
Adaptation Strategies of Seedling Root Response to Nitrogen and Phosphorus Addition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
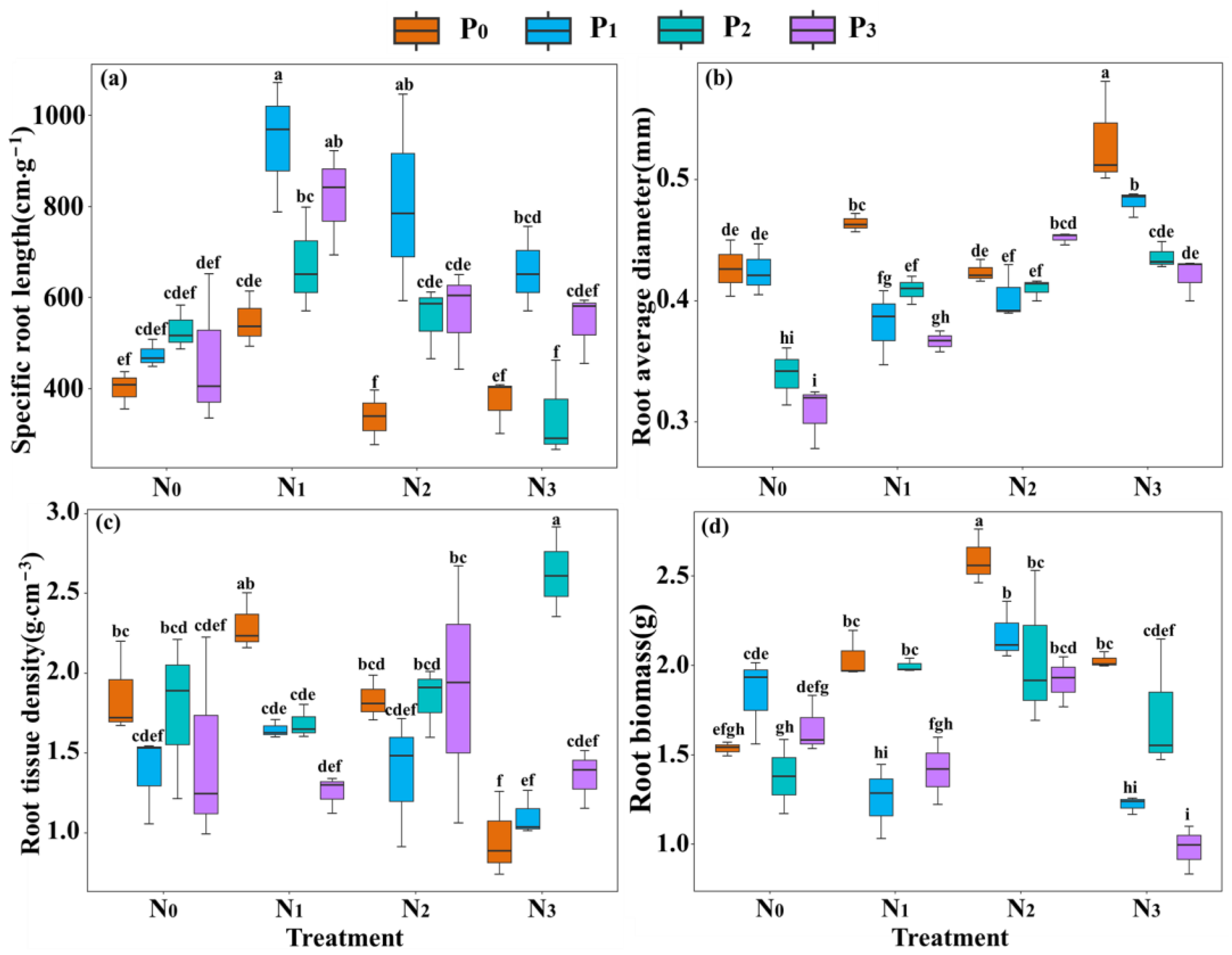
2.1. Root Morphology Traits in Response to Nitrogen and Phosphorus Addition
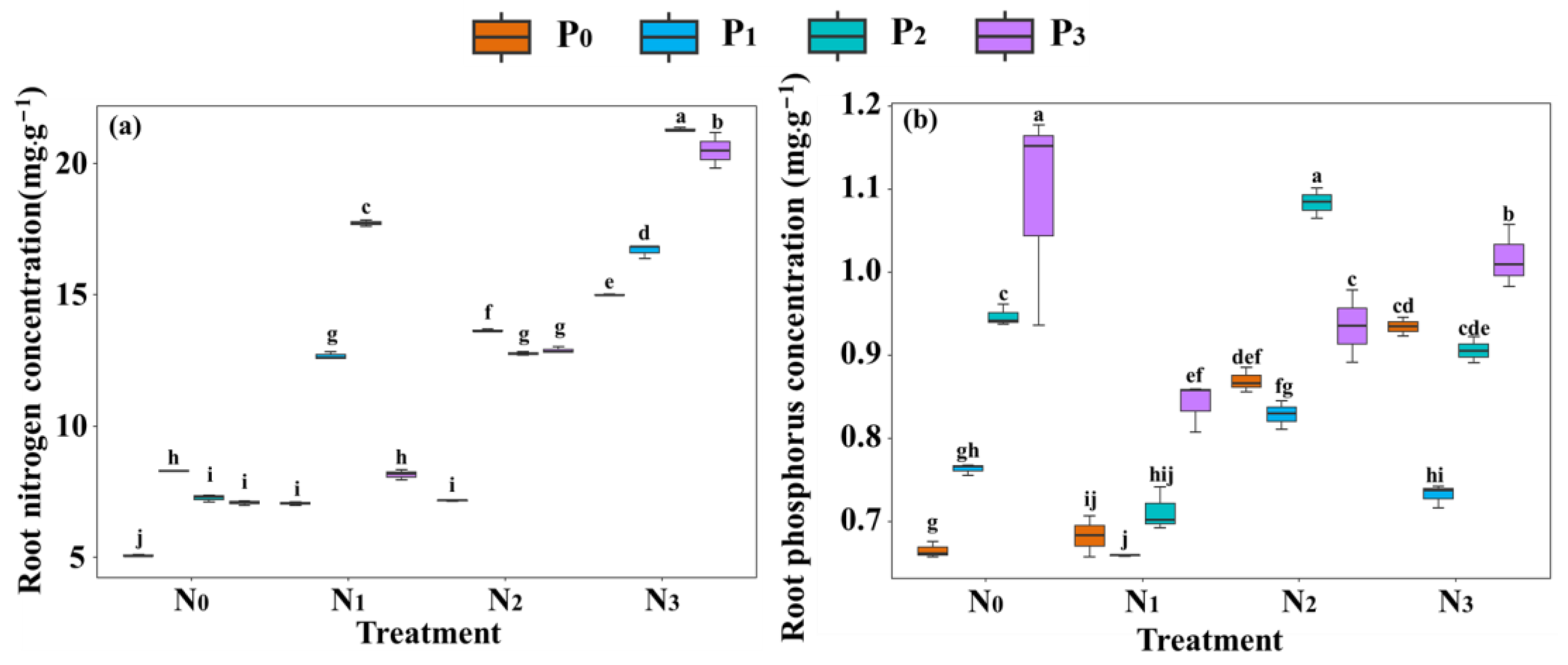
2.2. Response of Root Nitrogen and Phosphorus Content to Nitrogen and Phosphorus Addition
2.3. Resource-Use Strategies of Root Traits in Different Nitrogen and Phosphorus Addition
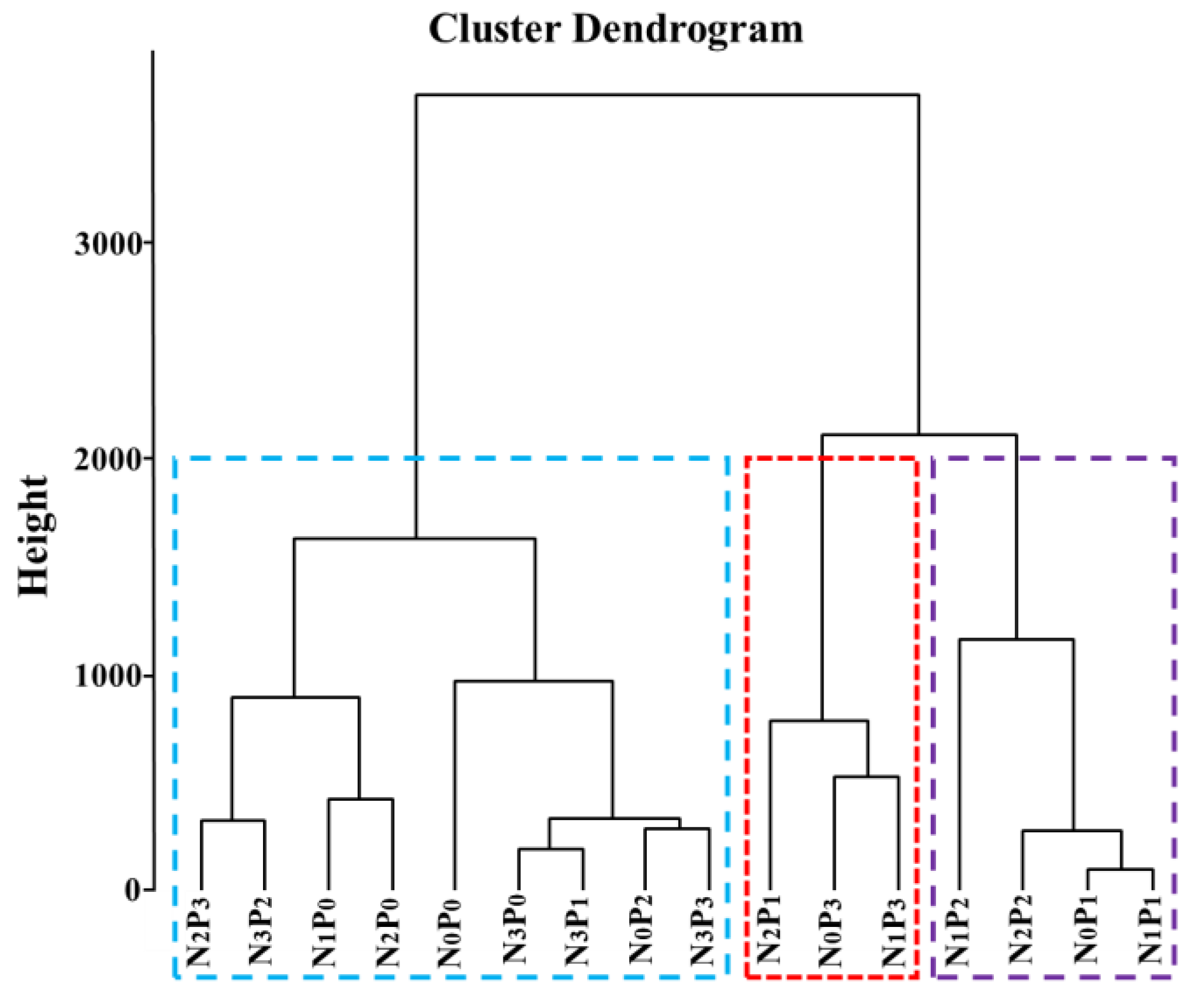
2.4. Comprehensive Evaluation of Nitrogen and Phosphorus Addition on Root Traits
3. Discussion
3.1. Effects of Single Nitrogen and Phosphorus Addition on Root Traits
3.2. Effects of Nitrogen and Phosphorus Interactive Addition on Root Traits
3.3. Trade-Off Strategies of Root in Response to Nitrogen and Phosphorus Addition
4. Materials and Methods
4.1. Materials Collection
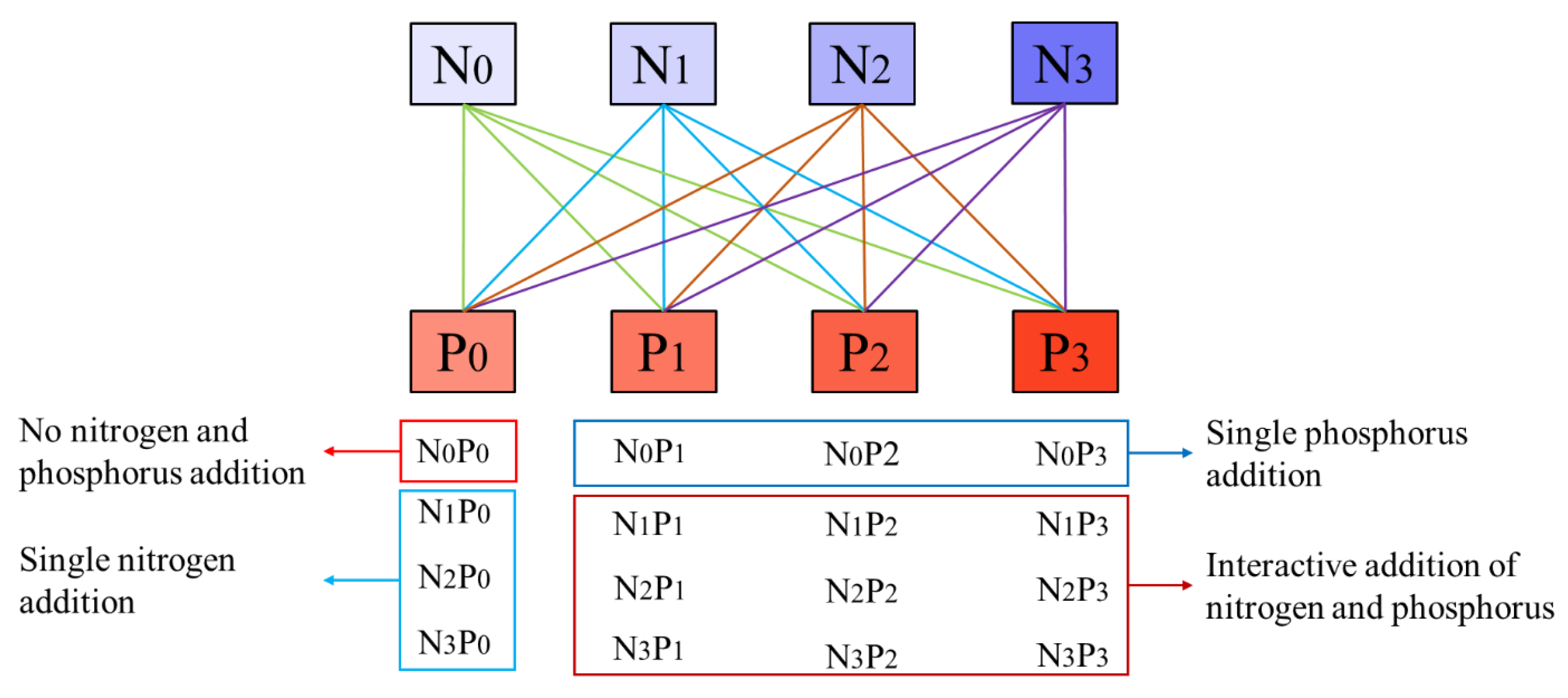
4.2. Experimental Design
4.3. Determination of Seedling Root Traits
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.F.; Yan, X.J.; Dayananda, B.; Luo, Y.L.; Li, J.Q. Frequency-dependent predation and seedling fate: Effect of forest litter on regeneration of the Quercus wutaishanica seedling. Glob. Ecol. Conserv. 2022, 38, e02233. [Google Scholar] [CrossRef]
- Wang, B.; Gu, Z.; Shu, L.F.; Shi, C.M.; Chen, F.; Bai, Y.; Liu, X.D. The characteristics of seedling regeneration and its influencing factors of Pinus tabuliformis under different wildfire severity of Liaoheyuan Nature Reserve. Nat. Hazards 2021, 1, 126–133. [Google Scholar] [CrossRef]
- Kong, D.L.; Ma, C.G.; Zhang, Q.; Li, L.; Chen, X.Y.; Zeng, H.; Guo, D.L. Leading dimensions in absorptive root trait variation across 96 subtropical forest species. New Phytol. 2014, 203, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.S.; Hobbie, S.E.; Reich, P.B.; Yang, Y.S.; Robinson, D. Allometry of fine roots in forest ecosystems. Ecol. Lett. 2019, 22, 322–331. [Google Scholar] [CrossRef]
- Kou, L.; Chen, W.W.; Gao, W.L.; Yang, H.; Wang, H.M.; Li, S.G. Effects of mixture of branch order-based roots and nitrogen addition on root decay in a subtropical pine plantation. Biol. Fertil. Soils 2015, 51, 947–957. [Google Scholar] [CrossRef]
- Adams, T.S.; McCormack, M.L.; Eissenstat, D.M. Foraging strategies in trees of different root morphology: The role of root lifespan. Tree Physiol. 2013, 33, 940–948. [Google Scholar] [CrossRef]
- Zhao, C.T.; Lin, Q.H.; Tian, D.; Ji, C.J.; Shen, H.H.; Fan, D.Y.; Wang, X.P.; Fang, J.Y. Nitrogen addition promotes conservative resource-use strategies via aggravating phosphorus limitation of evergreen trees in subtropical forest. Sci. Total Environ. 2023, 889, 164047. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root structure–function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef]
- Ding, J.X.; Kong, D.L.; Zhang, Z.L.; Cai, Q.X.; Xiao, J.; Liu, Q.; Yin, H.J. Climate and soil nutrients differentially drive multidimensional fine root traits in ectomycorrhizal-dominated alpine coniferous forests. J. Ecol. 2020, 180, 2544–2556. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Wang, R.L.; Wang, Q.F.; Zhao, N.; Xu, Z.W.; Zhu, X.J.; Jiao, C.C.; Yu, G.R.; He, N.P. Different phylogenetic and environmental controls of first-order root morphological and nutrient traits: Evidence of multidimensional root traits. Funct. Ecol. 2018, 32, 29–39. [Google Scholar] [CrossRef]
- Razaq, M.; Salahuddin; Shen, H.; Sher, H.; Zhang, P. Influence of biochar and nitrogen on fine root morphology, physiology, and chemistry of Acer mono. Sci. Rep. 2017, 7, 5367. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Blackwood, C.B. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees. Funct. Ecol. 2015, 29, 796–807. [Google Scholar] [CrossRef]
- Shen, Y.; Umaña, M.N.; Li, W.B.; Miao, F.; Chen, Y.X.; Lu, H.; Yu, S.S. Coordination of leaf, stem and root traits in determining seedling mortality in a subtropical forest. For. Ecol. Manag. 2019, 446, 285–292. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; Vries, F.T.D. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, 6486. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Rivas-ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 18, 3–6. [Google Scholar] [CrossRef]
- Huang, J.Y.; Yu, H.L.; Lin, H.; Zhang, Y.; Searle, E.B.; Yuan, Z.Q. Phosphorus amendment mitigates nitrogen addition-induced phosphorus limitation in two plant species in a desert steppe, China. Plant Soil 2016, 399, 221–232. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.T.; Chen, H.Y.H.; Ruan, H.H. Responses of C: N stoichiometry in plants, soil, and microorganisms to nitrogen addition. Plant Soil 2020, 456, 277–287. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Busta-mante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Terrer, C.; Pellegrini, A.F.A.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Tang, J.; Riley, W.J. Finding Liebig’s law of the minimum. Ecol. Appl. 2021, 31, e0245. [Google Scholar] [CrossRef] [PubMed]
- Ostonen, I.; Lohmus, K.; Helmisaari, H.; Truu, J.; Meel, S. Fine root morphological adaptations in Scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests. Tree Physiol. 2007, 27, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Schleuss, P.; Widdig, M.; Heintz-Buschart, A.; Kirkman, K.; Spohn, M. Interactions of nitrogen and phosphorus cycling promote P acquisition and explain synergistic plant-growth responses. Ecology 2020, 105, e03003. [Google Scholar] [CrossRef]
- Wei, H.X.; Xu, C.Y.; Ma, L.Y.; Duan, J.; Jiang, L.N.; Ren, J.H. Effect of late-season fertilization on nutrient reserves and carbohydrate accumulation in bareroot Larix olgensis seedlings. J. Plant Nutr. 2014, 37, 279–293. [Google Scholar] [CrossRef]
- Wang, G.L.; Fahey, T.J.; Xue, S.; Liu, F. Root morphology and architecture respond to N addition in Pinus tabuliformis, west China. Oecologia 2013, 171, 583–590. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. CRC Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Liu, L.L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef]
- Chen, G.T.; Tu, L.H.; Peng, Y.; Hu, H.L.; Hu, T.X.; Xu, Z.F.; Liu, L.; Tang, Y. Effect of nitrogen additions on root morphology and chemistry in a subtropical bamboo forest. Plant Soil 2017, 412, 441–451. [Google Scholar] [CrossRef]
- Liu, R.Q.; Huang, Z.Y.; Luke McCormack, M.; Zhou, X.H.; Wan, X.H.; Yu, Z.P.; Wang, M.H.; Zheng, L. Plasticity of fine-root functional traits in the litter layer in response to nitrogen addition in a subtropical forest plantation. Plant Soil 2017, 415, 317–330. [Google Scholar] [CrossRef]
- Buajan, S.; Liu, J.F.; He, Z.S.; Feng, X. Effect of gap sizes on specific leaf area and chlorophyll contents at the Castanopsis kawakamii Natural Reserve Forest, China. Forests 2018, 9, 682. [Google Scholar] [CrossRef]
- He, Z.S.; Wang, L.J.; Jiang, L.; Wang, Z.; Liu, J.F.; Xu, D.; Hong, W. Effect of microenvironment on species distribution patterns in the regeneration layer of forest gaps and non-gaps in a subtropical natural forest, China. Forests 2019, 10, 90. [Google Scholar] [CrossRef]
- He, Z.S.; Liu, J.F.; Su, S.J.; Zheng, S.Q.; Xu, D.W.; Wu, Z.Y.; Hong, W.; Wang, L.M. Effects of forest gaps on soil Properties in Castanopsis kawakamii Nature Forest. PLoS ONE 2015, 10, e0141203. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.L.; Liu, H.Y.; Zhao, F.F.; Xu, C.Y. Ecological stoichiometry of N:P:Si in China’s grasslands. Plant Soil 2014, 380, 165–179. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manag. 2011, 262, 2008–2023. [Google Scholar] [CrossRef]
- Zhu, F.F.; Yoh, M.; Gilliam, F.S.; Lu, X.K.; Mo, J.M. Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions. PLoS ONE 2013, 8, e82661. [Google Scholar] [CrossRef]
- Kou, L.; Jiang, L.; Fu, X.; Dai, X.Q.; Wang, H.M.; Li, S.G. Nitrogen deposition increases root production and turnover but slows root decomposition in Pinus elliottii plantations. New Phytol. 2018, 218, 1450–1461. [Google Scholar] [CrossRef]
- Lugli, L.F.; Rosa, J.S.; Andersen, K.M.; Ponzio, R.; Almeida, R.V.; Pires, M.; Cordeiro, A.L.; Cunha, H.F.V.; Martins, N.P.; Assis, R.L.; et al. Rapid responses of root traits and productivity to phosphorus and cation additions in a tropical lowland forest in Amazonia. New Phytol. 2021, 230, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Rui, Y.; Zhang, F.; Rengel, Z.; Shen, J.B. Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification. Field Crops Res. 2010, 119, 355–364. [Google Scholar] [CrossRef]
- Shan, S.; Devens, H.; Fahey, T.J.; Yanai, R.D.; Fisk, M.C. Fine root growth increases in response to nitrogen addition in phosphorus-limited northern hardwood forests. Ecosystems 2022, 25, 1589–1600. [Google Scholar] [CrossRef]
- Xiong, D.P.; Yang, Z.J.; Chen, G.S.; Liu, X.F.; Lin, W.X.; Huang, J.X.; Bowles, F.P.; Lin, C.F.; Xie, J.S.; Li, Y.P.; et al. Interactive effects of warming and nitrogen addition on fine root dynamics of a young subtropical plantation. Soil Biol. Biochem. 2018, 123, 180–189. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Eissenstat, D.M.; Lovett, G.M.; Bowden, R.D.; Rustad, L.E.; Driscoll, C.T. Root stress and nitrogen deposition: Consequences and research priorities. New Phytol. 2013, 197, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Mo, Q.F.; Han, X.G.; Hui, D.F.; Shen, W. Fine root dynamics responses to nitrogen addition depend on root order, soil layer, and experimental duration in a subtropical forest. Biol. Fertil. Soils 2019, 55, 723–736. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.M.; Chen, C.; Li, T.; Bu, Z.J.; Wang, M. Contrasting effects of nitrogen and phosphorus additions on fine root production and morphological traits of different plant functional types in an ombrotrophic peatland. Plant Soil 2023, 490, 451–467. [Google Scholar] [CrossRef]
- Wang, H.L.; Wang, J.; Teng, Z.; Fan, W.; Deng, P.; Wen, Z.; Zhou, K.; Xu, X. Nitrogen and phosphorus additions impact stability of soil organic carbon and nitrogen in subtropical evergreen broad-leaved forest. Eurasian Soil Sci. 2022, 55, 425–436. [Google Scholar] [CrossRef]
- Fort, F.; Cruz, P.; Lecloux, E.; Bittencourt de Oliveira, L.; Stroia, C.; Theau, J.; Jouany, C. Grassland root functional parameters vary according to a community-level resource acquisition-conservation trade-off. J. Veg. Sci. 2016, 27, 749–758. [Google Scholar] [CrossRef]
- Hu, B.; Chu, C.C. Nitrogen–phosphorus interplay: Old story with molecular tale. New Phytol. 2020, 225, 1455–1460. [Google Scholar] [CrossRef] [PubMed]
- Wurzburger, N.; Wright, S.J. Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology 2015, 96, 2137–2146. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, R.P.; Simpson, R.J.; Lambers, H.; Kidd, D.R.; Ryan, M.H. Root morphology acclimation to phosphorus supply by six cultivars of Trifolium subterraneum L. Plant Soil 2017, 412, 21–34. [Google Scholar] [CrossRef]
- Iversen, C.M.; McCormack, M.L.; Powell, A.S.; Blackwood, C.B.; Freschet, G.T.; Kattge, J.; Roumet, C.; Stover, D.B.; Soudzilovskaia, N.A.; Valverde-Barrantes, O.J.; et al. A global fine-root ecology database to address below-ground challenges in plant ecology. New Phytol. 2017, 215, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.; Weigelt, A.; van der Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Valverde-Barrantes, O.J.; Bruelheide, H.; Freschet, G.T.; Iversen, C.M.; et al. The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon–nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Dietze, M.C.; Bhatnagar, J.M. Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks. Glob. Chang. Biol. 2018, 24, 4544–4553. [Google Scholar] [CrossRef] [PubMed]
- Midgley, M.G.; Phillips, R.P. Resource stoichiometry and the biogeochemical consequences of nitrogen deposition in a mixed deciduous forest. Ecology 2016, 97, 3369–3378. [Google Scholar] [CrossRef]
- Zou, Y.; Li, B.; Peñuelas, J.; Sardans, J.; Yu, H.; Chen, X.; Deng, X.; Cheng, D.; Zhong, Q. Response of functional traits in Machilus pauhoi to nitrogen addition is influenced by differences of provenances. For. Ecol. Manag. 2022, 513, 120207. [Google Scholar] [CrossRef]
- Ye, X.; Bu, W.; Hu, X.; Liu, B.; Liang, K.; Chen, F. Species divergence in seedling leaf traits and tree growth response to nitrogen and phosphorus additions in an evergreen broadleaved forest of subtropical China. J. For. Res. 2023, 34, 137–150. [Google Scholar] [CrossRef]
- Báez, S.; Homeier, J. Functional traits determine tree growth and ecosystem productivity of a tropical montane forest: Insights from a long-term nutrient manipulation experiment. Glob. Chang. Biol. 2018, 24, 399–409. [Google Scholar] [CrossRef]
- DeForest, J.L.; Snell, R.S. Tree growth response to shifting soil nutrient economy depends on mycorrhizal associations. New Phytol. 2020, 225, 2557–2566. [Google Scholar] [CrossRef]
- Wu, A.P.; Hu, X.F.; Wang, F.C.; Guo, C.L.; Wang, H.M.; Chen, F.S. Nitrogen deposition and phosphorus addition alter mobility of trace elements in subtropical forests in China. Sci. Total Environ. 2021, 781, 146778. [Google Scholar] [CrossRef]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, X.; Zhu, J.; Wei, X.; Xiao, Q.; Xiao, J.; Jiang, L.; Xu, D.; Shen, C.; Liu, J.; He, Z. Adaptation Strategies of Seedling Root Response to Nitrogen and Phosphorus Addition. Plants 2024, 13, 536. https://doi.org/10.3390/plants13040536
Jin X, Zhu J, Wei X, Xiao Q, Xiao J, Jiang L, Xu D, Shen C, Liu J, He Z. Adaptation Strategies of Seedling Root Response to Nitrogen and Phosphorus Addition. Plants. 2024; 13(4):536. https://doi.org/10.3390/plants13040536
Chicago/Turabian StyleJin, Xing, Jing Zhu, Xin Wei, Qianru Xiao, Jingyu Xiao, Lan Jiang, Daowei Xu, Caixia Shen, Jinfu Liu, and Zhongsheng He. 2024. "Adaptation Strategies of Seedling Root Response to Nitrogen and Phosphorus Addition" Plants 13, no. 4: 536. https://doi.org/10.3390/plants13040536