1. Introduction
Cannabis sativa (hereafter, cannabis) is a dioecious annual that has been domesticated worldwide for medicinal, textile, recreational, and nutritional uses. Cannabis is well known for the cannabinoids it produces such as tetrahydrocannabinol (THC) and cannabidiol (CBD), which are produced in high abundance in stalked glandular trichomes associated with unfertilized floral tissues of female plants (Small, 2016) [
1].
Most cannabis cultivars that are grown commercially in indoor environments are drug-type (i.e., THC content > 0.3%), due to the relatively high value of their mature female floral tissues. Indoor-grown cannabis cultivars typically have short-day photoperiod responses, whereby daylengths greater than 16 h (i.e., uninterrupted dark periods ≤ 8 h) will keep plants in the vegetative growth stage while shortening the photoperiod below some critical level will transition plants toward producing generative tissues (i.e., flowering initiation).
The robustness of photoperiodic cannabis’ flowering response tends to increase as the daylength continues to be reduced below the critical photoperiod. Many modern drug-type cannabis cultivars have been developed by hybridizing cannabis cultivars grown from a broad range of latitudes with different seasonal photoperiod dynamics (Small, 2022) [
2]. Therefore, modern cannabis cultivars may have considerable variability in their optimum daylengths for promoting robust flowering responses and maximizing yield and quality. The range of photoperiodic responses has been well documented in hemp cultivars (e.g., Zhang et al., 2021) [
3], but the studies by Peterswald et al. (2023) [
4] and Ahrens et al. (2023) [
5] are the only recent studies on photoperiodic responses of modern indoor-grown cannabis cultivars. Indoor-grown cannabis crops are almost universally switched to a 12-h daily photoperiod to initiate the flowering stage of production (Potter, 2014) [
6]. This optimizes operational simplicity in cultivation systems that grow many different cultivars concurrently. However, recent studies have shown that some modern indoor-grown cannabis cultivars have robust flowering responses to moderately longer photoperiods (Peterswald et al., 2023; Ahrens et al., 2023; Potter, 2009) [
4,
5,
7]. Longer photoperiods have the advantages of either (1) increased daily light integrals (DLIs), which may result in higher yields (Rodriguez-Morrison et al., 2020) [
8], or (2) lower installed lighting levels (e.g., reduced fixture density), which reduces upfront lighting infrastructure and installation costs.
Our recent study demonstrated that about half of the ten investigated cultivars had minimal delays in flowering initiation in photoperiods up to 13.5 h (Ahrens et al., 2023) [
5]. However, this study ended well before the cultivars reached normal commercial maturity. Therefore, the effects of longer photoperiods on mature inflorescence yield and quality are still unknown. The objective of the present study was to compare the yield and quality of two cultivars (selected from the study by Ahrens et al. (2023) [
5]) grown to commercial maturity under 12 h or 13 h photoperiods. The hypotheses were that inflorescence yield would be proportional to the DLI associated with each photoperiod and that there would be no photoperiod treatment effects on the concentrations of the major cannabinoids.
2. Materials and Methods
2.1. Experimental Design and Plant Cultivation
This trial was conducted in a walk-in growth chamber at the University of Guelph with two rows of three compartments separated by opaque white curtains to prevent inter-compartment light contamination, as described in the study by Ahrens et al. (2023) [
5]. Two full spectrum LED fixtures (Jungle—LED G4i 1200, Allstate Garden Supply, Ontario, CA, USA) were hung in each compartment on height-adjustable pulley systems. These fixtures have onboard dimmers with fixed settings of 25%, 50%, 75%, and 100% of maximum intensity. The relative spectral photon flux distribution of the LED fixtures is provided in
Figure S1, which was the same as in the study by Ahrens et al. (2023) [
5].
Two cannabis cultivars, ‘Incredible Milk’ (IM) and ‘Gorilla Glue’ (GG), were sourced from a single commercial cultivator in Southwestern Ontario. Seventy-eight stem tip cuttings were taken from vegetative mother plants of each cultivar and inserted into 3.6 × 4.0 cm round rockwool plugs (Macroplug; Grodan, Milton, ON, Canada) on 7 November 2022 at the cultivator’s facility. The rooted cuttings were delivered to the University of Guelph on 29 November 2022 and transplanted on 1 December 2022 into presoaked 10 × 10 × 7.5 cm rockwool blocks (Grodan GRO-BLOCK Improved GR7.5 Medium 4”; Grodan). Transplants were grown vegetatively (18 h light/6 h dark) with the LEDs at 85 cm above the canopy. The dimmers were set at 25% on 29 November 2022, which provided an average canopy-level photosynthetic photon flux density (PPFD) of ≈135 µmol·m−2·s−1. Dimmer settings were incrementally increased every two days until reaching 100% (≈540 µmol·m−2·s−1) on 5 December 2022.
Eighteen uniformly sized plants of each cultivar were randomly assigned to one of two photoperiod treatments on 5 December 2022: 12 h light/12 h dark (12 h) or 13 h light/11 h dark (13 h). All LED fixtures were set to maximum intensity, resulting in uniform PPFD of ≈540 µmol·m
−2·s
−1 at an 85 cm hang height. The daily light integrals for the 12 h and 13 h photoperiods were 23.8 and 25.7 mol·m
−2·d
−1, respectively. The photoperiod treatments were randomly assigned to individual compartments, resulting in 3 concurrent replications of each treatment. Three plants of each cultivar were placed onto presoaked 100 × 15 × 7.5 cm rockwool slabs (Vital; Grodan), one slab per cultivar, in each compartment. The locations of each cultivar’s slab were randomized in each compartment (
Figure S2). Each slab was centered in a subirrigation tray (53 × 109 cm grow trays, Botanicare, Vancouver, WA, USA). The plants were 35 cm apart on the slab and 70 cm apart between slabs measured ‘on center’ (
Figure S3). The plants were subirrigated as needed for the first 9 d after the photoperiod treatments began and then drip-irrigated henceforth. One irrigation dripper (Supertif PCND-MOP 1.1 L·h
−1; Rivulis, Gvat, Israel) was placed in each rockwool block, and two drippers were placed in each rockwool slab, centered between adjacent plants (
Figure S3). There were eight irrigation events per day, with event lengths increased, as needed, to maintain a daily leachate percentage of at least 15%. The first irrigation event occurred ≤ 1 h after lights turned on, followed by 1 h rest periods between the next five irrigations and 2 h rest periods between the last two irrigations. The length of individual irrigations was the same throughout any given day but increased from 120 s at the beginning of the trial to 420 s by the end, to maintain ≥ 15% daily leachate levels. The cannabis flowering nutrient recipe from Zheng (2022) [
9] was used, mixed in deionized water to an electrical conductivity of 2.0 mS·cm
−1, and pH was adjusted to 5.7 with a 1.0 M potassium bicarbonate (Master Plant-Prod Inc., Brampton, ON, Canada).
The chamber temperature was set at a constant 25 °C, and relative humidity (RH) was set at 70% light/60% dark, which was controlled with a fogging system connected to the climate computer (Titan I/O Plus, Argus Control Systems Limited, Surrey, BC, Canada). The LEDs were turned on daily at 09:00 and turned off at 21:00 and 22:00 for the 12-h and 13-h treatments, respectively. The photoperiod was controlled with the computer. The curtains at the front of each compartment were opened daily at 09:00 and closed daily at 21:00. The LED fixtures were raised weekly to maintain the 85 cm distance between the canopy and the LEDs. There was no CO2 supplementation in this trial, but ample air exchanges ensured CO2 concentrations were consistently >400 PPM during lit periods.
2.2. Data Collection
Temperature and relative humidity data were logged every 300 s using three dataloggers (MX2301A; HOBO; Onset Computer Corporation; Bourne, MA, USA), with each logger assigned to one replicate compartment from each treatment. The loggers were switched between their respective compartments daily. The day and night aerial temperature and RH for each compartment are summarized in
Table S1.
Elapsed days to flowering (EDTF), which is defined as the number of days between invoking the photoperiod treatments and the appearance of ≥3 pairs of stigmas (i.e., an inflorescence) at the top of the primary shoot, was monitored daily starting at 1 d after the photoperiod treatments began. The apical inflorescence was defined as the conglomeration of inflorescence and foliar tissues (i.e., sugar leaves) with no visible gaps located at the top of the primary shoot. The percentage of brown stigmas and amber trichomes on the apical inflorescence were monitored daily to track inflorescence maturity, beginning 37 d after photoperiod treatments began.
The harvesting of each cultivar began after >95% of the stigmas in either photoperiod treatment had turned brown, and either 10% of the trichomes had turned to an amber color (IM) or the fan leaves began senescing (GG), whichever occurred first. The harvesting of IM and GG began on days 58 and 72, respectively, after initiating the photoperiod treatments. One replicate of each treatment (i.e., six plants) was harvested per day, for three successive days.
The harvest process followed the study by Ahrens et al. (2023) [
5] with the addition of separating the vegetative tissues into fan leaves, sugar leaves, and stems. To differentiate between fan and sugar leaves, any leaf with ≥1 cm of petiole visible when the longitudinal axis of the plant was at the nadir with respect to the observer (i.e., looking straight downwards from the top of the plant) was considered a fan leaf. Prior to cutting the plant at the substrate surface (i.e., the top of the rockwool blocks), plant height and width (at the widest point and perpendicular to this) were measured to determine the growth index following the study by Ruter (1992) [
10]. The apical inflorescences were separated from the rest of the plants to determine their volume (following Ahrens et al., 2023 [
5]) and fresh weight (FW). The apical inflorescence FW was divided by volume to determine apical inflorescence density (g·cm
−3). The apical inflorescence of the most representative plant from each photoperiod by cultivar combination in each replicate was air-dried at 23 °C and 70% RH to approximately 10% moisture content. During the harvesting process, photographs were taken of the whole plant, the whole plant with fan leaves removed, and the excised apical inflorescence of the most representative plant in each photoperiod by cultivar combination in each replicate. The FW of the fan leaves, sugar leaves, stems, and the nonapical inflorescence tissues of each plant were recorded. The harvest index was calculated as follows: total inflorescence FW/(total inflorescence FW + aboveground vegetative FW). The separated tissues of each photographed plant were dried to constant weight at 70 °C. The representative plants’ tissue dry weights (DWs) were then used to calculate the moisture content of each tissue type, which were then used to estimate the DWs of the remaining plants in each respective cultivar by photoperiod combination. All tissue weights were measured in grams using a digital balance (BCE2202-1S; Sartorius Lab Instruments, Göttingen, Germany).
2.3. Cannabinoid Analysis
The air-dried apical inflorescences of all of the representative plants were sent to a third-party lab (A&L Canada Laboratories Inc., London, ON, Canada) for the analysis of moisture content (i.e., loss on drying) and the concentrations of the following cannabinoids: cannabigerolic acid (CBGA), cannabigerol (CBG), cannabidiolic acid (CBDA), cannabidiol (CBD), cannabinolic acid (CBNA), cannabinol (CBN), cannabidivarinic acid (CBDVA), cannabidivarin (CBDV), Δ9-tetrahydrocannabinolic acid (THCA), Δ9-tetrahydrocannabinol (THC), tetrahydrocannabivarinic acid (THCVA), and tetrahydrocannabivarin (THCV). The total equivalent THC (T-THC) is an estimation of the amount of THC available to the consumer, once THCA has been decarboxylated into THC, such as from adding heat. The equation used is T-THC = (THCA × 0.877) + THC. The limit of quantitation (i.e., LOQ) for these analyses was reported as <0.05% for each cannabinoid. All cannabinoid concentrations are reported as the percentage of dry weight, based on the loss-on-drying determination of the sampled tissues’ moisture content. The total inflorescence dry weight was calculated by adding nonapical inflorescence dry weight (oven-dried) and apical inflorescence dry weight (loss on drying) for each plant.
2.4. Statistical Analysis
The experiment was a completely randomized design, with two treatments and three replications. Statistical analysis was performed using RStudio (v2021.9.0.351; Posit Software, PBC; Boston, MA, USA). The parameter means of each replicate in each cultivar by treatment combination were analyzed for the EDTF, harvest, and postharvest measurements (n = 3). Each sample was considered a replicate in the cannabinoid data (n = 3). Unpaired, two-tailed Student’s t-tests were performed between the 12 h and 13 h treatments for each parameter by cultivar combination. Statistical significance was determined at the p ≤ 0.05 level.
4. Discussion
An almost universal practice in indoor cannabis cultivation is to reduce the daily photoperiod to 12 h to transition vegetative plants to flowering. While this protocol works for virtually all indoor-grown cannabis cultivars, some cultivars have robust flowering responses to photoperiods that are modestly longer than 12 h (Peterswald et al., 2023; Ahrens et al., 2023) [
4,
5]. Since higher light levels can increase inflorescence yield and quality (Rodriguez-Morrison et al., 2021; Llewellyn et al., 2022) [
8,
11], longer flowering-stage photoperiods (with corresponding increases in DLIs) may have positive influences on cannabis inflorescence yield and quality. Also, in greenhouse production during summer months in higher latitude regions (e.g., Canada), blackout curtains can remain open longer, allowing for better utilization of natural light and reductions in some other climate-control-related costs. A 12 h flowering-stage photoperiod may not be optimized for maximizing the yield of all cultivars. Hence, cultivators who use a 12 h photoperiod for all cultivars may be ‘leaving yield on the floor’. The ≥35% increases in the total inflorescence yield in the 13 h treatment observed in the present study were similar to the yield increases in the 14 h vs. 12 h photoperiod reported by Peterswald et al. (2023) [
4], who attributed the higher yields to increased DLIs. However, in both the present study and that of Peterswald et al. (2023) [
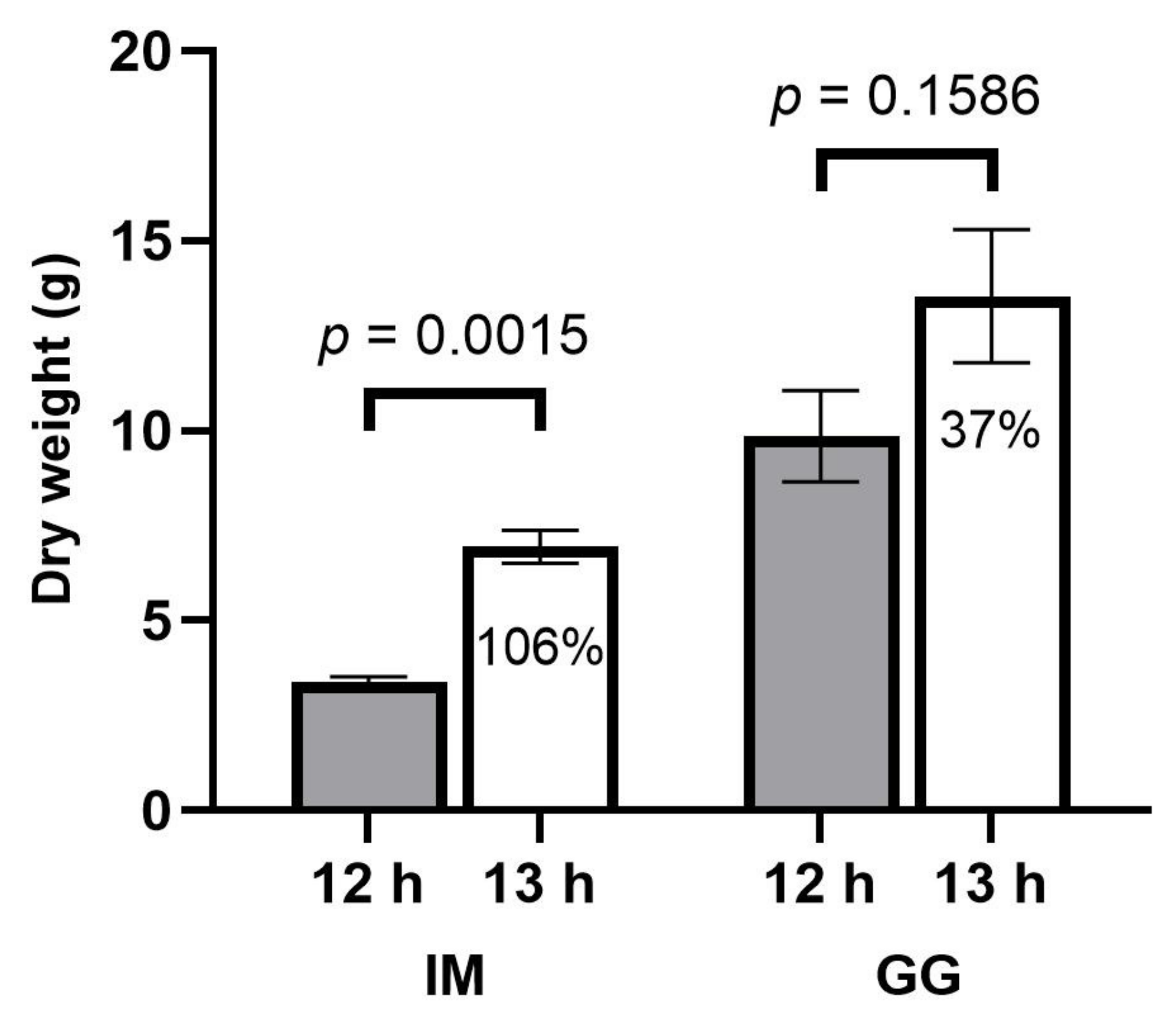
4], the observed yield increases under longer photoperiods were disproportionately higher than the increases in DLIs, contrary to our hypothesis. Other developmental and morphophysiological responses to longer photoperiods may be contributing to the yield enhancements.
In outdoor environments, after the summer solstice (i.e., 21 June in the Northern Hemisphere), daily photoperiods gradually reduce a few minutes each day, with daily changes in photoperiod depending on latitude and time of year. For example, in Guelph, Ontario, Canada (43.5° N, −80.2° W), there are ≈90 d between the summer solstice (15.5 h) and 12 h days (Sept 26) (Hoffmann, 2024) [
12]. For short-day plants, the seasonal reductions in daylength indicate the onset of the end of the growing season; photoperiodic cannabis cultivars respond accordingly by transitioning from vegetative to generative growth, ending in floral maturation, senescence, and plant death. In contrast to the outdoor environment, reducing from ≥16 h directly to 12 h in indoor cannabis cultivation is a relatively powerful signal that the end of the growing season (e.g., the onset of killing frost) approaches. Accordingly, regardless of their provenance, virtually all indoor-grown cannabis cultivars rapidly initiate flowering under a 12 h photoperiod, normally producing visible inflorescence tissues (groupings of ≥3 stigmas) in less than two weeks and mature inflorescences six to ten weeks later (Peterswald et al., 2023; Potter, 2014; Rodriguez-Morrison et al., 2021; Llewellyn et al., 2022; Potter and Duncombe, 2012) [
4,
6,
8,
11,
13].
While some indoor-grown cannabis cultivars can flower under photoperiods longer than 12 h, increasing the photoperiod may moderate the speed and intensity of the transition to reproductive growth. This is supported in the present study by the delayed time to visible inflorescences in IM and slower early floral development in both cultivars. These patterns were also reported by Ahrens et al. (2023) [
5] for some cultivars, and by Zhang et al. (2021) for essential oil cultivars [
3]. However, as seen in the present study, delayed or repressed flowering initiation in the 13 h photoperiod led to enhanced rather than repressed inflorescence biomass when plants reached commercial maturity. Since much of the vegetative growth after switching to the flowering photoperiod occurs during the first few weeks after the transition to short days (Potter, 2014; Yep et al., 2020) [
6,
14], enhancements in vegetative growth during this period increase foliar biomass, probably enhancing light interception and thus growth potential. Enhanced vegetative growth during the early phases of the flowering stage may also increase the number of potential flowering sites and capacity for structurally supporting higher floral biomass, which has been observed in vegetative-stage cannabis (Moher et al., 2022) [
15] and in flowering hemp grown in protected culture (Hall et al., 2014) [
16]. The plants of both cultivars in the 13 h treatment in the present study had higher growth indexes and appeared to be larger. However, increases in the DW of individual vegetative tissues in the 13 h treatment were only observed in IM. Regardless, the higher inflorescence yields in the 13 h treatment may be a consequence of generally larger plants, which are known to increase commercially mature inflorescence yield potential (Bevan et al., 2021) [
17].
Despite the early delays in inflorescence development, by the time the plants in the 12 h treatment reached commercial maturity, the total inflorescence yield and the size of the apical inflorescences were markedly higher in the 13 h treatment in both cultivars. Given that the increases in inflorescence yield were disproportionately higher than the increase in DLI, flowering photoperiod management may be one of the most efficacious cultural practices available to commercial indoor cannabis cultivators for increasing yield that is both simple and cost-effective to utilize. Furthermore, despite the plants in the 13 h treatment appearing to have delayed inflorescence maturation rates (e.g., slower stigma browning and reduced trichome ambering), the cannabinoid composition in the apical inflorescences was of comparable quality in both treatments in both cultivars. There were moderately higher CBGA concentrations in the 13 h treatment in IM, which suggests that the inflorescence tissues in this treatment may have been less mature (Aizpurua-Olaizola et al., 2016) [
18]. However, there was no detectable CBN in either cultivar, and the neutral form of THC was a low proportion of total THC, suggesting that neither treatment had been harvested beyond its optimum maturity level (Aizpurua-Olaizola et al., 2016; Russo, 2007) [
18,
19]. The CBDA levels in IM were also higher in the 13 h treatment. However, as the THCA-to-CBDA ratios were ≥400 in both cultivars, treatment effects in CBD content in the cultivars used in this study are likely not commercially relevant, at least in most markets that favor such highly THC-dominant cultivars. This is also the case with the observed treatment effects on THC concentrations, which represented only a small fraction of the total THC (T-THC) content. More relevant are the effects of photoperiod on the T-THC content, which were either insignificant (GG) or higher (IM) in the 13 h treatment. Since the 13 h treatment increased the inflorescence DW, the 13 h treatment also increased THC yield (g/plant) accordingly, by ≥38%. These results are in contrast with those obtained by Peterswald et al. (2023) [
4], who found approximately 40% reductions in the floral THC concentrations in the two THC-dominant cultivars when grown in the 14 h vs. 12 h photoperiod treatment. These results are also contrary to our hypothesis, which distinguishes the present study from many contemporary cannabis lighting studies that often showed only minor effects of light treatments on cannabinoid composition (e.g., Rodriguez-Morrison et al., 2021; Llewellyn et al., 2022; Potter and Duncombe, 2012) [
8,
11,
13]. Further study is needed to determine the mechanisms for how photoperiod, along with other environmental inputs, affect the composition of the metabolome in indoor-grown cannabis.
Another important quality aspect of cannabis is the density of the apical inflorescences, where more dense inflorescences are normally regarded as having higher quality. Apical inflorescence density has been shown to increase linearly with light intensity (Rodriguez-Morrison et al., 2021) [
8]. However, inflorescence density in IM was lower in the longer photoperiod in the current study, suggesting that the developmental ramifications of longer photoperiods on apical inflorescence tissues may override the benefits of higher DLIs. Overall, aside from lower apical inflorescence density in IM, the 13 h treatment substantially increased the apical inflorescence size, total inflorescence yield, and cannabinoid yield.
Despite having similar prescribed days to maturity in commercial production (Ahrens et al., 2023) [
5], GG required ≈25% longer to reach commercial maturity in the present study, regardless of the photoperiod treatment. Factoring in the relative lengths of the flowering cycle of each cultivar, IM was ≈25% more efficient (i.e., g·d
−1) than GG at producing floral biomass in both treatments. While further study is required to elucidate the specific mechanisms of photoperiod responses of indoor-grown cannabis cultivars, the observed cultivar differences in photoperiod responses generally illustrate the need to investigate individual cultivars’ photoperiod responses under the cultivators’ specific cultivation environment for full flowering cycles.
Due to the relatively small plot sizes in the present study, the massive per-plant yield increases may be tempered by competition for space and light in commercial indoor cannabis cultivation systems, depending on the planting density. Readers are therefore cautioned against inferring per-area yield increases (e.g., over commercially relevant production areas) from the per-plant yield increases reported in the present study. Future research should explore the effects of moderately longer photoperiods than 12 h on the inflorescence yield and quality of different indoor-grown cannabis cultivars. Particular focus should be directed toward investigating narrower discreet time differences between photoperiod treatments (e.g., ≤15 min) and the effects of planting density on the treatment effects of photoperiod on yield and quality, both on per-plant and per-area bases.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}