. J. Environ. Res. Public Health 2022, 5, 3141. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ni, T.; Xun, W.; Huang, X.; Huang, Q.; Ran, W.; Shen, B.; Zhang, R.; Shen, Q. Influence of Straw Incorporation with and without Straw Decomposer on Soil Bacterial Community Structure and Function in a Rice-Wheat Cropping System. Appl. Microbiol. Biotechnol. 2017, 101, 4761–4773. [Google Scholar] [CrossRef]
- Erickson, A.E. Tillage Effects on Soil Aeration. In Predicting Tillage Effects on Soil Physical Properties and Processes; Wiley Online Library: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Simmons, B.L.; Coleman, D.C. Microbial Community Response to Transition from Conventional to Conservation Tillage in Cotton Fields. Appl. Soil Ecol. 2008, 40, 518–528. [Google Scholar] [CrossRef]
- Fernandes, M.M.H.; da Silva, M.F.; Ferraudo, A.S.; Fernandes, C. Soil Structure under Tillage Systems with and without Cultivation in the Off-Season. Agric. Ecosyst. Environ. 2023, 342, 108237. [Google Scholar] [CrossRef]
- Khan, A.R. Influence of Tillage on Soil Aeration. J. Agron. Crop Sci. 1996, 177, 253–259. [Google Scholar] [CrossRef]
- Beyaert, R.P.; Paul Voroney, R. Estimation of Decay Constants for Crop Residues Measured over 15 Years in Conventional and Reduced Tillage Systems in a Coarse-Textured Soil in Southern Ontario. Can. J. Soil Sci. 2011, 91, 985–995. [Google Scholar] [CrossRef]
- Legrand, F.; Picot, A.; Cobo-Díaz, J.F.; Carof, M.; Chen, W.; Le Floch, G. Effect of Tillage and Static Abiotic Soil Properties on Microbial Diversity. Appl. Soil Ecol. 2018, 132, 135–145. [Google Scholar] [CrossRef]
- De Quadros, P.D.; Zhalnina, K.; Davis-Richardson, A.; Fagen, J.R.; Drew, J.; Bayer, C.; Camargo, F.A.O.; Triplett, E.W. The Effect of Tillage System and Crop Rotation on Soil Microbial Diversity and Composition in a Subtropical Acrisol. Diversity 2012, 4, 375. [Google Scholar] [CrossRef]
- Shanmugam, S.G.; Buehring, N.W.; Prevost, J.D.; Kingery, W.L. Soil Bacterial Community Diversity and Composition as Affected by Tillage Intensity Treatments in Corn-Soybean Production Systems. Microbiol. Res. 2021, 12, 157–172. [Google Scholar] [CrossRef]
- Duan, N.; Li, L.; Liang, X.; Fine, A.; Zhuang, J.; Radosevich, M.; Schaeffer, S.M. Variation in Bacterial Community Structure Under Long-Term Fertilization, Tillage, and Cover Cropping in Continuous Cotton Production. Front. Microbiol. 2022, 13, 847005. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.P.; Feng, Y.; Githinji, L.; Ankumah, R.; Balkcom, K.S. Impact of No-Tillage and Conventional Tillage Systems on Soil Microbial Communities. Appl. Environ. Soil Sci. 2012, 2012, 548620. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liu, Y.; Han, J.; Liao, Y. Conservation Tillage Enhances the Stability of the Rhizosphere Bacterial Community Responding to Plant Growth. Agron. Sustain. Dev. 2017, 37, 44. [Google Scholar] [CrossRef]
- Wang, Z.; Li, T.; Wen, X.; Liu, Y.; Han, J.; Liao, Y.; DeBruyn, J.M. Fungal Communities in Rhizosphere Soil under Conservation Tillage Shift in Response to Plant Growth. Front. Microbiol. 2017, 8, 1301. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Vashisht, M.; Singh, Y.; Thind, H.S. Soil Carbon Pools and Enzyme Activities in Aggregate Size Fractions after Seven Years of Conservation Agriculture in a Rice-Wheat System. Crop Pasture Sci. 2019, 70, 473–485. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, X.; Xu, Y.; Jin, M.; Ye, X.; Gao, H.; Chu, W.; Mao, J.; Thompson, M.L. Soil Labile Organic Carbon Fractions and Soil Enzyme Activities after 10 Years of Continuous Fertilization and Wheat Residue Incorporation. Sci. Rep. 2020, 10, 11318. [Google Scholar] [CrossRef]
- Sindelar, A.J.; Lamb, J.A.; Coulter, J.A. Short-Term Stover, Tillage, and Nitrogen Management Affect Near-Surface Soil Organic Matter. Soil Sci. Soc. Am. J. 2015, 79, 251–260. [Google Scholar] [CrossRef]
- Souza, R.C.; Cantão, M.E.; Vasconcelos, A.T.R.; Nogueira, M.A.; Hungria, M. Soil Metagenomics Reveals Differences under Conventional and No-Tillage with Crop Rotation or Succession. Appl. Soil Ecol. 2013, 72, 49–61. [Google Scholar] [CrossRef]
- Carbonetto, B.; Rascovan, N.; Álvarez, R.; Mentaberry, A.; Vázquez, M.P. Structure, Composition and Metagenomic Profile of Soil Microbiomes Associated to Agricultural Land Use and Tillage Systems in Argentine Pampas. PLoS ONE 2014, 9, e99949. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.; Schöler, A.; Ertl, J.; Xu, Z.; Schloter, M. Metagenomic Analyses Reveal No Differences in Genes Involved in Cellulose Degradation under Different Tillage Treatments. FEMS Microbiol. Ecol. 2015, 91, fiv069. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Villanueva, D.A.; Bello-López, J.M.; Navarro-Noya, Y.E.; Luna-Guido, M.; Verhulst, N.; Govaerts, B.; Dendooven, L. Bacterial Community Structure in Maize Residue Amended Soil with Contrasting Management Practices. Appl. Soil Ecol. 2015, 90, 49–59. [Google Scholar] [CrossRef]
- Chávez-Romero, Y.; Navarro-Noya, Y.E.; Reynoso-Martínez, S.C.; Sarria-Guzmán, Y.; Govaerts, B.; Verhulst, N.; Dendooven, L.; Luna-Guido, M. 16S Metagenomics Reveals Changes in the Soil Bacterial Community Driven by Soil Organic C, N-Fertilizer and Tillage-Crop Residue Management. Soil Tillage Res. 2016, 159, 1–8. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, J.E.; Hobbie, E.A. Microbes in Nature Are Limited by Carbon and Energy: The Starving-Survival Lifestyle in Soil and Consequences for Estimating Microbial Rates. Front. Microbiol. 2013, 4, 324. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Blagodatsky, S.; Khomyakov, N.; Myachina, O.; Kuzyakov, Y. Temperature Sensitivity and Enzymatic Mechanisms of Soil Organic Matter Decomposition along an Altitudinal Gradient on Mount Kilimanjaro. Sci. Rep. 2016, 6, 22240. [Google Scholar] [CrossRef]
- Knelman, J.E.; Graham, E.B.; Ferrenberg, S.; Lecoeuvre, A.; Labrado, A.; Darcy, J.L.; Nemergut, D.R.; Schmidt, S.K. Rapid Shifts in Soil Nutrients and Decomposition Enzyme Activity in Early Succession Following Forest Fire. Forests 2017, 8, 347. [Google Scholar] [CrossRef]
- Stott, D.E.; Andrews, S.S.; Liebig, M.A.; Wienhold, B.J.; Karlen, D.L. Evaluation of β-Glucosidase Activity as a Soil Quality Indicator for the Soil Management Assessment Framework. Soil Sci. Soc. Am. J. 2010, 74, 107–119. [Google Scholar] [CrossRef]
- Nevins, C.J.; Lacey, C.; Armstrong, S. The Synchrony of Cover Crop Decomposition, Enzyme Activity, and Nitrogen Availability in a Corn Agroecosystem in the Midwest United States. Soil Tillage Res. 2020, 197, 104518. [Google Scholar] [CrossRef]
- Chen, H.; Liang, Q.; Gong, Y.; Kuzyakov, Y.; Fan, M.; Plante, A.F. Reduced Tillage and Increased Residue Retention Increase Enzyme Activity and Carbon and Nitrogen Concentrations in Soil Particle Size Fractions in a Long-Term Field Experiment on Loess Plateau in China. Soil Tillage Res. 2019, 194, 104296. [Google Scholar] [CrossRef]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E.; et al. Lignocellulose Degradation Mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef]
- Kapahi, M.; Rani, R.; Kohli, K. Fungal Biorefineries for Biofuel Production for Sustainable Future Energy Systems. In Recent Trends in Mycological Research: Volume 2: Environmental and Industrial Perspective; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Rosso, M.N.; Berrin, J.G.; Lomascolo, A. Plant Wastes and Sustainable Refineries: What Can We Learn from Fungi? Curr. Opin. Green Sustain. Chem. 2022, 34, 100602. [Google Scholar] [CrossRef]
- Hammel, K.E. Fungal Degradation of Lignin. Driven Nat. Plant Litter Qual. Decompos. 1997, 33, 45. [Google Scholar]
- Sánchez, C. Lignocellulosic Residues: Biodegradation and Bioconversion by Fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef]
- Xu, J.-H. Carbohydrate Active Enzyme Database. In Catalysis from A to Z; Wiley Online Library: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Sharma, R.K.; Arora, D.S. Fungal Degradation of Lignocellulosic Residues: An Aspect of Improved Nutritive Quality. Crit. Rev. Microbiol. 2015, 41, 52–60. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and Classic Families of Secreted Fungal Heme Peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef]
- Vasina, D.V.; Moiseenko, K.V.; Fedorova, T.V.; Tyazhelova, T.V. Lignin-Degrading Peroxidases in White-Rot Fungus Trametes Hirsuta 072. Absolute Expression Quantification of Full Multigene Family. PLoS ONE 2017, 12, e0173813. [Google Scholar] [CrossRef]
- Xie, C.; Gong, W.; Zhu, Z.; Zhou, Y.; Xu, C.; Yan, L.; Hu, Z.; Ai, L.; Peng, Y. Comparative Secretome of White-Rot Fungi Reveals Co-Regulated Carbohydrate-Active Enzymes Associated with Selective Ligninolysis of Ramie Stalks. Microb. Biotechnol. 2021, 14, 911–922. [Google Scholar] [CrossRef]
- Rudakiya, D.M.; Gupte, A. Assessment of White Rot Fungus Mediated Hardwood Degradation by FTIR Spectroscopy and Multivariate Analysis. J. Microbiol. Methods 2019, 157, 123–130. [Google Scholar] [CrossRef]
- Marinović, M.; Aguilar-Pontes, M.V.; Zhou, M.; Miettinen, O.; de Vries, R.P.; Mäkelä, M.R.; Hildén, K. Temporal Transcriptome Analysis of the White-Rot Fungus Obba Rivulosa Shows Expression of a Constitutive Set of Plant Cell Wall Degradation Targeted Genes during Growth on Solid Spruce Wood. Fungal Genet. Biol. 2018, 112, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Marinović, M.; Nousiainen, P.; Dilokpimol, A.; Kontro, J.; Moore, R.; Sipilä, J.; De Vries, R.P.; Mäkelä, M.R.; Hildén, K. Selective Cleavage of Lignin β- O-4 Aryl Ether Bond by β-Etherase of the White-Rot Fungus Dichomitus Squalens. ACS Sustain. Chem. Eng. 2018, 6, 2878–2882. [Google Scholar] [CrossRef] [PubMed]
- Den, W.; Sharma, V.K.; Lee, M.; Nadadur, G.; Varma, R.S. Lignocellulosic Biomass Transformations via Greener Oxidative Pretreatment Processes: Access to Energy and Value Added Chemicals. Front. Chem. 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, B.; Su, X. Linking Enzymatic Oxidative Degradation of Lignin to Organics Detoxification. Int. J. Mol. Sci. 2018, 19, 3373. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, A.; Hammel, K.E. Fungal Biodegradation of Lignocelluloses. In Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Machado, A.S.; Valadares, F.; Silva, T.F.; Milagres, A.M.F.; Segato, F.; Ferraz, A. The Secretome of Phanerochaete Chrysosporium and Trametes Versicolor Grown in Microcrystalline Cellulose and Use of the Enzymes for Hydrolysis of Lignocellulosic Materials. Front. Bioeng. Biotechnol. 2020, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Wahab, A.; Bilal, M.; Iqbal, H.M.N. Delignification of Lignocellulose Biomasses by Alginate–Chitosan Immobilized Laccase Produced from Trametes Versicolor IBL-04. Waste Biomass Valorization 2018, 9, 2071–2079. [Google Scholar] [CrossRef]
- Su, Y.; Xian, H.; Shi, S.; Zhang, C.; Manik, S.M.N.; Mao, J.; Zhang, G.; Liao, W.; Wang, Q.; Liu, H. Biodegradation of Lignin and Nicotine with White Rot Fungi for the Delignification and Detoxification of Tobacco Stalk. BMC Biotechnol. 2016, 16, 81. [Google Scholar] [CrossRef] [PubMed]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; Cone, J.W. Fungal Treatment of Lignocellulosic Biomass: Importance of Fungal Species, Colonization and Time on Chemical Composition and in Vitro Rumen Degradability. Anim. Feed Sci. Technol. 2015, 209, 40–50. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, J.; Ma, F.; Tang, C.; Tang, Q.; Zhang, X. Investigation of Lignocellulolytic Enzymes during Different Growth Phases of Ganoderma Lucidum Strain G0119 Using Genomic, Transcriptomic and Secretomic Analyses. PLoS ONE 2018, 13, e0198404. [Google Scholar] [CrossRef]
- Krcmar, P.; Novotny, C.; Marais, M.F.; Joseleau, J.P. Structure of Extracellular Polysaccharide Produced by Lignin-Degrading Fungus Phlebia Radiata in Liquid Culture. Int. J. Biol. Macromol. 1999, 24, 61–64. [Google Scholar] [CrossRef]
- Fonseca, M.I.; Molina, M.A.; Winnik, D.L.; Busi, M.V.; Fariña, J.I.; Villalba, L.L.; Zapata, P.D. Isolation of a Laccase-Coding Gene from the Lignin-Degrading Fungus Phlebia Brevispora BAFC 633 and Heterologous Expression in Pichia Pastoris. J. Appl. Microbiol. 2018, 124, 1454–1468. [Google Scholar] [CrossRef]
- Hirakawa, M.P.; Rodriguez, A.; Tran-Gyamfi, M.B.; Light, Y.K.; Martinez, S.; Diamond-Pott, H.; Simmons, B.A.; Sale, K.L. Phenothiazines Rapidly Induce Laccase Expression and Lignin-Degrading Properties in the White-Rot Fungus Phlebia Radiata. J. Fungi 2023, 9, 371. [Google Scholar] [CrossRef]
- Xu, X.; Xu, Z.; Shi, S.; Lin, M. Lignocellulose Degradation Patterns, Structural Changes, and Enzyme Secretion by Inonotus Obliquus on Straw Biomass under Submerged Fermentation. Bioresour. Technol. 2017, 241, 415–423. [Google Scholar] [CrossRef]
- Floudas, D.; Bentzer, J.; Ahrén, D.; Johansson, T.; Persson, P.; Tunlid, A. Uncovering the Hidden Diversity of Litter-Decomposition Mechanisms in Mushroom-Forming Fungi. ISME J. 2020, 14, 2046–2059. [Google Scholar] [CrossRef]
- Qi, J.; Zhang, X.; Zhou, Y.; Zhang, C.; Wen, J.; Deng, S.; Luo, B.; Fan, M.; Xia, Y. Selectively Enzymatic Conversion of Wood Constituents with White and Brown Rot Fungi. Ind. Crops Prod. 2023, 199, 116703. [Google Scholar] [CrossRef]
- Goodell, B.; Zhu, Y.; Kim, S.; Kafle, K.; Eastwood, D.; Daniel, G.; Jellison, J.; Yoshida, M.; Groom, L.; Pingali, S.V.; et al. Modification of the Nanostructure of Lignocellulose Cell Walls via a Non-Enzymatic Lignocellulose Deconstruction System in Brown Rot Wood-Decay Fungi. Biotechnol. Biofuels 2017, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, E.; Schalchli, H.; Diez, M.C. Biodegradation Inducers to Enhance Wheat Straw Pretreatment by Gloeophyllum Trabeum to Second-Generation Ethanol Production. Environ. Sci. Pollut. Res. 2020, 27, 8467–8480. [Google Scholar] [CrossRef]
- Kaffenberger, J.T.; Schilling, J.S. Comparing Lignocellulose Physiochemistry after Decomposition by Brown Rot Fungi with Distinct Evolutionary Origins. Environ. Microbiol. 2015, 17, 4885–4897. [Google Scholar] [CrossRef] [PubMed]
- Obeng, E.M.; Adam, S.N.N.; Budiman, C.; Ongkudon, C.M.; Maas, R.; Jose, J. Lignocellulases: A Review of Emerging and Developing Enzymes, Systems, and Practices. Bioresour. Bioprocess. 2017, 4, 16. [Google Scholar] [CrossRef]
- Mirete, S.; Morgante, V.; González-Pastor, J.E. Acidophiles: Diversity and Mechanisms of Adaptation to Acidic Environments. In Adaption of Microbial Life to Environmental Extremes: Novel Research Results and Application, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Dennis, P.G.; Newsham, K.K.; Rushton, S.P.; O’Donnell, A.G.; Hopkins, D.W. Soil Bacterial Diversity Is Positively Associated with Air Temperature in the Maritime Antarctic. Sci. Rep. 2019, 9, 2686. [Google Scholar] [CrossRef] [PubMed]
- Walkup, J.; Dang, C.; Mau, R.L.; Hayer, M.; Schwartz, E.; Stone, B.W.; Hofmockel, K.S.; Koch, B.J.; Purcell, A.M.; Pett-Ridge, J.; et al. The Predictive Power of Phylogeny on Growth Rates in Soil Bacterial Communities. ISME Commun. 2023, 3, 73. [Google Scholar] [CrossRef]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T.A. Alkaline PH Homeostasis in Bacteria: New Insights. Biochim. Biophys. Acta—Biomembr. 2005, 1717, 67–88. [Google Scholar] [CrossRef]
- Harindintwali, J.D.; Zhou, J.; Yu, X. Lignocellulosic Crop Residue Composting by Cellulolytic Nitrogen-Fixing Bacteria: A Novel Tool for Environmental Sustainability. Sci. Total Environ. 2020, 715, 136912. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Algora, C.; Baldrian, P. Lignocellulolytic Systems of Soil Bacteria: A Vast and Diverse Toolbox for Biotechnological Conversion Processes. Biotechnol. Adv. 2019, 37, 107374. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.M.; Allgaier, M.; Chavarria, Y.; Fortney, J.L.; Hugenholtz, P.; Simmons, B.; Sublette, K.; Silver, W.L.; Hazen, T.C. Characterization of Trapped Lignin-Degrading Microbes in Tropical Forest Soil. PLoS ONE 2011, 6, e19306. [Google Scholar] [CrossRef] [PubMed]
- Tláskal, V.; Zrůstová, P.; Vrška, T.; Baldrian, P. Bacteria Associated with Decomposing Dead Wood in a Natural Temperate Forest. FEMS Microbiol. Ecol. 2017, 93, fix157. [Google Scholar] [CrossRef]
- Arcand, M.M.; Helgason, B.L.; Lemke, R.L. Microbial Crop Residue Decomposition Dynamics in Organic and Conventionally Managed Soils. Appl. Soil Ecol. 2016, 107, 347–359. [Google Scholar] [CrossRef]
- Georgiadou, D.N.; Avramidis, P.; Ioannou, E.; Hatzinikolaou, D.G. Microbial Bioprospecting for Lignocellulose Degradation at a Unique Greek Environment. Heliyon 2021, 7, e07122. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Liu, W.; Ren, S.; Wang, D. Effect of nano-magnetite and ethanol on methane production in the anaerobic digestion of wheat straw. J. ASABE 2022, 65, 707–714. [Google Scholar] [CrossRef]
- Singhvi, M.S.; Gokhale, D.V. Lignocellulosic Biomass: Hurdles and Challenges in Its Valorization. Appl. Microbiol. Biotechnol. 2019, 103, 9305–9320. [Google Scholar] [CrossRef]
- Hassan, S.S.; Williams, G.A.; Jaiswal, A.K. Moving towards the Second Generation of Lignocellulosic Biorefineries in the EU: Drivers, Challenges, and Opportunities. Renew. Sustain. Energy Rev. 2019, 101, 590–599. [Google Scholar] [CrossRef]
- Beaton, D.; Pelletier, P.; Goulet, R.R. Microbial Degradation of Cellulosic Material and Gas Generation: Implications for the Management of Low-and Intermediate-Level Radioactive Waste. Front. Microbiol. 2019, 10, 204. [Google Scholar] [CrossRef]
- Piccinni, F.; Murua, Y.; Ghio, S.; Talia, P.; Rivarola, M.; Campos, E. Draft Genome Sequence of Cellulolytic and Xylanolytic Cellulomonas sp. Strain B6 Isolated from Subtropical Forest Soil. Genome Announc. 2016, 4, e00891-16. [Google Scholar] [CrossRef]
- Wang, W.; Yan, L.; Cui, Z.; Gao, Y.; Wang, Y.; Jing, R. Characterization of a Microbial Consortium Capable of Degrading Lignocellulose. Bioresour. Technol. 2011, 102, 9321–9324. [Google Scholar] [CrossRef] [PubMed]
- De Lima Brossi, M.J.; Jiménez, D.J.; Cortes-Tolalpa, L.; van Elsas, J.D. Soil-Derived Microbial Consortia Enriched with Different Plant Biomass Reveal Distinct Players Acting in Lignocellulose Degradation. Microb. Ecol. 2016, 71, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, G.L.; Rodriguez, J.E.; Domont, G.B.; de Souza, W.; Junqueira, M.; Frases, S. Biochemical Characterization of Streptomyces Sp. I1.2 Secretome Reveals the Presence of Multienzymatic Complexes Containing Cellulases and Accessory Enzymes. Bioenergy Res. 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Olk, D.C.; Samson, M.I.; Gapas, P. Inhibition of Nitrogen Mineralization in Young Humic Fractions by Anaerobic Decomposition of Rice Crop Residues. Eur. J. Soil Sci. 2007, 58, 270–281. [Google Scholar] [CrossRef]
- Gao, H.; Chen, X.; Wei, J.; Zhang, Y.; Zhang, L.; Chang, J.; Thompson, M.L. Decomposition Dynamics and Changes in Chemical Composition of Wheat Straw Residue under Anaerobic and Aerobic Conditions. PLoS ONE 2016, 11, e0158172. [Google Scholar] [CrossRef] [PubMed]
- Bierke, A.; Kaiser, K.; Guggenberger, G. Crop Residue Management Effects on Organic Matter in Paddy Soils—The Lignin Component. Geoderma 2008, 146, 48–57. [Google Scholar] [CrossRef]
- Olk, D.C.; Cassman, K.G.; Schmidt-Rohr, K.; Anders, M.M.; Mao, J.D.; Deenik, J.L. Chemical Stabilization of Soil Organic Nitrogen by Phenolic Lignin Residues in Anaerobic Agroecosystems. Soil Biol. Biochem. 2006, 38, 3303–3312. [Google Scholar] [CrossRef]
- Doran, J.W. Microbial Biomass and Mineralizable Nitrogen Distributions in No-Tillage and Plowed Soils. Biol. Fertil. Soils 1987, 5, 68–75. [Google Scholar] [CrossRef]
- Olk, D.C.; Cassman, K.G.; Mahieu, N.; Randall, E.W. Conserved Chemical Properties of Young Humic Acid Fractions in Tropical Lowland Soil under Intensive Irrigated Rice Cropping. Eur. J. Soil Sci. 1998, 49, 337–349. [Google Scholar] [CrossRef]
- Ljungdahl, L.G.; Eriksson, K.-E. Ecology of Microbial Cellulose Degradation; Springer: Boston, MA, USA, 1985. [Google Scholar] [CrossRef]
- Kinet, R.; Destain, J.; Hiligsmann, S.; Thonart, P.; Delhalle, L.; Taminiau, B.; Daube, G.; Delvigne, F. Thermophilic and Cellulolytic Consortium Isolated from Composting Plants Improves Anaerobic Digestion of Cellulosic Biomass: Toward a Microbial Resource Management Approach. Bioresour. Technol. 2015, 189, 138–144. [Google Scholar] [CrossRef]
- Hamann, P.R.V.; Silva, L.D.M.B.; Gomes, T.C.; Noronha, E.F. Assembling Mini-Xylanosomes with Clostridium Thermocellum XynA, and Their Properties in Lignocellulose Deconstruction. Enzyme Microb. Technol. 2021, 150, 109887. [Google Scholar] [CrossRef] [PubMed]
- Eibinger, M.; Ganner, T.; Plank, H.; Nidetzky, B. A Biological Nanomachine at Work: Watching the Cellulosome Degrade Crystalline Cellulose. ACS Cent. Sci. 2020, 6, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Rainey, F.A.; Ward-Rainey, N.L. Proposal for a New Hierarchic Classification System, Actinobacteria Classis Nov. Int. J. Syst. Bacteriol. 1997, 47, 479–491. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Větrovský, T.; Steffen, K.T.; Baldrian, P. Potential of Cometabolic Transformation of Polysaccharides and Lignin in Lignocellulose by Soil Actinobacteria. PLoS ONE 2014, 9, e89108. [Google Scholar] [CrossRef]
- Bastian, F.; Bouziri, L.; Nicolardot, B.; Ranjard, L. Impact of Wheat Straw Decomposition on Successional Patterns of Soil Microbial Community Structure. Soil Biol. Biochem. 2009, 41, 262–275. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, S.; Niu, Q.; Yan, H.; Li, Q. The Influences of Illite/Smectite Clay on Lignocellulose Decomposition and Humification Process Revealed by Metagenomics Analysis during Cattle Manure Composting. Waste Manag. 2021, 127, 1–9. [Google Scholar] [CrossRef]
- Wang, C.; Dong, D.; Wang, H.; Müller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic Analysis of Microbial Consortia Enriched from Compost: New Insights into the Role of Actinobacteria in Lignocellulose Decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef]
- Wang, W.; Yu, Y.; Dou, T.Y.; Wang, J.Y.; Sun, C. Species of Family Promicromonosporaceae and Family Cellulomonadeceae That Produce Cellulosome-like Multiprotein Complexes. Biotechnol. Lett. 2018, 40, 335–341. [Google Scholar] [CrossRef]
- Lewin, G.R.; Carlos, C.; Chevrette, M.G.; Horn, H.A.; McDonald, B.R.; Stankey, R.J.; Fox, B.G.; Currie, C.R. Evolution and Ecology of Actinobacteria and Their Bioenergy Applications. Annu. Rev. Microbiol. 2016, 70, 235–254. [Google Scholar] [CrossRef]
- Seneviratne, G. Litter Quality and Nitrogen Release in Tropical Agriculture: A Synthesis. Biol. Fertil. Soils 2000, 31, 60–64. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Senthilkumar, M.; Ramakrishnan, B. Endophytic Actinobacteria: Nitrogen Fixation, Phytohormone Production, and Antibiosis. In Plant Growth Promoting Actinobacteria: A New Avenue for Enhancing the Productivity and Soil Fertility of Grain Legumes; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Mitra, D.; Mondal, R.; Khoshru, B.; Senapati, A.; Radha, T.K.; Mahakur, B.; Uniyal, N.; Myo, E.M.; Boutaj, H.; Sierra, B.E.G.; et al. Actinobacteria-Enhanced Plant Growth, Nutrient Acquisition, and Crop Protection: Advances in Soil, Plant, and Microbial Multifactorial Interactions. Pedosphere 2022, 32, 149–170. [Google Scholar] [CrossRef]
- Hamedi, J.; Mohammadipanah, F. Biotechnological Application and Taxonomical Distribution of Plant Growth Promoting Actinobacteria. J. Ind. Microbiol. Biotechnol. 2015, 42, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, G.L.; de Azevedo-Martins, A.C.; Albano, R.M.; de Souza, W.; Frases, S. Comprehensive Analysis of the Cellulolytic System Reveals Its Potential for Deconstruction of Lignocellulosic Biomass in a Novel Streptomyces Sp. Appl. Microbiol. Biotechnol. 2017, 101, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, H.M.; El-Shatoury, S.A. Actinomycetes in Rice Straw Decomposition. Waste Manag. 2007, 27, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Lam, M.Q.; Thevarajoo, S.; Abd Manan, F.; Yahya, A.; Chong, C.S. Genome Analysis of Cellulose and Hemicellulose Degrading Micromonospora Sp. CP22. 3 Biotech 2020, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, M.; Ball, A. Degradation of Lignocellulose by Extracellular Enzymes Produced by Thermomonospora Fusca BD25. Appl. Microbiol. Biotechnol. 2002, 58, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.Y.; Huang, Y.C.; Chen, W.L.; Chen, C.Y.; Yang, C.H. Properties of Thermobifida Fusca Peroxidase Tfu-1649 and Its Combined Synergistic Effects with Xylanase on Lignocellulose Degradation. Bioresources 2021, 16, 942–953. [Google Scholar] [CrossRef]
- Yin, Y.R.; Sang, P.; Xian, W.D.; Li, X.; Jiao, J.Y.; Liu, L.; Hozzein, W.N.; Xiao, M.; Li, W.J. Expression and Characteristics of Two Glucose-Tolerant GH1 β-Glucosidases From Actinomadura Amylolytica YIM 77502T for Promoting Cellulose Degradation. Front. Microbiol. 2018, 9, 3149. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; He, C.; Wang, Z.; Luo, X.; Sun, X.; Zhao, J.; Gao, X.; Xiang, W.; Song, J.; Wang, X. Nocardia Rosealba Sp. Nov., a Novel Ligninase-Producing Actinobacterium Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2022, 72, 005416. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Singh, D.; Laskar, D.D.; Chen, S. Degradation of Native Wheat Straw Lignin by Streptomyces Viridosporus T7A. Int. J. Environ. Sci. Technol. 2013, 10, 165–174. [Google Scholar] [CrossRef]
- Gong, X.; Yu, Y.; Hao, Y.; Wang, Q.; Ma, J.; Jiang, Y.; Lv, G.; Li, L.; Qian, C. Characterizing Corn-Straw-Degrading Actinomycetes and Evaluating Application Efficiency in Straw-Returning Experiments. Front. Microbiol. 2022, 13, 1003157. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.W.; Reddy, A.P.; D’haeseleer, P.; Khudyakov, J.; Billis, K.; Pati, A.; Simmons, B.A.; Singer, S.W.; Thelen, M.P.; Vandergheynst, J.S. Metatranscriptomic Analysis of Lignocellulolytic Microbial Communities Involved in High-Solids Decomposition of Rice Straw. Biotechnol. Biofuels 2014, 7, 495. [Google Scholar] [CrossRef]
- Xia, Y.; Lin, X. Efficient Biodegradation of Straw and Persistent Organic Pollutants by a Novel Strategy Using Recombinant Trichoderma Reesei. Bioresour. Bioprocess. 2022, 9, 91. [Google Scholar] [CrossRef]
- Organo, N.D.; Granada, S.M.J.M.; Pineda, H.G.S.; Sandro, J.M.; Nguyen, V.H.; Gummert, M. Assessing the Potential of a Trichoderma-Based Compost Activator to Hasten the Decomposition of Incorporated Rice Straw. Sci. Rep. 2022, 12, 448. [Google Scholar] [CrossRef]
- Choudhary, M.; Sharma, P.C.; Garg, N. Crop Residue Degradation by Autochthonous Fungi Isolated from Cropping System Management Scenarios. Bioresources 2015, 10, 5809–5819. [Google Scholar] [CrossRef]
- Wendt, L.M.; Wahlen, B.D.; Walton, M.R.; Nguyen, J.A.; Lin, Y.; Brown, R.M.; Zhao, H. Exploring Filamentous Fungi Depolymerization of Corn Stover in the Context Bioenergy Queuing Operations. Food Energy Secur. 2022, 11, e333. [Google Scholar] [CrossRef]
- Hermosilla, E.; Rubilar, O.; Schalchli, H.; da Silva, A.S.A.; Ferreira-Leitao, V.; Diez, M.C. Sequential White-Rot and Brown-Rot Fungal Pretreatment of Wheat Straw as a Promising Alternative for Complementary Mild Treatments. Waste Manag. 2018, 79, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Che, S.; Qin, X.; Xu, Y.; Tian, S.; Zhu, Y.; Song, J.; Guan, Y.; Wang, D.; Wu, M.; et al. Identification, Characteristics and Rice Growth Promotion of a Highly Efficient Cellulolytic Bacterial Strain, Cellulomonas Iranensis ZJW-6, Isolated from Paddy Soil in Central China. Front. Microbiol. 2023, 14, 1152966. [Google Scholar] [CrossRef] [PubMed]
- Vu, V.; Farkas, C.; Riyad, O.; Bujna, E.; Kilin, A.; Sipiczki, G.; Sharma, M.; Usmani, Z.; Gupta, V.K.; Nguyen, Q.D. Enhancement of the Enzymatic Hydrolysis Efficiency of Wheat Bran Using the Bacillus Strains and Their Consortium. Bioresour. Technol. 2022, 343, 126092. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Khatiwada, J.R.; Kognou, A.L.M.; Chio, C.; Qin, W. Biomass-Degrading Enzyme(s) Production and Biomass Degradation by a Novel Streptomyces Thermocarboxydus. Curr. Microbiol. 2023, 80, 71. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Dong, C.; Wang, S.; Danso, B.; Dar, M.A.; Pandit, R.S.; Pawar, K.D.; Geng, A.; Zhu, D.; Li, X.; et al. Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion. Insects 2023, 14, 403. [Google Scholar] [CrossRef]
- Guan, Y.; Zhu, H.; Zhu, Y.; Zhao, H.; Shu, L.; Song, J.; Yang, X.; Wu, Z.; Wu, L.; Yang, M. Microbial Consortium Composed of Cellulomonas ZJW-6 and Acinetobacter DA-25 Improves Straw Lignocellulose Degradation. Arch. Microbiol. 2022, 204, 139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Elzenga, T.; van Elsas, J.D. Effect of Culture Conditions on the Performance of Lignocellulose-Degrading Synthetic Microbial Consortia. Appl. Microbiol. Biotechnol. 2021, 105, 7981–7995. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. World Crop Residues Production and Implications of Its Use as a Biofuel. Environ. Int. 2005, 31, 575–584. [Google Scholar] [CrossRef]
- Toronto, C.E.; Remington, R. A Step-by-Step Guide to Conducting an Integrative Review; Toronto, C.E., Remington, R., Eds.; Springer International Publishing: Cham, Switzerland, 2020; Volume 1. [Google Scholar] [CrossRef]
- Priem, J.; Piwowar, H.; Orr, R. OpenAlex: A fully-open index of scholarly works, authors, venues, institutions, and concepts. arXiv 2022, arXiv:2205.01833. [Google Scholar]
- Van Eck, N.J.; Waltman, L. Software Survey: VOSviewer, a Computer Program for Bibliometric Mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cereal Crops | Area (103 Kha) | Production (103 Kt) | Residue Production (103 Kt) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2001 | 2011 | 2021 | 2001 | 2011 | 2021 | 2001 | 2011 | 2021 | |
| Barley | 3892.5 | 3123.3 | 3100 | 17,213 | 14,865.3 | 15,764.350 | 25,820.1 | 22,298 | 23,646.5 |
| Maize | 4.7 | 20.2 | 26 | 10.9 | 127.2 | 164.850 | 10.9 | 127.2 | 164.9 |
| Oats | 1126.2 | 881.8 | 1065.6 | 3859.3 | 3284.9 | 3739.330 | 3859.2 | 3284.9 | 3739.3 |
| Rye | 324.5 | 203.5 | 279.6 | 985 | 733 | 1361.719 | 1477.5 | 1099.4 | 2042.6 |
| Wheat | 3539.9 | 4543.2 | 4806.6 | 21,793.7 | 27,699 | 30,037.550 | 32,690.6 | 41,548.5 | 45,056.3 |
| Total | 8887.9 | 8772 | 9277.9 | 43,862.3 | 46,709.3 | 51,067.799 | 63,858.4 | 68,357.9 | 74,649.6 |
| Cereal Crops | Area (103 Kha) | Production (103 Kt) | Residue Production (103 Kt) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2001 | 2011 | 2021 | 2001 | 2011 | 2021 | 2001 | 2011 | 2021 | |
| Barley | 336 | 252.7 | 144.7 | 776.2 | 759.8 | 500.4 | 1164.3 | 1139.7 | 750.6 |
| Maize | 4.7 | 9.6 | 17.9 | 10.9 | 71.9 | 104.7 | 10.9 | 71.9 | 104.7 |
| Oats | 47.9 | 63.2 | 92.4 | 84.3 | 128.5 | 170.3 | 84.3 | 128.5 | 170.3 |
| Rye | 111.3 | 42 | 26.1 | 231.1 | 85 | 63.5 | 346.7 | 127.5 | 95.2 |
| Wheat | 352.2 | 551.1 | 944.2 | 1076.3 | 1869.3 | 4248.9 | 1614.5 | 2804 | 6373.3 |
| Total | 851.6 | 918.6 | 1225.3 | 2178.8 | 2914.5 | 5087.7 | 3220.6 | 4271.6 | 7494 |
| Crop Residue | Cellulose (%) | Hemicellulose (%) | Lignin (%) | Reference |
|---|---|---|---|---|
| Wheat straw | 34–39 | 26–32 | 16–19 | [35,36] |
| Oat straw | 27–34 | 24–33 | 9–15 | [37,38,39] |
| Rye straw | 26–36 | 28–34 | 13–21 | [40,41] |
| Maize stalks | 38–43 | 26–31 | 4–10 | [36,42] |
| Barley straw | 35–44 | 27–36 | 14–18 | [41,43] |
| Microorganism | Residue Type | Method | Days | Mass Loss, % | Enzyme(s) Evaluated | Reference |
|---|---|---|---|---|---|---|
| Trichoderma reesei | Rice straw, bran | Solid-state fermentation | 10 | 51.16 | Laccase, xylanase, β-Glucosidase, cellobiohydrolase, endoglucanase | [172] |
| Trichoderma harzianum | Rice straw | In situ | 28 | 23.69 | - | [173] |
| Aspergillus niger | Rice and wheat straw (4:1) | Solid-state fermentation | 10 | 16 | CMCase, endoglucanase, cellobiase, β-1,4-xylanase | [174] |
| Phanerochaete chrysosporium | Maize stover | Solid-state fermentation | 28 | 21 | - | [175] |
| Ganoderma lobatum | Wheat straw | Solid-state fermentation | 40 | 21.04 | β-glucosidase | [176] |
| Cellulomonas sp. | Rice straw | Submerged fermentation | 4 | 49.3 | β-glucosidase, endoglucanase, exoglucanase, xylanase, lignin peroxidase, manganese peroxidase, laccase. | [177] |
| Bacillus sp. | Wheat bran | Submerged fermentation | 7 | 60 | Cellulase, endoglucanase, xylanase, laccase, mannase | [178] |
| Streptomyces sp. | Barley straw | Submerged fermentation | 7 | 60.55 | Exoglucanase, endoglucanase, β-glucosidase | [179] |
| Enterobacter sp. | Rice straw | Submerged fermentation | 7 | 45.52 | Endoglucanases, exoglucanase, xylanase | [180] |
| Ganoderma lobatum + Gloeophyllum trabeum | Wheat straw | Solid-state fermentation | 20 | 15.52 | β-glucosidase | [176] |
| Cellulomonas ZJW-6 + Acinetobacter DA-25 | Rice straw | Submerged fermentation | 4 | 57.62 | β-glucosidase, endoglucanase, xylanase, lignin peroxidase, laccase, manganese peroxidase, β-glucosidase | [181] |
| Streptomyces sp. G1T + Streptomyces sp. G2T + Streptomyces sp. G3T | Maize stalk | solid-state fermentation | 119 | 66.37 | - | [170] |
| Citrobacter freundii so4 + Sphingobacterium multivorum w15 + Coniochaeta sp. 2T2.1 | Wheat straw | Submerged fermentation | 10 | 12.82 | - | [182] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamshitov, A.; Kadžienė, G.; Supronienė, S. The Role of Soil Microbial Consortia in Sustainable Cereal Crop Residue Management. Plants 2024, 13, 766. https://doi.org/10.3390/plants13060766
Shamshitov A, Kadžienė G, Supronienė S. The Role of Soil Microbial Consortia in Sustainable Cereal Crop Residue Management. Plants. 2024; 13(6):766. https://doi.org/10.3390/plants13060766
Chicago/Turabian StyleShamshitov, Arman, Gražina Kadžienė, and Skaidrė Supronienė. 2024. "The Role of Soil Microbial Consortia in Sustainable Cereal Crop Residue Management" Plants 13, no. 6: 766. https://doi.org/10.3390/plants13060766