

Distributing Plant Developmental Regulatory Proteins via Plasmodesmata


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Setting the Stage for Intercellular Transport: Biogenesis and Regulation of Plasmodesmata
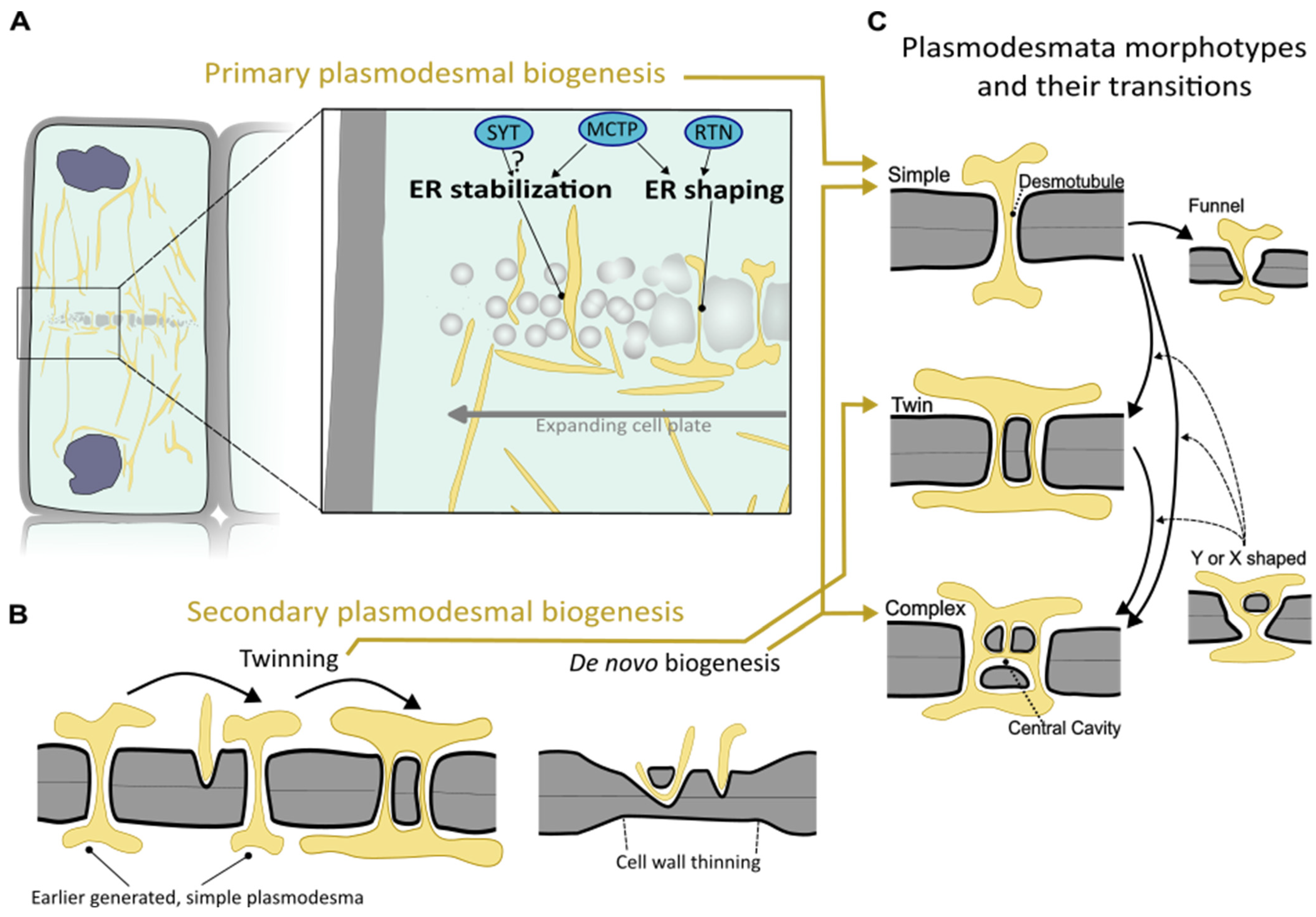
2.1. Formation and Control over Primary Plasmodesmata
2.2. Secondary Plasmodesmata and Diversification of Plasmodesmal Shape
2.3. Dynamic Regulation of Plasmodesmata
2.3.1. Callose Turnover as Tuning Mechanism for Plasmodesmal Function
2.3.2. Turgor Pressure and Plasmodesmal Conductivity
2.3.3. Control by Actin and Associated Proteins
3. Molecular Mechanisms Regulating Transport of Proteins Carrying Developmental Instructions
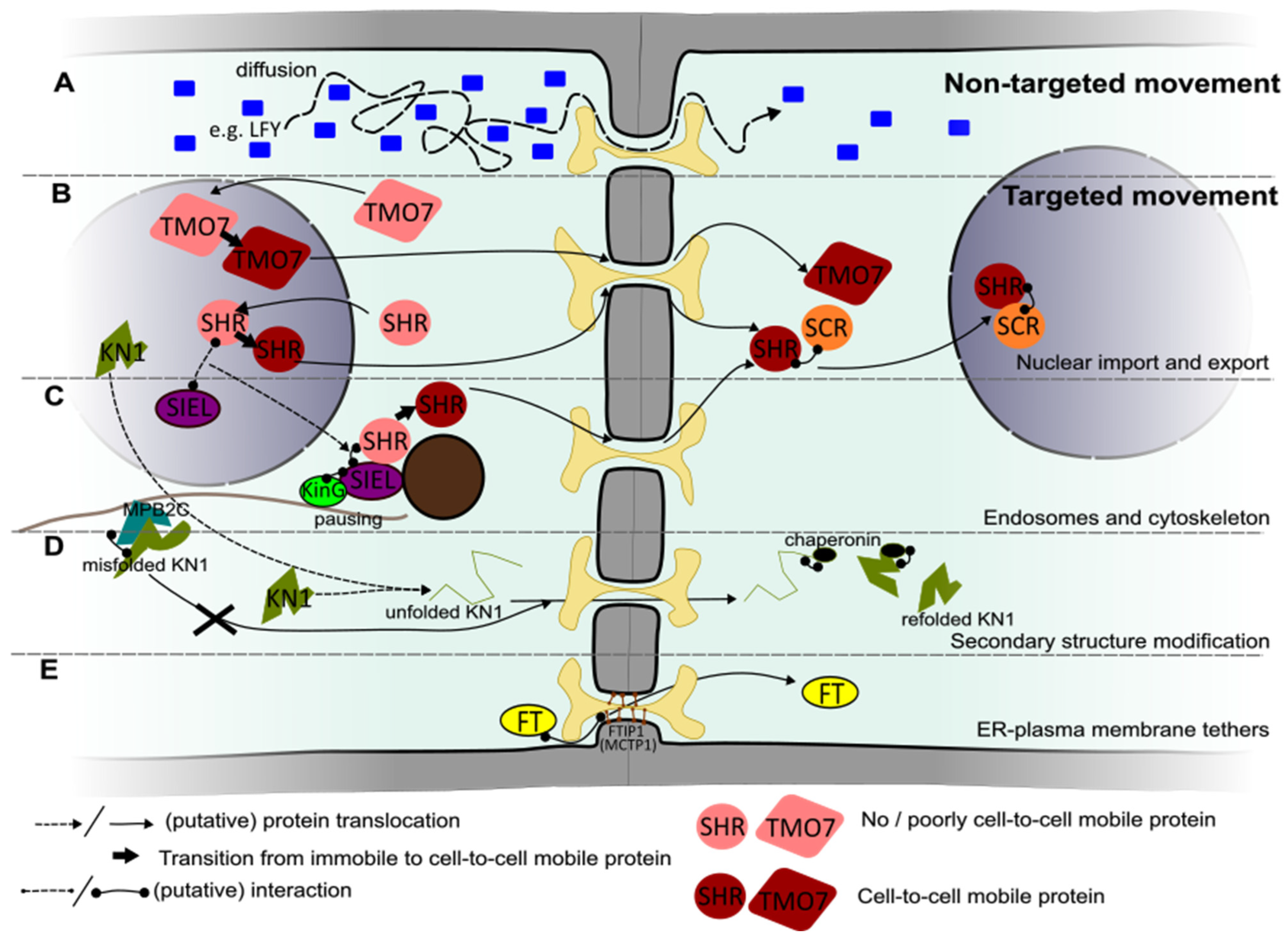
3.1. Non-Targeted Transport
3.2. Targeted Transport
3.2.1. Nuclear Localization Required for Intercellular Protein Transport
3.2.2. Role of Endosomes and the Cytoskeleton in Intercellular Transport
3.3. How Does a Protein Traverse through Plasmodesmata?
4. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaigne, A.; Brunet, T. Incomplete Abscission and Cytoplasmic Bridges in the Evolution of Eukaryotic Multicellularity. Curr. Biol. 2022, 32, R385–R397. [Google Scholar] [CrossRef] [PubMed]
- Wegner, L.; Porth, M.L.; Ehlers, K. Multicellularity and the Need for Communication—A Systematic Overview on (Algal) Plasmodesmata and Other Types of Symplasmic Cell Connections. Plants 2023, 12, 3342. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Zambryski, P.C. Plasmodesmata Paradigm Shift: Regulation from without versus within. Annu. Rev. Plant Biol. 2012, 63, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Sager, R.; Lee, J.-Y. Plasmodesmata in Integrated Cell Signalling: Insights from Development and Environmental Signals and Stresses. J. Exp. Bot. 2014, 65, 6337–6358. [Google Scholar] [CrossRef]
- Cheval, C.; Faulkner, C. Plasmodesmal Regulation during Plant–Pathogen Interactions. New Phytol. 2018, 217, 62–67. [Google Scholar] [CrossRef]
- Wang, Y.; Perez-Sancho, J.; Platre, M.P.; Callebaut, B.; Smokvarska, M.; Ferrer, K.; Luo, Y.; Nolan, T.M.; Sato, T.; Busch, W.; et al. Plasmodesmata Mediate Cell-to-Cell Transport of Brassinosteroid Hormones. Nat. Chem. Biol. 2023, 19, 1331–1341. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Gao, Y.; Zhang, Z.; Gao, C.; Chen, S.; Liesche, J. Parallel Auxin Transport via PINs and Plasmodesmata during the Arabidopsis Leaf Hyponasty Response. Plant Cell Rep. 2023, 43, 4. [Google Scholar] [CrossRef]
- Linh, N.M.; Scarpella, E. Leaf Vein Patterning Is Regulated by the Aperture of Plasmodesmata Intercellular Channels. PLOS Biol. 2022, 20, e3001781. [Google Scholar] [CrossRef]
- Band, L.R. Auxin Fluxes through Plasmodesmata. New Phytol. 2021, 231, 1686–1692. [Google Scholar] [CrossRef]
- Paterlini, A. Uncharted Routes: Exploring the Relevance of Auxin Movement via Plasmodesmata. Biol. Open 2020, 9, bio055541. [Google Scholar] [CrossRef] [PubMed]
- Maizel, A.; Markmann, K.; Timmermans, M.; Wachter, A. To Move or Not to Move: Roles and Specificity of Plant RNA Mobility. Curr. Opin. Plant Biol. 2020, 57, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Furuta, K.; Lichtenberger, R.; Helariutta, Y. The Role of Mobile Small RNA Species during Root Growth and Development. Curr. Opin. Cell Biol. 2012, 24, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Sparks, E.; Wachsman, G.; Benfey, P.N. Spatiotemporal Signalling in Plant Development. Nat. Rev. Genet. 2013, 14, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Rim, Y.; Wang, J.; Jackson, D. A Novel Cell-to-Cell Trafficking Assay Indicates That the KNOX Homeodomain Is Necessary and Sufficient for Intercellular Protein and mRNA Trafficking. Genes Dev. 2005, 19, 788. [Google Scholar] [CrossRef]
- Benitez-Alfonso, Y.; Faulkner, C.; Pendle, A.; Miyashima, S.; Helariutta, Y.; Maule, A. Symplastic Intercellular Connectivity Regulates Lateral Root Patterning. Dev. Cell 2013, 26, 136–147. [Google Scholar] [CrossRef]
- Wu, S.; Gallagher, K.L. Transcription Factors on the Move. Curr. Opin. Plant Biol. 2012, 15, 645–651. [Google Scholar] [CrossRef]
- Gundu, S.; Tabassum, N.; Blilou, I. Moving with Purpose and Direction: Transcription Factor Movement and Cell Fate Determination Revisited. Curr. Opin. Plant Biol. 2020, 57, 124–132. [Google Scholar] [CrossRef]
- Ehlers, K.; Kollmann, R. Primary and Secondary Plasmodesmata: Structure, Origin, and Functioning. Protoplasma 2001, 216, 1–30. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Stonebloom, S.; Xu, M.; Zambryski, P.C. Plasmodesmata during Development: Re-Examination of the Importance of Primary, Secondary, and Branched Plasmodesmata Structure versus Function. Protoplasma 2011, 248, 61–74. [Google Scholar] [CrossRef]
- Sinclair, R.; Hsu, G.; Davis, D.; Chang, M.; Rosquete, M.; Iwasa, J.H.; Drakakaki, G. Plant Cytokinesis and the Construction of New Cell Wall. FEBS Lett. 2022, 596, 2243–2255. [Google Scholar] [CrossRef]
- Porter, K.R.; Machado, R.D. Studies on the Endoplasmic Reticulum: IV. Its Form and Distribution during Mitosis in Cells of Onion Root Tip. J. Biophys. Biochem. Cytol. 1960, 7, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K. Endoplasmic Reticulum in the Formation of the Cell Plate and Plasmodesmata. Protoplasma 1982, 111, 121–133. [Google Scholar] [CrossRef]
- Hawes, C.R.; Juniper, B.E.; Horne, J.C. Low and High Voltage Electron Microscopy of Mitosis and Cytokinesis in Maize Roots. Planta 1981, 152, 397–407. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M.; Austin, J.R.; White, E.A.; Staehelin, L.A. Electron Tomographic Analysis of Somatic Cell Plate Formation in Meristematic Cells of Arabidopsis Preserved by High-Pressure Freezing. Plant Cell 2004, 16, 836–856. [Google Scholar] [CrossRef]
- Nicolas, W.J.; Grison, M.S.; Trépout, S.; Gaston, A.; Fouché, M.; Cordelières, F.P.; Oparka, K.; Tilsner, J.; Brocard, L.; Bayer, E.M. Architecture and Permeability of Post-Cytokinesis Plasmodesmata Lacking Cytoplasmic Sleeves. Nat. Plants 2017, 3, 17082. [Google Scholar] [CrossRef]
- Li, Z.P.; Moreau, H.; Petit, J.D.; Souza-Moraes, T.; Smokvarska, M.; Perez-Sancho, J.; Petrel, M.; Decoeur, F.; Brocard, L.; Chambaud, C.; et al. Plant Plasmodesmata Bridges Form through ER-Driven Incomplete Cytokinesis. bioRxiv 2023. [Google Scholar] [CrossRef]
- Shibata, Y.; Voss, C.; Rist, J.M.; Hu, J.; Rapoport, T.A.; Prinz, W.A.; Voeltz, G.K. The Reticulon and DP1/Yop1p Proteins Form Immobile Oligomers in the Tubular Endoplasmic Reticulum. J. Biol. Chem. 2008, 283, 18892–18904. [Google Scholar] [CrossRef]
- Tolley, N.; Sparkes, I.; Craddock, C.P.; Eastmond, P.J.; Runions, J.; Hawes, C.; Frigerio, L. Transmembrane Domain Length Is Responsible for the Ability of a Plant Reticulon to Shape Endoplasmic Reticulum Tubules in Vivo. Plant J. 2010, 64, 411–418. [Google Scholar] [CrossRef]
- Knox, K.; Wang, P.; Kriechbaumer, V.; Tilsner, J.; Frigerio, L.; Sparkes, I.; Hawes, C.; Oparka, K. Putting the Squeeze on Plasmodesmata: A Role for Reticulons in Primary Plasmodesmata Formation. Plant Physiol. 2015, 168, 1563–1572. [Google Scholar] [CrossRef]
- Fernandez-Calvino, L.; Faulkner, C.; Walshaw, J.; Saalbach, G.; Bayer, E.; Benitez-Alfonso, Y.; Maule, A. Arabidopsis Plasmodesmal Proteome. PLoS ONE 2011, 6, e18880. [Google Scholar] [CrossRef] [PubMed]
- Tilsner, J.; Kriechbaumer, V. Reticulons 3 and 6 Interact with Viral Movement Proteins. Mol. Plant Pathol. 2022, 23, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Ungermann, C.; Kümmel, D. Structure of Membrane Tethers and Their Role in Fusion. Traffic 2019, 20, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Tilsner, J.; Nicolas, W.; Rosado, A.; Bayer, E.M. Staying Tight: Plasmodesmal Membrane Contact Sites and the Control of Cell-to-Cell Connectivity in Plants. Annu. Rev. Plant Biol. 2016, 67, 337–364. [Google Scholar] [CrossRef]
- Rosado, A.; Bayer, E.M. Geometry and Cellular Function of Organelle Membrane Interfaces. Plant Physiol. 2021, 185, 650–662. [Google Scholar] [CrossRef]
- Brault, M.L.; Petit, J.D.; Immel, F.; Nicolas, W.J.; Glavier, M.; Brocard, L.; Gaston, A.; Fouché, M.; Hawkins, T.J.; Crowet, J.-M.; et al. Multiple C2 Domains and Transmembrane Region Proteins (MCTPs) Tether Membranes at Plasmodesmata. EMBO Rep. 2019, 20, e47182. [Google Scholar] [CrossRef]
- Pérez-Sancho, J.; Smokvarska, M.; Glavier, M.; Sritharan, S.; Dubois, G.; Dietrich, V.; Platre, M.P.; Li, Z.P.; Paterlini, A.; Moreau, H.; et al. Plasmodesmata Act as Unconventional Membrane Contact Sites Regulating Inter-Cellular Molecular Exchange in Plants. bioRxiv 2023. [Google Scholar]
- Benitez-Fuente, F.; Botella, M.A. Biological Roles of Plant Synaptotagmins. Eur. J. Cell Biol. 2023, 102, 151335. [Google Scholar] [CrossRef]
- Schapire, A.L.; Voigt, B.; Jasik, J.; Rosado, A.; Lopez-Cobollo, R.; Menzel, D.; Salinas, J.; Mancuso, S.; Valpuesta, V.; Baluska, F.; et al. Arabidopsis Synaptotagmin 1 Is Required for the Maintenance of Plasma Membrane Integrity and Cell Viability. Plant Cell 2008, 20, 3374–3388. [Google Scholar] [CrossRef]
- Ishikawa, K.; Tamura, K.; Fukao, Y.; Shimada, T. Structural and Functional Relationships between Plasmodesmata and Plant Endoplasmic Reticulum—Plasma Membrane Contact Sites Consisting of Three Synaptotagmins. New Phytol. 2020, 226, 798–808. [Google Scholar] [CrossRef]
- Uchiyama, A.; Shimada-Beltran, H.; Levy, A.; Zheng, J.Y.; Javia, P.A.; Lazarowitz, S.G. The Arabidopsis Synaptotagmin SYTA Regulates the Cell-to-Cell Movement of Diverse Plant Viruses. Front. Plant Sci. 2014, 5, 584. [Google Scholar] [CrossRef]
- Tilney, L.G.; Cooke, T.J.; Connelly, P.S.; Tilney, M.S. The Distribution of Plasmodesmata and Its Relationship to Morphogenesis in Fern Gametophytes. Development 1990, 110, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Gunning, B.E.S. Age-Related and Origin-Related Control of the Numbers of Plasmodesmata in Cell Walls of Developing Azolla Roots. Planta 1978, 143, 181–190. [Google Scholar] [CrossRef]
- Wu, S.; O’Lexy, R.; Xu, M.; Sang, Y.; Chen, X.; Yu, Q.; Gallagher, K.L. Symplastic Signaling Instructs Cell Division, Cell Expansion, and Cell Polarity in the Ground Tissue of Arabidopsis thaliana Roots. Proc. Natl. Acad. Sci. USA 2016, 113, 11621–11626. [Google Scholar] [CrossRef]
- Zhu, T.; Lucas, W.J.; Rost, T.L. Directional Cell-to-Cell Communication in the Arabidopsis Root Apical Meristem I. An Ultrastructural and Functional Analysis. Protoplasma 1998, 203, 35–47. [Google Scholar] [CrossRef]
- Seagull, R.W. Differences in the Frequency and Disposition of Plasmodesmata Resulting from Root Cell Elongation. Planta 1983, 159, 497–504. [Google Scholar] [CrossRef]
- Faulkner, C.; Akman, O.E.; Bell, K.; Jeffree, C.; Oparka, K. Peeking into Pit Fields: A Multiple Twinning Model of Secondary Plasmodesmata Formation in Tobacco. Plant Cell 2008, 20, 1504–1518. [Google Scholar] [CrossRef]
- Knox, J.P.; Benitez-Alfonso, Y. Roles and Regulation of Plant Cell Walls Surrounding Plasmodesmata. Curr. Opin. Plant Biol. 2014, 22, 93–100. [Google Scholar] [CrossRef]
- Amsbury, S.; Kirk, P.; Benitez-Alfonso, Y. Emerging Models on the Regulation of Intercellular Transport by Plasmodesmata-Associated Callose. J. Exp. Bot. 2017, 69, 105–115. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Brunkard, J.O.; Choi, Y.G.; Zambryski, P.C. Organelle—Nucleus Cross-Talk Regulates Plant Intercellular Communication via Plasmodesmata. Proc. Natl. Acad. Sci. USA 2011, 108, E1451–E1460. [Google Scholar] [CrossRef]
- Ehlers, K.; van Bel, A.J.E. Dynamics of Plasmodesmal Connectivity in Successive Interfaces of the Cambial Zone. Planta 2010, 231, 371–385. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Zambryski, P.C. Loss of INCREASED SIZE EXCLUSION LIMIT (ISE)1 or ISE2 Increases the Formation of Secondary Plasmodesmata. Curr. Biol. 2010, 20, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Imaichi, R.; Hiratsuka, R. Evolution of Shoot Apical Meristem Structures in Vascular Plants with Respect to Plasmodesmatal Network. Am. J. Bot. 2007, 94, 1911–1921. [Google Scholar] [CrossRef]
- Kim, I.; Hempel, F.D.; Sha, K.; Pfluger, J.; Zambryski, P.C. Identification of a Developmental Transition in Plasmodesmatal Function during Embryogenesis in Arabidopsis thaliana. Development 2002, 129, 1261–1272. [Google Scholar] [CrossRef]
- Stonebloom, S.; Burch-Smith, T.; Kim, I.; Meinke, D.; Mindrinos, M.; Zambryski, P. Loss of the Plant DEAD-Box Protein ISE1 Leads to Defective Mitochondria and Increased Cell-to-Cell Transport via Plasmodesmata. Proc. Natl. Acad. Sci. USA 2009, 106, 17229–17234. [Google Scholar] [CrossRef] [PubMed]
- Carlotto, N.; Wirth, S.; Furman, N.; Ferreyra Solari, N.; Ariel, F.; Crespi, M.; Kobayashi, K. The Chloroplastic DEVH-Box RNA Helicase INCREASED SIZE EXCLUSION LIMIT 2 Involved in Plasmodesmata Regulation Is Required for Group II Intron Splicing. Plant Cell Environ. 2016, 39, 165–173. [Google Scholar] [CrossRef]
- Stonebloom, S.; Brunkard, J.O.; Cheung, A.C.; Jiang, K.; Feldman, L.; Zambryski, P. Redox States of Plastids and Mitochondria Differentially Regulate Intercellular Transport via Plasmodesmata. Plant Physiol. 2012, 158, 190–199. [Google Scholar] [CrossRef]
- Ganusova, E.E.; Reagan, B.C.; Fernandez, J.C.; Azim, M.F.; Sankoh, A.F.; Freeman, K.M.; McCray, T.N.; Patterson, K.; Kim, C.; Burch-Smith, T.M. Chloroplast-to-Nucleus Retrograde Signalling Controls Intercellular Trafficking via Plasmodesmata Formation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190408. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Alfonso, Y.; Cilia, M.; San Roman, A.; Thomas, C.; Maule, A.; Hearn, S.; Jackson, D. Control of Arabidopsis Meristem Development by Thioredoxin-Dependent Regulation of Intercellular Transport. Proc. Natl. Acad. Sci. USA 2009, 106, 3615–3620. [Google Scholar] [CrossRef]
- Carlotto, N.; Robles-Luna, G.; Nedo, A.; Wang, X.; Attorresi, A.; Caplan, J.; Lee, J.Y.; Kobayashi, K. Evidence for Reduced Plasmodesmata Callose Accumulation in Nicotiana benthamiana Leaves with Increased Symplastic Cell-to-Cell Communication Caused by RNA Processing Defects of Chloroplasts. Plant Physiol. Biochem. 2022, 179, 58–64. [Google Scholar] [CrossRef]
- Azim, M.F.; Burch-Smith, T.M. Organelles-Nucleus-Plasmodesmata Signaling (ONPS): An Update on Its Roles in Plant Physiology, Metabolism and Stress Responses. Curr. Opin. Plant Biol. 2020, 58, 48–59. [Google Scholar] [CrossRef]
- Brunkard, J.O.; Xu, M.; Scarpin, M.R.; Chatterjee, S.; Shemyakina, E.A.; Goodman, H.M.; Zambryski, P. TOR Dynamically Regulates Plant Cell-Cell Transport. Proc. Natl. Acad. Sci. USA 2020, 117, 5049–5058. [Google Scholar] [CrossRef]
- Ehlers, K.; Westerloh, M.G. Developmental Control of Plasmodesmata Frequency, Structure, and Function. In Symplasmic Transport in Vascular Plants; Sokołowska, K., Sowiński, P., Eds.; Springer: New York, NY, USA, 2013; pp. 41–82. ISBN 978-1-4614-7765-5. [Google Scholar]
- Kollmann, R.; Glockmann, C. Studies on Graft Unions. Protoplasma 1991, 165, 71–85. [Google Scholar] [CrossRef]
- Chambaud, C.; Cookson, S.J.; Ollat, N.; Bayer, E.; Brocard, L. A Correlative Light Electron Microscopy Approach Reveals Plasmodesmata Ultrastructure at the Graft Interface. Plant Physiol. 2022, 188, 44–55. [Google Scholar] [CrossRef]
- Ostermeyer, G.P.; Jensen, K.H.; Franzen, A.R.; Peters, W.S.; Knoblauch, M. Diversity of Funnel Plasmodesmata in Angiosperms: The Impact of Geometry on Plasmodesmal Resistance. Plant J. 2022, 110, 707–719. [Google Scholar] [CrossRef]
- Oparka, K.J.; Roberts, A.G.; Boevink, P.; Cruz, S.S.; Roberts, I.; Pradel, K.S.; Imlau, A.; Kotlizky, G.; Sauer, N.; Epel, B. Simple, but Not Branched, Plasmodesmata Allow the Nonspecific Trafficking of Proteins in Developing Tobacco Leaves. Cell 1999, 97, 743–754. [Google Scholar] [CrossRef]
- Johnston, M.G.; Breakspear, A.; Samwald, S.; Zhang, D.; Papp, D.; Faulkner, C.; de Keijzer, J. Comparative Phyloproteomics Identifies Conserved Plasmodesmal Proteins. J. Exp. Bot. 2023, 74, 1821–1835. [Google Scholar] [CrossRef]
- Wu, S.-W.; Kumar, R.; Iswanto, A.B.B.; Kim, J.-Y. Callose Balancing at Plasmodesmata. J. Exp. Bot. 2018, 69, 5325–5339. [Google Scholar] [CrossRef]
- De Storme, N.; Geelen, D. Callose Homeostasis at Plasmodesmata: Molecular Regulators and Developmental Relevance. Front. Plant Sci. 2014, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Zavaliev, R.; Ueki, S.; Epel, B.L.; Citovsky, V. Biology of Callose (β-1,3-Glucan) Turnover at Plasmodesmata. Protoplasma 2011, 248, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Radford, J.E.; White, R.G. Localization of a Myosin-like Protein to Plasmodesmata. Plant J. 1998, 14, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R.; et al. Callose Biosynthesis Regulates Symplastic Trafficking during Root Development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Rinne, P.L.H.; Welling, A.; Vahala, J.; Ripel, L.; Ruonala, R.; Kangasjärvi, J.; van der Schoot, C. Chilling of Dormant Buds Hyperinduces FLOWERING LOCUS T and Recruits GA-Inducible 1,3-β-Glucanases to Reopen Signal Conduits and Release Dormancy in Populus. Plant Cell 2011, 23, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Lee, J.-Y. Arabidopsis Callose Synthases CalS1/8 Regulate Plasmodesmal Permeability during Stress. Nat. Plants 2016, 2, 16034. [Google Scholar] [CrossRef] [PubMed]
- Tee, E.E.; Johnston, M.G.; Papp, D.; Faulkner, C. A PDLP-NHL3 Complex Integrates Plasmodesmal Immune Signaling Cascades. Proc. Natl. Acad. Sci. USA 2023, 120, e2216397120. [Google Scholar] [CrossRef] [PubMed]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M.; et al. Photoperiodic Control of Seasonal Growth Is Mediated by ABA Acting on Cell-Cell Communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef]
- Pan, W.; Li, J.; Du, Y.; Zhao, Y.; Xin, Y.; Wang, S.; Liu, C.; Lin, Z.; Fang, S.; Yang, Y.; et al. Epigenetic Silencing of Callose Synthase by VIL1 Promotes Bud-Growth Transition in Lily Bulbs. Nat. Plants 2023, 9, 1451–1467. [Google Scholar] [CrossRef]
- Guseman, J.M.; Lee, J.S.; Bogenschutz, N.L.; Peterson, K.M.; Virata, R.E.; Xie, B.; Kanaoka, M.M.; Hong, Z.; Torii, K.U. Dysregulation of Cell-to-Cell Connectivity and Stomatal Patterning by Loss-of-Function Mutation in Arabidopsis Chorus (Glucan Synthase-like 8). Development 2010, 137, 1731–1741. [Google Scholar] [CrossRef]
- Ali, O.; Cheddadi, I.; Landrein, B.; Long, Y. Revisiting the Relationship between Turgor Pressure and Plant Cell Growth. New Phytol. 2023, 238, 62–69. [Google Scholar] [CrossRef]
- Beauzamy, L.; Nakayama, N.; Boudaoud, A. Flowers under Pressure: Ins and Outs of Turgor Regulation in Development. Ann. Bot. 2014, 114, 1517–1533. [Google Scholar] [CrossRef] [PubMed]
- Rygol, J.; Pritchard, J.; Zhu, J.J.; Tomos, A.D.; Zimmermann, U. Transpiration Induces Radial Turgor Pressure Gradients in Wheat and Maize Roots. Plant Physiol. 1993, 103, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Cheddadi, I.; Mosca, G.; Mirabet, V.; Dumond, M.; Kiss, A.; Traas, J.; Godin, C.; Boudaoud, A. Cellular Heterogeneity in Pressure and Growth Emerges from Tissue Topology and Geometry. Curr. Biol. 2020, 30, 1504–1516.e8. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, M.; Hughes, A.; Morris, R.J. An Update on Passive Transport in and out of Plant Cells. Plant Physiol. 2021, 187, 1973–1984. [Google Scholar] [CrossRef]
- Ding, D.-Q.; Tazawa, M. Influence of Cytoplasmic Streaming and Turgor Pressure Gradient on the Transnodal Transport of Rubidium and Electrical Conductance in Chara corallina. Plant Cell Physiol. 1989, 30, 739–748. [Google Scholar] [CrossRef]
- Reid, R.J.; Overall, R.L. Intercellular Communication in Chara: Factors Affecting Transnodal Electrical Resistance and Solute Fluxes. Plant Cell Environ. 1992, 15, 507–517. [Google Scholar] [CrossRef]
- Oparka, K.J.; Prior, D.A.M. Direct Evidence for Pressure-Generated Closure of Plasmodesmata. Plant J. 1992, 2, 741–750. [Google Scholar] [CrossRef]
- Park, K.; Knoblauch, J.; Oparka, K.; Jensen, K.H. Controlling Intercellular Flow through Mechanosensitive Plasmodesmata Nanopores. Nat. Commun. 2019, 10, 3564. [Google Scholar] [CrossRef]
- White, R.G.; Badelt, K.; Overall, R.L.; Vesk, M. Actin Associated with Plasmodesmata. Protoplasma 1994, 180, 169–184. [Google Scholar] [CrossRef]
- Radford, J.E.; White, R.G. Inhibitors of Myosin, but Not Actin, Alter Transport through Tradescantia Plasmodesmata. Protoplasma 2011, 248, 205–216. [Google Scholar] [CrossRef]
- Diao, M.; Huang, S. An Update on the Role of the Actin Cytoskeleton in Plasmodesmata: A Focus on Formins. Front. Plant Sci. 2021, 12, 647123. [Google Scholar] [CrossRef]
- Zhao, Y.; Pan, W.; Xin, Y.; Wu, J.; Li, R.; Shi, J.; Long, S.; Qu, L.; Yang, Y.; Yi, M.; et al. Regulating Bulb Dormancy Release and Flowering in Lily through Chemical Modulation of Intercellular Communication. Plant Methods 2023, 19, 136. [Google Scholar] [CrossRef]
- White, R.G.; Barton, D.A. The Cytoskeleton in Plasmodesmata: A Role in Intercellular Transport? J. Exp. Bot. 2011, 62, 5249–5266. [Google Scholar] [CrossRef]
- van Gisbergen, P.A.C.; Bezanilla, M. Plant Formins: Membrane Anchors for Actin Polymerization. Trends Cell Biol. 2013, 23, 227–233. [Google Scholar] [CrossRef]
- Diao, M.; Ren, S.; Wang, Q.; Qian, L.; Shen, J.; Liu, Y.; Huang, S. Arabidopsis Formin 2 Regulates Cell-to-Cell Trafficking by Capping and Stabilizing Actin Filaments at Plasmodesmata. eLife 2018, 7, e36316. [Google Scholar] [CrossRef] [PubMed]
- Tilsner, J.; Amari, K.; Torrance, L. Plasmodesmata Viewed as Specialised Membrane Adhesion Sites. Protoplasma 2011, 248, 39–60. [Google Scholar] [CrossRef]
- Wang, P.; Hawkins, T.J.; Richardson, C.; Cummins, I.; Deeks, M.J.; Sparkes, I.; Hawes, C.; Hussey, P.J. The Plant Cytoskeleton, NET3C, and VAP27 Mediate the Link between the Plasma Membrane and Endoplasmic Reticulum. Curr. Biol. 2014, 24, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Crawford, K.M.; Zambryski, P.C. Subcellular Localization Determines the Availability of Non-Targeted Proteins to Plasmodesmatal Transport. Curr. Biol. 2000, 10, 1032–1040. [Google Scholar] [CrossRef]
- Gallagher, K.L.; Paquette, A.J.; Nakajima, K.; Benfey, P.N. Mechanisms Regulating SHORT-ROOT Intercellular Movement Nontargeted Movement Involves a Protein Moving with-out Interacting with the Plasmodesmal Apparatus. In This Case, the Process Resembles Passive Diffusion, as For. Curr. Biol. 2004, 14, 1847–1851. [Google Scholar] [CrossRef]
- Schönknecht, G.; Brown, J.E.; Verchot-Lubicz, J. Plasmodesmata Transport of GFP Alone or Fused to Potato Virus X TGBp1 Is Diffusion Driven. Protoplasma 2008, 232, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Dashevskaya, S.; Kopito, R.B.; Friedman, R.; Elbaum, M.; Epel, B.L. Diffusion of Anionic and Neutral GFP Derivatives through Plasmodesmata in Epidermal Cells of Nicotiana benthamiana. Protoplasma 2008, 234, 13–23. [Google Scholar] [CrossRef]
- Sessions, A.S.; Yanofsky, M.F.; Weigel, D. Cell-Cell Signaling and Movement by the Floral Transcription Factors LEAFY and APETALA1. Science 2000, 289, 779–781. [Google Scholar] [CrossRef]
- Wu, X.; Dinneny, J.R.; Crawford, K.M.; Rhee, Y.; Citovsky, V.; Zambryski, P.C.; Weigel, D. Modes of Intercellular Transcription Factor Movement in the Arabidopsis Apex. Development 2003, 130, 3735–3745. [Google Scholar] [CrossRef]
- Moyroud, E.; Kusters, E.; Monniaux, M.; Koes, R.; Parcy, F. LEAFY Blossoms. Trends Plant Sci. 2010, 15, 346–352. [Google Scholar] [CrossRef]
- Zambryski, P.; Crawford, K. PLASMODESMATA: Gatekeepers for Cell-to-Cell Transport of Developmental Signals in Plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 393–421. [Google Scholar] [CrossRef]
- Rim, Y.; Huang, L.; Chu, H.; Han, X.; Cho, W.K.; Jeon, C.O.; Kim, H.J.; Hong, J.-C.; Lucas, W.J.; Kim, J.-Y. Analysis of Arabidopsis Transcription Factor Families Revealed Extensive Capacity for Cell-to-Cell Movement as Well as Discrete Trafficking Patterns. Mol. Cells 2011, 32, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Colinas, J.; Wang, J.Y.; Mace, D.; Ohler, U.; Benfey, P.N. Transcriptional and Posttranscriptional Regulation of Transcription Factor Expression in Arabidopsis Roots. Proc. Natl. Acad. Sci. USA 2006, 103, 6055–6060. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Veit, B.; Hake, S. Expression of Maize KNOTTED1 Related Homeobox Genes in the Shoot Apical Meristem Predicts Patterns of Morphogenesis in the Vegetative Shoot. Development 1994, 120, 405–413. [Google Scholar] [CrossRef]
- Lucas, W.J.; Bouché-Pillon, S.; Jackson, D.P.; Nguyen, L.; Baker, L.; Ding, B.; Hake, S. Selective Trafficking of KNOTTED1 Homeodomain Protein and Its mRNA through Plasmodesmata. Science 1995, 270, 1980–1983. [Google Scholar] [CrossRef]
- Kim, J.Y.; Yuan, Z.; Jackson, D. Developmental Regulation and Significance of KNOX Protein Trafficking in Arabidopsis. Development 2003, 130, 4351–4362. [Google Scholar] [CrossRef]
- Helariutta, Y.; Fukaki, H.; Wysocka-Diller, J.; Nakajima, K.; Jung, J.; Sena, G.; Hauser, M.T.; Benfey, P.N. The SHORT-ROOT Gene Controls Radial Patterning of the Arabidopsis Root through Radial Signaling. Cell 2000, 101, 555–567. [Google Scholar] [CrossRef]
- Nakajima, K.; Sena, G.; Nawy, T.; Benfey, P.N. Intercellular Movement of the Putative Transcription Factor SHR in Root Patterning. Nature 2001, 413, 307–311. [Google Scholar] [CrossRef]
- Gallagher, K.L.; Benfey, P.N. Both the Conserved GRAS Domain and Nuclear Localization Are Required for SHORT-ROOT Movement. Plant J. 2009, 57, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Welch, D.; Hassan, H.; Blilou, I.; Immink, R.; Heidstra, R.; Scheres, B. Arabidopsis JACKDAW and MAGPIE Zinc Finger Proteins Delimit Asymmetric Cell Division and Stabilize Tissue Boundaries by Restricting SHORT-ROOT Action. Genes Dev. 2007, 21, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Goedhart, J.; Schneijderberg, M.; Terpstra, I.; Shimotohno, A.; Bouchet, B.P.; Akhmanova, A.; Gadella, T.W.J.; Heidstra, R.; Scheres, B.; et al. SCARECROW-LIKE23 and SCARECROW Jointly Specify Endodermal Cell Fate but Distinctly Control SHORT-ROOT Movement. Plant J. 2015, 84, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.J.; De Rybel, B.; Van Mourik, H.; Weijers, D. Regulation of Intercellular Target of Monopteros 7 Protein Transport in the Arabidopsis Root. Development 2018, 145, dev152892. [Google Scholar] [CrossRef] [PubMed]
- Schlereth, A.; Möller, B.; Liu, W.; Kientz, M.; Flipse, J.; Rademacher, E.H.; Schmid, M.; Jürgens, G.; Weijers, D. MONOPTEROS Controls Embryonic Root Initiation by Regulating a Mobile Transcription Factor. Nature 2010, 464, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Kurata, T.; Okada, K.; Wada, T. A Genetic Regulatory Network in the Development of Trichomes and Root Hairs. Annu. Rev. Plant Biol. 2008, 59, 365–386. [Google Scholar] [CrossRef]
- Kurata, T.; Okada, K.; Wada, T. Intercellular Movement of Transcription Factors. Curr. Opin. Plant Biol. 2005, 8, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Zhou, J. Function and Identification of Mobile Transcription Factors. In Short and Long Distance Signaling; Advances in Plant Biology; Kragler, F., Hülskamp, M., Eds.; Springer: New York, NY, USA, 2012; pp. 61–86. ISBN 978-1-4419-1532-0. [Google Scholar]
- Koizumi, K.; Wu, S.; MacRae-Crerar, A.; Gallagher, K.L. An Essential Protein That Interacts with Endosomes and Promotes Movement of the SHORT-ROOT Transcription Factor. Curr. Biol. 2011, 21, 1559–1564. [Google Scholar] [CrossRef]
- Wu, S.; Gallagher, K.L. The Movement of the Non-Cell-Autonomous Transcription Factor, SHORT-ROOT Relies on the Endomembrane System. Plant J. 2014, 80, 396–409. [Google Scholar] [CrossRef]
- Spiegelman, Z.; Lee, C.-M.; Gallagher, K.L. KinG Is a Plant-Specific Kinesin That Regulates Both Intra- and Intercellular Movement of SHORT-ROOT. Plant Physiol. 2018, 176, 392–405. [Google Scholar] [CrossRef]
- Wu, S.; Gallagher, K.L. Intact Microtubules Are Required for the Intercellular Movement of the SHORT-ROOT Transcription Factor. Plant J. 2013, 74, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, Z.; Wu, S.; Gallagher, K.L. A Role for the Endoplasmic Reticulum in the Cell-to-Cell Movement of SHORT-ROOT. Protoplasma 2019, 256, 1455–1459. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, I.; Frantová, N.; Zouhar, J. A Sword or a Buffet: Plant Endomembrane System in Viral Infections. Front. Plant Sci. 2023, 14, 1226498. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, S.; Chen, Y.; Kimberlin, A.N.; Cahoon, E.B.; Yu, B. snRNA 3′ End Processing by a CPSF73-Containing Complex Essential for Development in Arabidopsis. PLOS Biol. 2016, 14, e1002571. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, C.; Song, S.; Teo, Z.W.N.; Shen, L.; Wang, Y.; Jackson, D.; Yu, H. FTIP-Dependent STM Trafficking Regulates Shoot Meristem Development in Arabidopsis. Cell Rep. 2018, 23, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Winter, N.; Kollwig, G.; Zhang, S.; Kragler, F. MPB2C, a Microtubule-Associated Protein, Regulates Non-Cell-Autonomy of the Homeodomain Protein KNOTTED1. Plant Cell 2007, 19, 3001–3018. [Google Scholar] [CrossRef] [PubMed]
- Kragler, F.; Monzer, J.; Xoconostle-Cázares, B.; Lucas, W.J. Peptide Antagonists of the Plasmodesmal Macromolecular Trafficking Pathway. EMBO J. 2000, 19, 2856–2868. [Google Scholar] [CrossRef]
- Kragler, F.; Monzer, J.; Shash, K.; Xoconostle-Cázares, B.; Lucas, W.J. Cell-to-Cell Transport of Proteins: Requirement for Unfolding and Characterization of Binding to a Putative Plasmodesmal Receptor. Plant J. 1998, 15, 367–381. [Google Scholar] [CrossRef]
- Xu, X.M.; Wang, J.; Xuan, Z.; Goldshmidt, A.; Borrill, P.G.M.; Hariharan, N.; Kim, J.Y.; Jackson, D. Chaperonins Facilitate KNOTTED1 Cell-to-Cell Trafficking and Stem Cell Function. Science 2011, 333, 1141–1144. [Google Scholar] [CrossRef]
- Horwich, A.L.; Fenton, W.A.; Chapman, E.; Farr, G.W. Two Families of Chaperonin: Physiology and Mechanism. Annu. Rev. Cell Dev. Biol. 2007, 23, 115–145. [Google Scholar] [CrossRef]
- Liu, L.; Li, C.; Liang, Z.; Yu, H. Characterization of Multiple C2 Domain and Transmembrane Region Proteins in Arabidopsis. Plant Physiol. 2018, 176, 2119–2132. [Google Scholar] [CrossRef]
- Liu, L.; Liu, C.; Hou, X.; Xi, W.; Shen, L.; Tao, Z.; Wang, Y.; Yu, H. FTIP1 Is an Essential Regulator Required for Florigen Transport. PLoS Biol. 2012, 10, 1001313. [Google Scholar] [CrossRef]
- Endo, M.; Yoshida, M.; Sasaki, Y.; Negishi, K.; Horikawa, K.; Daimon, Y.; Kurotani, K.; Notaguchi, M.; Abe, M.; Araki, T. Re-Evaluation of florigen transport kinetics with separation of functions by mutations that uncouple flowering initiation and long-distance transport. Plant Cell Physiol. 2018, 8, 1621–1629. [Google Scholar] [CrossRef]
- Liu, L.; Li, C.; Teo, Z.W.N.; Zhang, B.; Yu, H. The MCTP-SNARE Complex Regulates Florigen Transport in Arabidopsis. Plant Cell 2019, 31, 2475–2490. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Kwak, S.-H.; Nam, K.H.; Schiefelbein, J.; Lee, M.M. QUIRKY Regulates Root Epidermal Cell Patterning through Stabilizing SCRAMBLED to Control CAPRICE Movement in Arabidopsis. Nat. Commun. 2019, 10, 1744. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, D.; Batoux, M.; Fulton, L.; Pfister, K.; Yadav, R.K.; Schellenberg, M.; Schneitz, K. STRUBBELIG Defines a Receptor Kinase-Mediated Signaling Pathway Regulating Organ Development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 9074–9079. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.-H.; Woo, S.; Lee, M.M.; Schiefelbein, J. Distinct Signaling Mechanisms in Multiple Developmental Pathways by the SCRAMBLED Receptor of Arabidopsis. Plant Physiol. 2014, 166, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, M.; Jennings, J.; Johnston, M.G.; Breakspear, A.; Liu, X.; Stark, K.; Morris, R.; De Keijzer, J.; Faulkner, C. Assaying Effector Cell-to-Cell Mobility in Plant Tissues Identifies Hypermobility and Indirect Manipulation of Plasmodesmata. MPMI 2023. [Google Scholar] [CrossRef] [PubMed]
- Arnoux-Courseaux, M.; Coudert, Y. Re-Examining Meristems through the Lens of Evo-Devo. Trends Plant Sci. 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Gombos, S.; Miras, M.; Howe, V.; Xi, L.; Pottier, M.; Jasemi, N.S.K.; Schladt, M.; Ejike, J.O.; Neumann, U.; Hänsch, S.; et al. A High-confidence Physcomitrium Patens Plasmodesmata Proteome by Iterative Scoring and Validation Reveals Diversification of Cell Wall Proteins during Evolution. New Phytol. 2023, 238, 637–653. [Google Scholar] [CrossRef]
- Kitagawa, M.; Fujita, T. Quantitative Imaging of Directional Transport through Plasmodesmata in Moss Protonemata via Single-Cell Photoconversion of Dendra2. J. Plant Res. 2013, 126, 577–585. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schreiber, J.M.; Limpens, E.; de Keijzer, J. Distributing Plant Developmental Regulatory Proteins via Plasmodesmata. Plants 2024, 13, 684. https://doi.org/10.3390/plants13050684
Schreiber JM, Limpens E, de Keijzer J. Distributing Plant Developmental Regulatory Proteins via Plasmodesmata. Plants. 2024; 13(5):684. https://doi.org/10.3390/plants13050684
Chicago/Turabian StyleSchreiber, Joyce M., Erik Limpens, and Jeroen de Keijzer. 2024. "Distributing Plant Developmental Regulatory Proteins via Plasmodesmata" Plants 13, no. 5: 684. https://doi.org/10.3390/plants13050684