
Physiological and Biochemical Aspects of Silicon-Mediated Resistance in Maize against Maydis Leaf Blight
 and
and
Abstract
:1. Introduction
2. Results
2.1. Analysis of Variance
2.2. Foliar Si and K Concentrations and X-ray Microanalysis
2.3. Symptoms of MLB and Severity
2.4. Leaf Gas Exchange Parameters
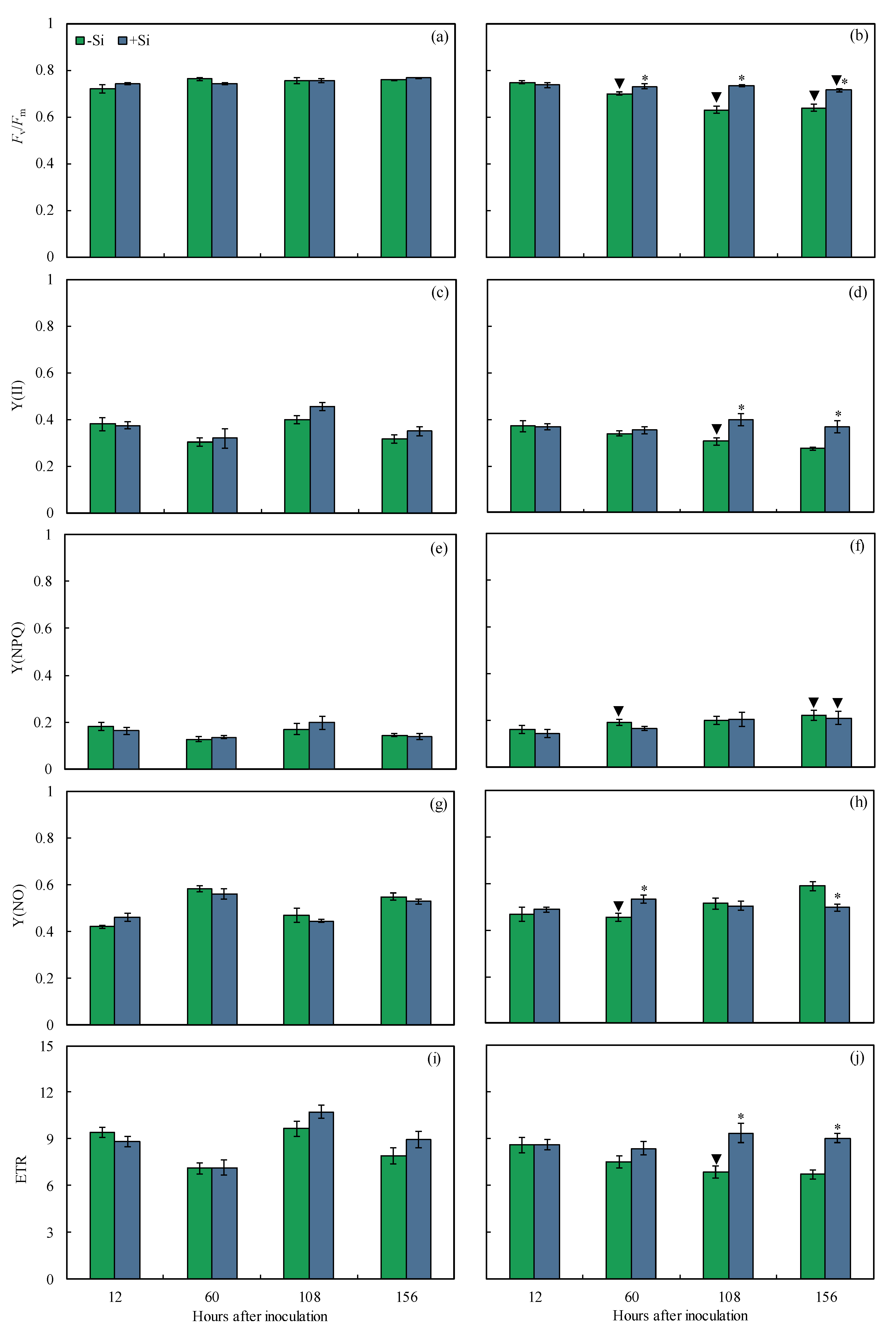
2.5. Imaging and Quantification of Chl a Fluorescence Parameters
2.6. Photosynthetic Pigments
2.7. Carbohydrates
2.8. Concentrations of MDA, H2O2, and O2•−
2.9. Activities of Defense Enzymes
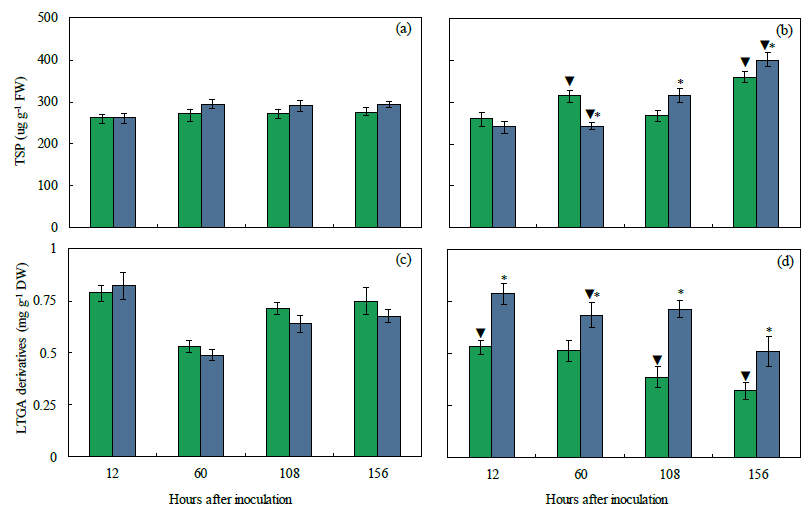
2.10. Concentrations of TSP and LTGA Derivatives
2.11. Antioxidant Enzymes
3. Discussion
4. Material and methods
4.1. Plant Material, Fertilizantion, and Growth Conditions
4.2. Inoculum Production and Plant Inoculation
4.3. Evaluation of MLB Severity
4.4. Determination of Foliar Si and K Concentrations and Their Detection Using X-ray Microanalysis
4.5. Determination of Leaf Gas Exchange Parameters
4.6. Imaging and Quantification of Chl a Fluorescence Parameters
4.7. Determination of Photosynthetic Pigments Concentration
4.8. Biochemical Assays
4.8.1. Determining Sugars and Starch Concentrations
4.8.2. Determining MDA Concentration
4.8.3. Determining H2O2 and O2•− Concentrations
4.8.4. Determining Defense-Related Enzymes Activities
4.8.5. Determining TSP and LTGA Derivatives
4.8.6. Determining Antioxidant Enzyme Activities
4.9. Experimental Design and Data Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global Maize Production, Consumption and Trade: Trends and R&D Implications. Food Sec. 2022, 14, 1295–1319. [Google Scholar]
- Munkvold, G.P.; White, D.G. Compendium of Corn Diseases, 4th ed.; The American Phytopathological Society: Saint Paul, MN, USA, 2016; 165p. [Google Scholar]
- Kumar, B.; Choudhary, M.; Kumar, K.; Kumar, P.; Kumar, S.; Bagaria, P.K.; Sharma, M.; Lahkar, C.; Singh, B.K.; Pradhan, H.; et al. Maydis Leaf Blight of Maize: Update on Status, Sustainable Management and Genetic Architecture of its Resistance. Physiol. Mol. Plant Pathol. 2022, 121, 101889. [Google Scholar] [CrossRef]
- Dai, Y.; Gan, L.; Ruan, H.; Shi, N.; Du, Y.; Chen, F.; Yang, X. Characterization of Natural Isolates of Bipolaris maydis Associated with Mating Types, Genetic Diversity, and Pathogenicity in Fujian Province, China. Plant Dis. 2020, 104, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Manamgoda, D.S.; Rossman, A.Y.; Castlebury, L.A.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Hyde, K.D. The Genus Bipolaris. Stud. Mycol. 2014, 79, 221–288. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, H. Ultrastructure of Penetration by Helminthosporium maydis. Physiol. Plant Pathol. 1977, 11, 171–178. [Google Scholar] [CrossRef]
- Smith, D.R.; Hooker, A.L.; Lim, S.M. Physiologic Races of Helminthosporium maydis. Plant Dis. Report 1970, 54, 819–822. [Google Scholar]
- Meshram, S.; Gogoi, R.; Bashyal, B.M.; Kumar, A.; Mandal, P.K.; Hossain, F. Comparative Transcriptome Analysis of Fungal Pathogen Bipolaris maydis to Understand Pathogenicity Behavior on Resistant and Susceptible Non-CMS Maize Genotypes. Front. Microbiol. 2022, 13, 837056. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Gan, L.; Ruan, H.; Shi, N.; Du, Y.; Liao, L.; Wei, Z.; Teng, Z.; Chen, F.; Yang, X. Sensitivity of Cochliobolus heterostrophus to Three Demethylation Inhibitor Fungicides, Propiconazole, Diniconazole and Prochloraz, and Their Efficacy against Southern Corn Leaf Blight in Fujian Province, China. Eur. J. Plant Pathol. 2018, 152, 447–459. [Google Scholar] [CrossRef]
- Kumar, C.; Chand, P.; Choudhary, C.; Akhtar, N.; Rai, B. In Vitro Evaluation of Fungicides, Botanicals and Bio-Agents against the Maydis Leaf Light Disease of Maize Caused by Helminthosporium maydis. Pharma Innov. 2021, 10, 399–406. [Google Scholar]
- Tripathi, R.; Tewari, R.; Singh, K.P.; Keswani, C.; Minkina, T.; Srivastava, A.K.; De Corato, U.; Sansinenea, E. Plant Mineral Nutrition and Disease Resistance: A Significant Linkage for Sustainable Crop Protection. Front. Plant Sci. 2022, 13, 883970. [Google Scholar] [CrossRef]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef]
- Debona, D.; Datnoff, L.; Rodrigues, F.A. Silicon and Plant Disease. In Mineral Nutrition and Plant Disease; In Datnoff, L.E., Elmer, W.H., Rodrigues, F.A., Eds.; The American Phytopathological Society: Saint. Paul, MN, USA, 2023; pp. 381–424. [Google Scholar]
- Zellner, W.; Tubaña, B.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Plant Stress Reduction and why this Element is not Used Routinely for Managing Plant Health. Plant Dis. 2021, 105, 2033–2049. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A Cooperative System of Silicon Transport in Plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of Maize Silicon Influx Transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Datnoff, L.E. Silicon and Plant Diseases; Springer: Cham, Switzerland, 2015; 148p. [Google Scholar]
- Rodrigues, F.A.; Bispo, W.; Aucique-Pérez, C.E. The Effect of Silicon on Plant Photosynthesis during Pathogens Infection. In Nafees Khan (Org.). Photosynthesis: Functional Genomics, Physiological Processes and Environmental Issues, 1st ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2014; Volume 1, pp. 211–220. [Google Scholar]
- Hawerroth, C.; Araujo, L.; Bermúdez-Cardona, M.B.; Silveira, P.R.; Wordell Filho, J.A.; Rodrigues, F.A. Silicon-Mediated Maize Resistance to Macrospora Leaf Spot. Trop. Plant Pathol. 2019, 44, 192–196. [Google Scholar] [CrossRef]
- Silveira, P.R.; Aucique-Pérez, C.E.; Cruz, M.F.A.; Rodrigues, F.A. Biochemical and Physiological Changes in Maize Plants Supplied with Silicon and Infected by Exserohilum turcicum. J. Phytopathol. 2021, 169, 393–408. [Google Scholar] [CrossRef]
- Berger, S.; Sinha, A.K.; Roitsch, T. Plant Physiology Meets Phytopathology: Plant Primary Metabolism and Plant-Pathogen Interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef] [PubMed]
- Scholes, J.; Rolfe, S. Chlorophyll Fluorescence Imaging as Tool for Understanding the Impact of Fungal Diseases on Plant Performance: A Phenomics Perspective. Funct. Plant Biol. 2009, 36, 880–892. [Google Scholar] [CrossRef] [PubMed]
- Kumudini, B.S.; Jayamohan, N.S.; Patil, S.V.; Govardhana, M. Primary Plant Metabolism During Plant-Pathogen Interactions and Its Role in Defense; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128126899. [Google Scholar]
- Domiciano, G.P.; Cacique, I.S.; Freitas, C.C.; Filippi, M.C.C.; DaMatta, F.M.; Vale, F.X.R.; Rodrigues, F.A. Alterations in Gas Exchange and Oxidative Metabolism in Rice Leaves Infected by Pyricularia oryzae are Attenuated by Silicon. Phytopathology 2015, 105, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Tatagiba, S.D.; Damatta, F.M.; Rodrigues, F.A. Silicon Partially Preserves the Photosynthetic Performance of Rice Plants Infected by Monographella albescens. Ann. Appl. Biol. 2016, 168, 111–121. [Google Scholar] [CrossRef]
- Aucique-Pérez, C.E.; Rodrigues, F.A.; Moreira, W.; Damatta, F.M. Leaf Gas Exchange and Chlorophyll a Fluorescence in Wheat Plants Supplied with Silicon and Infected with Pyricularia oryzae. Biochem. Cell Biol. Leaf 2014, 104, 143–149. [Google Scholar]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-Selective Toxins Produced by the Plant Pathogenic Fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef]
- Silva, B.N.; Picanço, B.B.M.; Hawerroth, C.; Silva, L.C.; Rodrigues, F.A. Physiological and Biochemical Insights into Induced Resistance on Tomato against Septoria Leaf Spot by a Phosphite Combined with Free Amino Acids. Physiol. Mol. Plant Pathol. 2022, 120, 101854. [Google Scholar] [CrossRef]
- Calatayud, A.; San Bautista, A.; Pascual, B.; Maroto, J.V.; López-Galarza, S. Use of Chlorophyll Fluorescence Imaging as Diagnostic Technique to Predict Compatibility in Melon Graft. Sci. Hortic. 2013, 149, 13–18. [Google Scholar] [CrossRef]
- Rochaix, J.D. Regulation of Photosynthetic Electron Transport. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 375–383. [Google Scholar] [CrossRef]
- Rastogi, A.; Yadav, S.; Hussain, S.; Kataria, S.; Hajihashemi, S.; Kumari, P.; Yang, X.; Brestic, M. Does Silicon Really Matter for the Photosynthetic Machinery in Plants…? Plant Physiol. Biochem. 2021, 169, 40–48. [Google Scholar] [CrossRef]
- Etienne, P.; Trouverie, J.; Haddad, C.; Arkoun, M.; Yvin, J.C.; Caïus, J.; Brunaud, V.; Laîné, P. Root Silicon Treatment Modulates the Shoot Transcriptome in Brassica napus L. and in Particular Upregulates Genes Related to Ribosomes and Photosynthesis. Silicon 2021, 13, 4047–4055. [Google Scholar] [CrossRef]
- Holz, S.; Kube, M.; Bartoszewski, G.; Huettel, B.; Büttner, C. Initial Studies on Cucumber Transcriptome Analysis under Silicon Treatment. Silicon 2019, 11, 2365–2369. [Google Scholar] [CrossRef]
- Aucique-Pérez, C.E.; Rios, V.S.; Neto, L.B.C.; Rios, J.A.; Martins, S.C.V.; Rodrigues, F.A. Photosynthetic Changes in Wheat Cultivars with Contrasting Levels of Resistance to Blast. J. Phytopathol. 2020, 168, 721–729. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Sharif, Y.; Zafar, M.H.; Ali, H.; Khan, K.A. Role of Primary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2019, 137, 103728. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, P.; Jha, G. Alterations in Plant Sugar Metabolism: Signatory of Pathogen Attack. Planta 2019, 249, 305–318. [Google Scholar] [CrossRef]
- Van Bockhaven, J.; Steppe, K.; Bauweraerts, I.; Kikuchi, S.; Asano, T.; Höfte, M.; De Vleesschauwer, D. Primary Metabolism Plays a Central Role in Moulding Silicon-Inducible Brown Spot Resistance in Rice. Mol. Plant Pathol. 2015, 16, 811–824. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; Chaves, A.R.M.; Vale, F.X.R.; Damatta, F.M. Photosynthesis and Sugar Concentration Are Impaired by the Defective Active Silicon Uptake in Rice Plants Infected with Bipolaris oryzae. Plant Pathol. 2013, 62, 120–129. [Google Scholar] [CrossRef]
- Sun, S.; Yang, Z.; Song, Z.; Wang, N.; Guo, N.; Niu, J.; Liu, A.; Bai, B.; Ahammed, G.J.; Chen, S. Silicon Enhances Plant Resistance to Fusarium Wilt by Promoting Antioxidant Potential and Photosynthetic Capacity in Cucumber (Cucumis Sativus L.). Front. Plant Sci. 2022, 13, 1011859. [Google Scholar] [CrossRef]
- Peixoto, M.U.; Rios, J.A.; Silva, E.T.; Rodrigues, F.A. Silicon Alleviates Changes in the Source-Sink Relationship of Wheat Plants Infected by Pyricularia oryzae. Phytopathology 2019, 109, 1129–1140. [Google Scholar]
- Mayer, A.M.; Staples, R.C.; Gil-ad, N.L. Mechanisms of Survival of Necrotrophic Fungal Plant Pathogens in Hosts Expressing the Hypersensitive Response. Phytochemistry 2001, 58, 33–41. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Xue, G.; Cui, P.; Fan, F.; Liu, H.; Yin, C.; Sun, W.; Liang, Y. The Role of Silicon in Enhancing Resistance to Bacterial Blight of Hydroponic- and Soil-Cultured Rice. Sci. Rep. 2016, 6, 24640. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Tayyab, M.; Khalil, F.; Hua, Z.; Huang, Z.; Chen, H.Y.H. Silicon-Mediated Plant Defense against Pathogens and Insect Pests. Pestic. Biochem. Physiol. 2020, 168, 104641. [Google Scholar] [CrossRef] [PubMed]
- Sundaresha, S.; Kumar, A.M.; Rohini, S.; Math, S.A.; Keshamma, E.; Chandrashekar, S.C.; Udayakumar, M. Enhanced Protection against Two Major Fungal Pathogens of Groundnut, Cercospora arachidicola and Aspergillus flavus in Transgenic Groundnut over-Expressing a Tobacco β 1-3 Glucanase. Eur. J. Plant Pathol. 2010, 126, 497–508. [Google Scholar] [CrossRef]
- Fortunato, A.A.; Da Silva, W.L.; Rodrigues, F.A. Phenylpropanoid Pathway Is Potentiated by Silicon in the Roots of Banana Plants during the Infection Process of Fusarium Oxysporum f. sp. cubense. Phytopathology 2014, 104, 597–603. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Antisense Downregulation of Polyphenol Oxidase Results in Enhanced Disease Susceptibility. Planta 2004, 220, 105–117. [Google Scholar] [CrossRef]
- Rahman, A.; Wallis, C.M.; Uddin, W. Silicon-Induced Systemic Defense Responses in Perennial Ryegrass against Infection by Magnaporthe oryzae. Phytopathology 2015, 105, 748–757. [Google Scholar] [CrossRef]
- Viswanath, K.K.; Varakumar, P.; Pamuru, R.R.; Basha, S.J.; Mehta, S.; Rao, A.D. Plant Lipoxygenases and Their Role in Plant Physiology. J. Plant Biol. 2020, 63, 83–95. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Jeong, H.J.; Kim, D.H.; Shin, J.S.; Kim, J.G.; Yeon, M.H.; Lee, I.J. Regulation of Jasmonic Acid Biosynthesis by Silicon Application during Physical Injury to Oryza sativa L. J. Plant Res. 2014, 127, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Bhat, J.A.; Siddiqui, M.H.; Rinklebe, J.; Ahmad, P. Integration of Silicon and Secondary Metabolites in Plants: A Significant Association in Stress Tolerance. J. Exp. Bot. 2020, 71, 6758–6774. [Google Scholar] [CrossRef]
- Fagundes-Nacarath, I.R.F.; Debona, D.; Rodrigues, F.A. Oxalic Acid-Mediated Biochemical and Physiological Changes in the Common Bean-Sclerotinia sclerotiorum Interaction. Plant Physiol. Biochem. 2018, 129, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Korndörfer, G.; Pereira, H.; Nolla, A. Análise de Silício: Solo, Planta e Fertilizante; Universidade Federal de Uberlândia (UFU): Uberlândia, Brazil, 2004. [Google Scholar]
- Mesquita, G.L.; Tanaka, F.A.O.; Zambrosi, F.C.B.; Chapola, R.; Cursi, D.; Habermann, G.; Massola, N.S.; Ferreira, V.P.; Gaziola, S.A.; Azevedo, R.A. Foliar Application of Manganese Increases Sugarcane Resistance to Orange Rust. Plant Pathol. 2019, 68, 1296–1307. [Google Scholar] [CrossRef]
- Goldstein, J.; Newbury, D.; Joy, D.; Lyman, C.; Echlin, P.; Lifshin, E.; Sawyer, L.; Michael, J. Scanning Electron Microscopy and X-Ray Microanalysis; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003. [Google Scholar]
- Williams, J.S.; Hall, S.A.; Hawkesford, M.J.; Beale, M.H.; Cooper, R.M. Elemental Sulfur and Thiol Accumulation in Tomato and Defense against a Fungal Vascular Pathogen. Plant Physiol. 2002, 128, 150–159. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, History and Modelling. Ann. Bot. 2020, 126, 511–537. [Google Scholar] [CrossRef]
- Marçal, D.M.S.; Avila, R.T.; Quiroga-Rojas, L.F.; Souza, R.P.B.; Gomes Junior, C.C.; Ponte, L.R.; Barbosa, M.L.; Oliveira, L.A.; Martins, S.C.V.; Ramalho, J.D.C.; et al. Elevated [CO2] Benefits Coffee Growth and Photosynthetic Performance Regardless of Light Availability. Plant Physiol. Biochem. 2021, 158, 524–535. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Fernie, A.R. The Contribution of Plastidial Phosphoglucomutase to the Control of Starch Synthesis within the Potato Tuber. Planta 2001, 213, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.S.; Rios, J.A.; Einhardt, A.M.; Chaves, J.A.A.; Rodrigues, F.A. Effect of Glutamate on Pyricularia oryzae Infection of Rice Monitored by Changes in Photosynthetic Parameters and Antioxidant Metabolism. Physiol. Plant. 2020, 169, 179–193. [Google Scholar] [CrossRef]
- Chaves, J.A.A.; Oliveira, L.M.; Silva, L.C.; Silva, B.N.; Dias, C.S.; Rios, J.A.; Rodrigues, F.A. Physiological and Biochemical Responses of Tomato Plants to White Mold Affected by Manganese Phosphite. J. Phytopathol. 2021, 169, 149–167. [Google Scholar] [CrossRef]
- Fortunato, A.; Debona, D.; Bernardeli, A.; Rodrigues, F.A. Defence-Related Enzymes in Soybean Resistance to Target Spot. J. Phytopathol. 2015, 104, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Debona, D.; Rodrigues, F.A.; Rios, J.A.; Nascimento, K.J.T. Biochemical Changes in the Leaves of Wheat Plants Infected by Pyricularia oryzae. Phytopathology 2012, 102, 1121–1129. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables/Parameters * | Si | PI | Si × PI |
|---|---|---|---|
| Si | <0.001 | 0.244 | 0.901 |
| K | 0.185 | 0.222 | 0.634 |
| Sev | <0.001 | - | - |
| A | 0.0297 | <0.001 | 0.075 |
| gs | 0.093 | 0.709 | 0.502 |
| Ci | 0.653 | 0.083 | 0.749 |
| E | 0.117 | 0.159 | 0.122 |
| Fv/Fm | 0.002 | <0.001 | 0.004 |
| Y(II) | 0.008 | 0.258 | 0.377 |
| Y(NPQ) | 0.641 | 0.005 | 0.461 |
| Y(NO) | 0.813 | 0.718 | 0.799 |
| ETR | 0.003 | 0.06 | 0.095 |
| Chl a+b | 0.724 | <0.001 | 0.374 |
| Car | 0.894 | 0.008 | 0.593 |
| Glucose | 0.124 | <0.001 | 0.811 |
| Fructose | 0.229 | 0.205 | 0.485 |
| Sucrose | 0.067 | 0.001 | 0.028 |
| Starch | 0.345 | <0.001 | 0.585 |
| MDA | <0.001 | <0.001 | 0.001 |
| H2O2 | 0.293 | 0.082 | 0.198 |
| O2•− | 0.001 | <0.001 | 0.001 |
| TSP | 0.783 | 0.039 | 0.375 |
| LTGA derivatives | 0.005 | <0.001 | <0.001 |
| CHI | 0.363 | 0.015 | 0.203 |
| GLU | 0.958 | 0.008 | 0.941 |
| PAL | 0.040 | <0.001 | 0.166 |
| PPO | 0.463 | <0.001 | 0.944 |
| POX | 0.569 | <0.001 | 0.476 |
| LOX | 0.915 | <0.001 | 0.845 |
| SOD | 0.110 | <0.001 | 0.062 |
| APX | 0.128 | <0.001 | 0.053 |
| CAT | 0.405 | <0.001 | 0.568 |
| GR | 0.952 | 0.492 | 0.908 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lata-Tenesaca, L.F.; Oliveira, M.J.B.; Barros, A.V.; Picanço, B.B.M.; Rodrigues, F.Á. Physiological and Biochemical Aspects of Silicon-Mediated Resistance in Maize against Maydis Leaf Blight. Plants 2024, 13, 531. https://doi.org/10.3390/plants13040531
Lata-Tenesaca LF, Oliveira MJB, Barros AV, Picanço BBM, Rodrigues FÁ. Physiological and Biochemical Aspects of Silicon-Mediated Resistance in Maize against Maydis Leaf Blight. Plants. 2024; 13(4):531. https://doi.org/10.3390/plants13040531
Chicago/Turabian StyleLata-Tenesaca, Luis Felipe, Marcos José Barbosa Oliveira, Aline Vieira Barros, Bárbara Bezerra Menezes Picanço, and Fabrício Ávila Rodrigues. 2024. "Physiological and Biochemical Aspects of Silicon-Mediated Resistance in Maize against Maydis Leaf Blight" Plants 13, no. 4: 531. https://doi.org/10.3390/plants13040531