
Metabolomic Response of Thalassiosira weissflogii to Erythromycin Stress: Detoxification Systems, Steroidal Metabolites, and Energy Metabolism
Abstract
:1. Introduction
2. Results
2.1. Erythromycin Exposure Modified the Growth of Marine Microalga
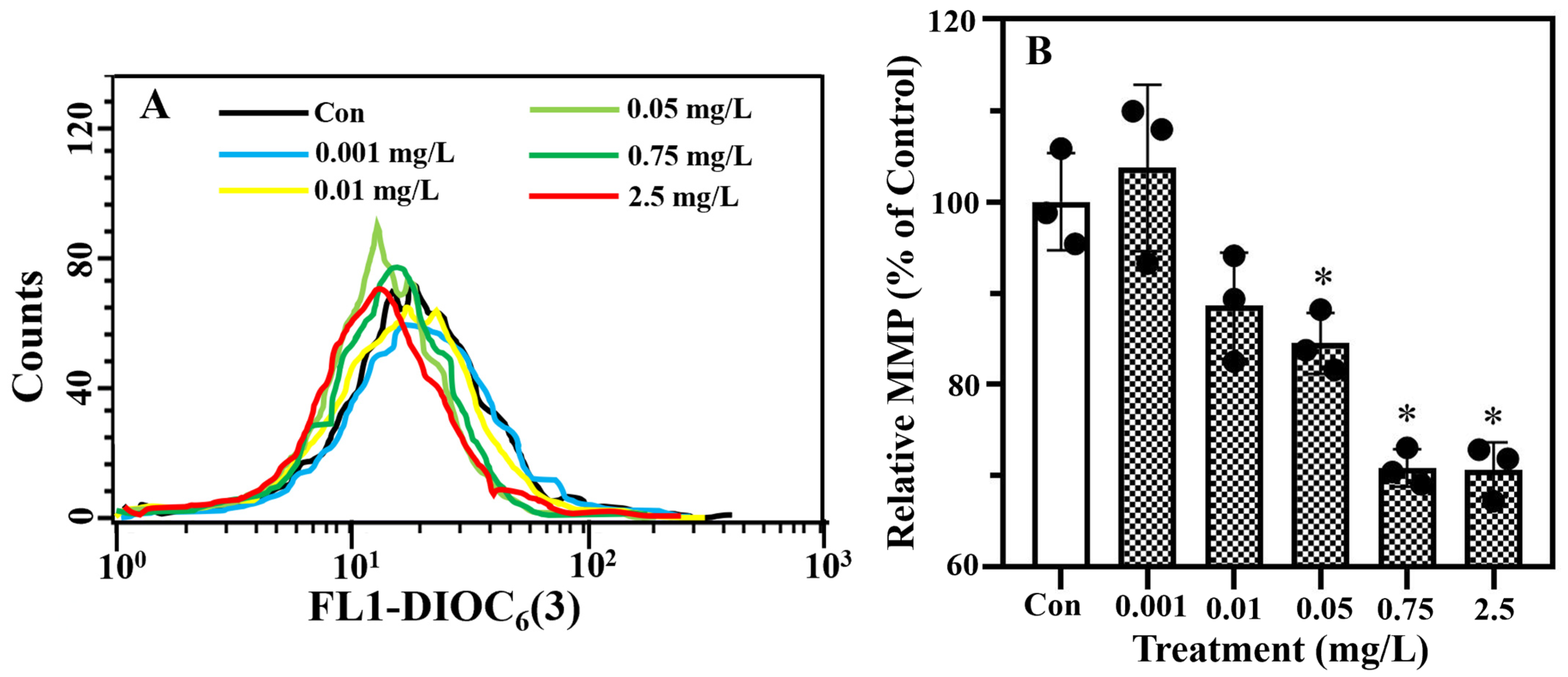
2.2. Erythromycin Exposure Modified MMP and Caused Algal Cell Death
2.3. Erythromycin Exposure Altered Metabolomic Profiles
2.4. Conjoint Analysis of Metabolomic and Transcriptomic Data
3. Discussion
3.1. Detoxification Systems
3.2. Energy Metabolism
3.3. Thiamine Metabolism and Metabolism of Porphyrin and Chlorophyll
3.4. Environmental Implications
4. Materials and Methods
4.1. Algal Cultivation and Toxicity Testing
4.2. Determination of Biochemical Parameters
4.3. Metabolomic Profiling
4.4. Conjoint Analysis of Metabolome and Transcriptome
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: An analysis of national pharmaceutical sales data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Danner, M.C.; Robertson, A.; Behrends, V.; Reiss, J. Antibiotic pollution in surface fresh waters: Occurrence and effects. Sci. Total. Environ. 2019, 664, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Daughton, C.G.; Ternes, T.A. Pharmaceuticals and personal care products in the environment: Agents of subtle change? Environ. Health Perspect. 1999, 107 (Suppl. 6), 907–938. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wu, W.; Zhang, J.; Lv, P.; Xu, L.; Yan, Y. Progress of research on the toxicology of antibiotic pollution in aquatic organisms. Acta Ecol. Sin. 2018, 38, 36–41. [Google Scholar] [CrossRef]
- Li, J.; Li, W.; Liu, K.; Guo, Y.; Ding, C.; Han, J.; Li, P. Global review of macrolide antibiotics in the aquatic environment: Sources, occurrence, fate, ecotoxicity, and risk assessment. J. Hazard. Mater. 2022, 439, 129628. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Guo, J.; Iwata, H.; Diamond, J.; Qu, C.; Xiong, J.; Han, J. What approaches should be used to prioritize pharmaceuticals and personal care products for research on environmental and human health exposure and effects? Environ. Toxicol. Chem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Liu, B.; Liu, W.; Nie, X.; Guan, C.; Yang, Y.; Wang, Z.; Liao, W. Growth response and toxic effects of three antibiotics on Selenastrum capricornutum evaluated by photosynthetic rate and chlorophyll biosynthesis. J. Environ. Sci. 2011, 23, 1558–1563. [Google Scholar] [CrossRef]
- Välitalo, P.; Kruglova, A.; Mikola, A.; Vahala, R. Toxicological impacts of antibiotics on aquatic micro-organisms: A minireview. Int. J. Hyg. Environ. Health 2017, 220, 558–569. [Google Scholar] [CrossRef]
- Ashraf, A.; Liu, G.; Yousaf, B.; Arif, M.; Ahmed, R.; Irshad, S.; Gulzaman, H. Recent trends in advanced oxidation process-based degradation of erythromycin: Pollution status, ecotoxicity and degradation mechanism in aquatic ecosystems. Sci. Total. Environ. 2021, 772, 145389. [Google Scholar] [CrossRef]
- Lin, A.Y.; Tsai, Y.T. Occurrence of pharmaceuticals in Taiwan’s surface waters: Impact of waste streams from hospitals and pharmaceutical production facilities. Sci. Total Environ. 2009, 407, 3793–3802. [Google Scholar] [CrossRef] [PubMed]
- Alighardashi, A.; Pandolfi, D.; Potier, O.; Pons, M.N. Acute sensitivity of activated sludge bacteria to erythromycin. J. Hazard. Mater. 2009, 172, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Campa-Córdova, A.I.; Luna-Gonzalez, A.; Ascencio, F.; Cortés-Jacinto, E.; Cáceres-Martínez, C.J. Effects of chloramphenicol, erythromycin, and furazolidone on growth of Isochrysis galbana and Chaetoceros gracilis. Aquaculture 2006, 260, 145–150. [Google Scholar] [CrossRef]
- Guo, J.; Ma, Z.; Peng, J.; Mo, J.; Li, Q.; Guo, J.; Yang, F. Transcriptomic analysis of Raphidocelis subcapitata exposed to erythromycin: The role of DNA replication in hormesis and growth inhibition. J. Hazard. Mater. 2021, 402, 123512. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Ma, Z.; Yan, S.; Cheung, N.K.; Yang, F.; Yao, X.; Guo, J. Metabolomic profiles in a green alga (Raphidocelis subcapitata) following erythromycin treatment: ABC transporters and energy metabolism. J. Environ. Sci. 2023, 124, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.P.; Liu, B.Y.; Yu, H.J.; Liu, W.Q.; Yang, Y.F. Toxic effects of erythromycin, ciprofloxacin and sulfamethoxazole exposure to the antioxidant system in Pseudokirchneriella subcapitata. Environ. Pollut. 2013, 172, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Kang, Y.H.; Lee, C.H.; Chung, I.K. Effects of six antibiotics on the activity of the photosynthetic apparatus and ammonium uptake of thallus of Porphyra yezoensis. Algae 2005, 20, 121–125. [Google Scholar] [CrossRef]
- Sendra, M.; Moreno-Garrido, I.; Blasco, J.; Araújo, C.V. Effect of erythromycin and modulating effect of CeO2 NPs on the toxicity exerted by the antibiotic on the microalgae Chlamydomonas reinhardtii and Phaeodactylum tricornutum. Environ. Pollut. 2018, 242, 357–366. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, Q.; Li, J.; Chen, X.; Lang, Q.; Kuang, S. Combined effects of erythromycin and enrofloxacin on antioxidant enzymes and photosynthesis-related gene transcription in Chlorella vulgaris. Aquat. Toxicol. 2019, 212, 138–145. [Google Scholar] [CrossRef]
- Zhang, Y.; Calabrese, E.J.; Zhang, J.; Gao, D.; Qin, M.; Lin, Z. A trigger mechanism of herbicides to phytoplankton blooms: From the standpoint of hormesis involving cytochrome b559, reactive oxygen species and nitric oxide. Water Res. 2020, 173, 115584. [Google Scholar] [CrossRef]
- Deng, C.N.; Zhang, D.Y.; Pan, X.L. Toxic effects of erythromycin on photosystem I and II in Microcystis aeruginosa. Photosynthetica 2014, 52, 574–580. [Google Scholar] [CrossRef]
- Machado, M.D.; Soares, E.V. Impact of erythromycin on a non-target organism: Cellular effects on the freshwater microalga Pseudokirchneriella subcapitata. Aquat. Toxicol. 2019, 208, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Guo, P.; Peng, X.; Wen, K. Effect of erythromycin exposure on the growth, antioxidant system and photosynthesis of Microcystis flos-aquae. J. Hazard. Mater. 2015, 283, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zeng, Y.; Qu, J.Y.; Wang, W.X. Mercury effects on Thalassiosira weissflogii: Applications of two-photon excitation chlorophyll fluorescence lifetime imaging and flow cytometry. Aquat. Toxicol. 2012, 110, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wei, D.; Xie, J. Diatoms as cell factories for high-value products: Chrysolaminarin, eicosapentaenoic acid, and fucoxanthin. Crit. Rev. Biotechnol. 2020, 40, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Marella, T.K.; Bhattacharjya, R.; Tiwari, A. Impact of organic carbon acquisition on growth and functional biomolecule production in diatoms. Microb. Cell Factories 2021, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, Y.; Li, Z.; Xu, J.; Gao, K. Effects of seawater acidification on the growth rates of the diatom Thalassiosira (Conticribra) weissflogii under different nutrient, light, and UV radiation regimes. J. Appl. Phycol. 2017, 29, 133–142. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiragushi, N.; Kitaguchi, H.; Mitsutani, A.; Nagai, S.; Yoshimura, M. A highly CO2-tolerant diatom, Thalassiosira weissflogii H1, enriched from coastal sea, and its fatty acid composition. Fish. Sci. 2000, 66, 655–659. [Google Scholar] [CrossRef]
- Filimonova, V.; Nys, C.; De Schamphelaere, K.A.; Gonçalves, F.; Marques, J.C.; Gonçalves, A.M.; De Troch, M. Ecotoxicological and biochemical mixture effects of an herbicide and a metal at the marine primary producer diatom Thalassiosira weissflogii and the primary consumer copepod Acartia tonsa. Environ. Sci. Pollut. Res. 2018, 25, 22180–22195. [Google Scholar] [CrossRef]
- Bielmyer-Fraser, G.K.; Jarvis, T.A.; Lenihan, H.S.; Miller, R.J. Cellular partitioning of nanoparticulate versus dissolved metals in marine phytoplankton. Environ. Sci. Technol. 2014, 48, 13443–13450. [Google Scholar] [CrossRef]
- Jarvis, T.A.; Miller, R.J.; Lenihan, H.S.; Bielmyer, G.K. Toxicity of ZnO nanoparticles to the copepod Acartia tonsa, exposed through a phytoplankton diet. Environ. Toxicol. Chem. 2013, 32, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, A.F.; Gonçalves, F.J.; Rocha, C.P.; Marques, J.C.; Gonçalves, A.M. Biochemical effects of two pesticides in three different temperature scenarios on the diatom Thalassiosira weissflogii. Processes 2021, 9, 1247. [Google Scholar] [CrossRef]
- Reinfelder, J.R.; Jablonka, R.E.; Cheney, M. Metabolic responses to subacute toxicity of trace metals in a marine microalga (Thalassiosira weissflogii) measured by calorespirometry. Environ. Toxicol. Chem. 2000, 19, 448–453. [Google Scholar]
- Mo, J.; Lv, R.; Qin, X.; Wu, X.; Chen, H.; Yan, N.; Shi, J.; Wu, Y.; Liu, W.; Kong, R.Y.C.; et al. Mechanistic insights into hormesis induced by erythromycin in the marine alga Thalassiosira weissflogii. Ecotoxicol. Environ. Saf. 2023, 263, 115242. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Hormesis: Highly generalizable and beyond laboratory. Trends Plant Sci. 2020, 25, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Steinman, A.D.; Xue, Q.; Zhao, Y.; Xu, Y.; Xie, L. Effects of erythromycin and sulfamethoxazole on Microcystis aeruginosa: Cytotoxic endpoints, production and release of microcystin-LR. J. Hazard. Mater. 2020, 399, 123021. [Google Scholar] [CrossRef] [PubMed]
- Labine, L.M.; Simpson, M.J. The use of nuclear magnetic resonance (NMR) and mass spectrometry (MS)–based metabolomics in environmental exposure assessment. Curr. Opin. Environ. Sci. Health 2020, 15, 7–15. [Google Scholar] [CrossRef]
- Carmen, B. Metabolomics in environmental toxicology: Applications and challenges. Trends Environ. Anal. Chem. 2022, 34, e00161. [Google Scholar]
- Bártíková, H.; Skálová, L.; Stuchlíková, L.; Vokřál, I.; Vaněk, T.; Podlipná, R. Xenobiotic-metabolizing enzymes in plants and their role in uptake and biotransformation of veterinary drugs in the environment. Drug Metab. Rev. 2015, 47, 374–387. [Google Scholar]
- Ramel, F.; Sulmon, C.; Serra, A.A.; Gouesbet, G.; Couée, I. Xenobiotic sensing and signalling in higher plants. J. Exp. Bot. 2012, 63, 3999–4014. [Google Scholar] [CrossRef]
- Shoji, T. ATP-binding cassette and multidrug and toxic compound extrusion transporters in plants: A common theme among diverse detoxification mechanisms. Int. Rev. Cell. Mol. Biol. 2014, 309, 303–346. [Google Scholar] [PubMed]
- Do, T.H.T.; Martinoia, E.; Lee, Y. Functions of ABC transporters in plant growth and development. Curr. Opin. Plant Biol. 2018, 41, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Fernie, A.R.; Nunes-Nesi, A. The central carbon and energy metabolism of marine diatoms. Metabolites 2013, 3, 325–346. [Google Scholar] [CrossRef] [PubMed]
- Atteia, A.; van Lis, R.; Tielens, A.G.; Martin, W.F. Anaerobic energy metabolism in unicellular photosynthetic eukaryotes. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Zimorski, V.; Mentel, M.; Tielens, A.G.; Martin, W.F. Energy metabolism in anaerobic eukaryotes and Earth’s late oxygenation. Free Radic. Biol. Med. 2019, 140, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Melser, S.; Lavie, J.; Bénard, G. Mitochondrial degradation and energy metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 2812–2821. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Janeczko, A.; Skoczowski, A. Mammalian sex hormones in plants. Folia Histochem. Cytobiol. 2005, 43, 71–79. [Google Scholar]
- Chantilis, S.; Dombroski, R.; Shackleton, C.H.; Casey, M.L.; MacDonald, P.C. Metabolism of 5 alpha-dihydroprogesterone in women and men: 3 beta-and 3 alpha-, 6 alpha-dihydroxy-5 alpha-pregnan-20-ones are major urinary metabolites. J. Clin. Endocrinol. Metab. 1996, 81, 3644–3649. [Google Scholar]
- Kennedy, T.G. Proceedings: Vaginal mucification in the ovariectomized rat in response to 5alpha-pregnan-3, 20-dione, testosterone and 5alpha-androstan-17beta-ol-3-one: Test for progestagenic activity. J. Reprod. Fertil. 1974, 36, 488–489. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H. Steroid metabolism in the cellular components of the rat ovarian follicle. Nihon Sanka Fujinka Gakkai Zasshi 1987, 39, 1545–1552. [Google Scholar] [PubMed]
- Kozlova, T.A.; Hardy, B.P.; Levin, D.B. Effect of fish steroids 17β-estradiol and 17, 20β-dihydroxy-4-pregnen-3-one on growth, accumulation of pigments, and fatty acid profiles in the microalgae Scenedesmus quadricauda (CPCC-158). Renew. Energy 2020, 148, 798–806. [Google Scholar] [CrossRef]
- Agathokleous, E.; Guo, J.; Peñuelas, J. Low doses of toxicants can enhance algae potential as biodiesel and biomass feedstocks. Renew. Sust. Energy Rev. 2022, 168, 112858. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Hormesis in plants: Its common occurrence across stresses. Curr. Opin. Toxicol. 2022, 30, 100333. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef]
- Fridman, Y.; Savaldi-Goldstein, S. Brassinosteroids in growth control: How, when and where. Plant Sci. 2013, 209, 24–31. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.D.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Human and veterinary antibiotics induce hormesis in plants: Scientific and regulatory issues and an environmental perspective. Environ. Int. 2018, 120, 489–495. [Google Scholar] [CrossRef]
- Murdock, L.; Burke, T.; Coumoundouros, C.; Culham, D.E.; Deutch, C.E.; Ellinger, J.; Wood, J.M. Analysis of strains lacking known osmolyte accumulation mechanisms reveals contributions of osmolytes and transporters to protection against abiotic stress. Appl. Environ. Microbiol. 2014, 80, 5366–5378. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 2006, 18, 2767–2781. [Google Scholar] [CrossRef] [PubMed]
- Brown, G. Defects of thiamine transport and metabolism. J. Inherit. Metab. Dis. 2014, 37, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Goyer, A. Thiamine in plants: Aspects of its metabolism and functions. Phytochemistry 2010, 71, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Rapala-Kozik, M. Vitamin B1 (thiamine): A cofactor for enzymes involved in the main metabolic pathways and an environmental stress protectant. Adv. Bot. Res. 2011, 58, 37–91. [Google Scholar]
- Croce, R.; van Amerongen, H. Light harvesting in oxygenic photosynthesis: Structural biology meets spectroscopy. Science 2020, 369, eaay2058. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, history and modelling. Ann. Bot. 2020, 126, 511–537. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Sharma, R.; Handa, N.; Kaur, H.; Rattan, A.; Yadav, P.; Bhardwaj, R. Redox homeostasis in plants under abiotic stress: Role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 2015, 3, 13. [Google Scholar] [CrossRef]
- Heinemann, B.; Hildebrandt, T.M. The role of amino acid metabolism in signaling and metabolic adaptation to stress-induced energy deficiency in plants. J. Exp. Bot. 2021, 72, 4634–4645. [Google Scholar] [CrossRef]
- Siqueira, J.A.; Hardoim, P.; Ferreira, P.C.; Nunes-Nesi, A.; Hemerly, A.S. Unraveling interfaces between energy metabolism and cell cycle in plants. Trends Plant Sci. 2018, 23, 731–747. [Google Scholar] [CrossRef]
- Crowley, L.C.; Scott, A.P.; Marfell, B.J.; Boughaba, J.A.; Chojnowski, G.; Waterhouse, N.J. Measuring cell death by propidium iodide uptake and flow cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087163. [Google Scholar] [CrossRef]
- Franklin, N.M.; Adams, M.S.; Stauber, J.L.; Lim, R.P. Development of an improved rapid enzyme inhibition bioassay with marine and freshwater microalgae using flow cytometry. Arch. Environ. Contam. Toxicol. 2001, 40, 469–480. [Google Scholar] [PubMed]
- Li, Q.; Lu, D.; Sun, H.; Guo, J.; Mo, J. Tylosin toxicity in the alga Raphidocelis subcapitata revealed by integrated analyses of transcriptome and metabolome: Photosynthesis and DNA replication-coupled repair. Aquat. Toxicol. 2021, 239, 105964. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pathways | Map ID | Map Name | p Value | Down-DAMs | Up-DAMs |
|---|---|---|---|---|---|
| Control vs. 0.001 mg/L erythromycin treatment | |||||
| Lipid metabolism | ko00140 | Steroid hormone biosynthesis | 0.040 | - | 5.alpha.-pregnane-3.alpha.,20.alpha.-diol; 5alpha-androstan-17beta-ol-3-one |
| Control vs. 0.75 mg/L erythromycin treatment | |||||
| Lipid metabolism | ko00590 | Arachidonic acid metabolism | 0.001 | - | 5s-hydroxy-6e,8z,11z,14z-eicosatetraenoic acid; Leukotriene f4; Prostaglandin i2; Prostaglandin a2; PC (16:0/16:0) |
| Lipid metabolism | ko00564 | Glycerophospholipid metabolism | 0.001 | - | PC (16:0/16:0); Phosphocholine; Choline; Glycerophosphocholine |
| Amino acid metabolism | ko00330 | Arginine and proline metabolism | 0.004 | - | Caldine; Guanidoacetic acid; DL-Glutamic acid |
| Membrane transport | ko02010 | ABC transporters | 0.005 | - | Thiamine; N-acetyl-d-glucosamine; Choline; DL-Glutamic acid |
| Lipid metabolism | ko00592 | alpha-Linolenic acid metabolism | 0.005 | - | Linolenic acid; Stearidonic acid; PC (16:0/16:0) |
| Amino acid metabolism | ko00260 | Glycine, serine, and threonine metabolism | 0.008 | - | Ectoine; Choline; Guanidoacetic acid |
| Biosynthesis of other secondary metabolites | ko00332 | Carbapenem biosynthesis | 0.028 | - | DL-Glutamic acid |
| Lipid metabolism | ko00140 | Steroid hormone biosynthesis | 0.046 | - | 5.alpha.-pregnane 3.alpha.,20.alpha.-diol; 5alpha-androstan-17beta-ol-3-one; Aldosterone |
| Control vs. 2.5 mg/L erythromycin treatment | |||||
| Membrane transport | ko02010 | ABC transporters | 1.89 × 10−5 | - | Phosphoric acid; Thiamine; L-glutamate; D-glutamine; Sucrose; Glycerol; N-acetyl-d-glucosamine; Deoxyadenosine; Choline |
| Amino acid metabolism | ko00330 | Arginine and proline metabolism | 0.001 | Caldine; Guanidoacetic acid | L-glutamate; G-guanidinobutyrate; gamma-aminobutyric acid |
| Amino acid metabolism | ko00250 | Alanine, aspartate, and glutamate metabolism | 0.001 | - | L-glutamate; D-glutamine; gamma-aminobutyric acid |
| Translation | ko00970 | Aminoacyl-tRNA biosynthesis | 0.002 | - | L-glutamate; D-glutamine |
| Amino acid metabolism | ko00220 | Arginine biosynthesis | 0.007 | - | L-glutamate; D-glutamine |
| Lipid metabolism | ko00590 | Arachidonic acid metabolism | 0.009 | - | 5s-hydroxy-6e,8z,11z,14z-eicosatetraenoic acid; Prostaglandin i2; PC (16:0/16:0); 15-deoxy-delta-12,14-pgj2; Lipoxin a4 |
| Lipid metabolism | ko00564 | Glycerophospholipid metabolism | 0.012 | - | Phosphocholine; PC (16:0/16:0); Glycerophosphocholine; Choline |
| Metabolism of other amino acids | ko00471 | D-Glutamine and D-glutamate metabolism | 0.020 | - | L-glutamate; D-glutamine |
| Global and overview maps | ko01230 | Biosynthesis of amino acids | 0.022 | Isocitric acid | D-glutamine; L-glutamate |
| Cell growth and death | ko04111 | Cell cycle—yeast | 0.034 | - | Phosphoric acid |
| Lipid metabolism | ko00592 | alpha-Linolenic acid metabolism | 0.039 | - | Stearidonic acid; Linolenic acid; PC (16:0/16:0) |
| Energy metabolism | ko00910 | Nitrogen metabolism | 0.041 | - | D-glutamine; L-glutamate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Tong, Y.; Li, T.; Guo, J.; Liu, W.; Mo, J. Metabolomic Response of Thalassiosira weissflogii to Erythromycin Stress: Detoxification Systems, Steroidal Metabolites, and Energy Metabolism. Plants 2024, 13, 354. https://doi.org/10.3390/plants13030354
Wu X, Tong Y, Li T, Guo J, Liu W, Mo J. Metabolomic Response of Thalassiosira weissflogii to Erythromycin Stress: Detoxification Systems, Steroidal Metabolites, and Energy Metabolism. Plants. 2024; 13(3):354. https://doi.org/10.3390/plants13030354
Chicago/Turabian StyleWu, Xintong, Yongqi Tong, Tong Li, Jiahua Guo, Wenhua Liu, and Jiezhang Mo. 2024. "Metabolomic Response of Thalassiosira weissflogii to Erythromycin Stress: Detoxification Systems, Steroidal Metabolites, and Energy Metabolism" Plants 13, no. 3: 354. https://doi.org/10.3390/plants13030354