Figure 1.
Short-term-effect experiment: mass fractions of rhizomes, stems, and leaves of S. origanifolia adult plants as affected by the Illumination treatment mean effect. Data are means ± SE of two sites and three replicates (n = 6). FS, full sun; CS, canopy shade. *, indicates significant difference for the corresponding mass fraction at p ≤ 0.05, Tukey test.
Figure 1.
Short-term-effect experiment: mass fractions of rhizomes, stems, and leaves of S. origanifolia adult plants as affected by the Illumination treatment mean effect. Data are means ± SE of two sites and three replicates (n = 6). FS, full sun; CS, canopy shade. *, indicates significant difference for the corresponding mass fraction at p ≤ 0.05, Tukey test.
Figure 2.
Short-term-effect experiment. Number of seedlings that emerged from soil seed banks (SB): (a) as affected by the Site × Time interaction, with data means ± SE of two light conditions, two Sa presence treatments, and three replicates (n = 12); (b) as affected by the Site × Illumination × Time interaction, with data means ± SE of two sites and three replicates (n = 6). LD, low density site; HD, high density site; FS, full sun; CS, canopy shade; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Figure 2.
Short-term-effect experiment. Number of seedlings that emerged from soil seed banks (SB): (a) as affected by the Site × Time interaction, with data means ± SE of two light conditions, two Sa presence treatments, and three replicates (n = 12); (b) as affected by the Site × Illumination × Time interaction, with data means ± SE of two sites and three replicates (n = 6). LD, low density site; HD, high density site; FS, full sun; CS, canopy shade; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Figure 3.
Short-term-effect experiment: (a) number of seedlings for each growth form, and (b) relative abundance of growth forms in the seed bank communities. Data area means of two sites and three replicates (n = 6). Within a growth form, the * superscript bar differs significantly at p ≤ 0.05, Tukey test.
Figure 3.
Short-term-effect experiment: (a) number of seedlings for each growth form, and (b) relative abundance of growth forms in the seed bank communities. Data area means of two sites and three replicates (n = 6). Within a growth form, the * superscript bar differs significantly at p ≤ 0.05, Tukey test.
Figure 4.
Short-term-effect experiment: (a) number, (b) biomass, and (c) N, and (d) P content of seed bank (SB) seedlings partitioned into resident and S. origanifolia (Sa SB) seedlings, as affected by the Site × Sa presence interaction. Data area means of two sites and three replicates (n = 6). Within a series, bars with the same letter are not significantly different at p ≤ 0.05, Tukey test. Lowercase letters indicate significance for resident species; uppercase letters for Sa seedlings. In the Sa SB series, the letters C were not indicated. LD, low density site; HD, high density site; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Figure 4.
Short-term-effect experiment: (a) number, (b) biomass, and (c) N, and (d) P content of seed bank (SB) seedlings partitioned into resident and S. origanifolia (Sa SB) seedlings, as affected by the Site × Sa presence interaction. Data area means of two sites and three replicates (n = 6). Within a series, bars with the same letter are not significantly different at p ≤ 0.05, Tukey test. Lowercase letters indicate significance for resident species; uppercase letters for Sa seedlings. In the Sa SB series, the letters C were not indicated. LD, low density site; HD, high density site; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Figure 5.
Short-term-effect experiment: nitrogen concentration of resident and S. origanifolia (Sa SB) seedlings, as affected by the Site × Sa presence interaction. Data area means ± SE of two light conditions, and three replicates (n = 6). LD, low density site; HD, high density site; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Figure 5.
Short-term-effect experiment: nitrogen concentration of resident and S. origanifolia (Sa SB) seedlings, as affected by the Site × Sa presence interaction. Data area means ± SE of two light conditions, and three replicates (n = 6). LD, low density site; HD, high density site; +Sa, presence of Sa adults; −Sa, absence of Sa adults.
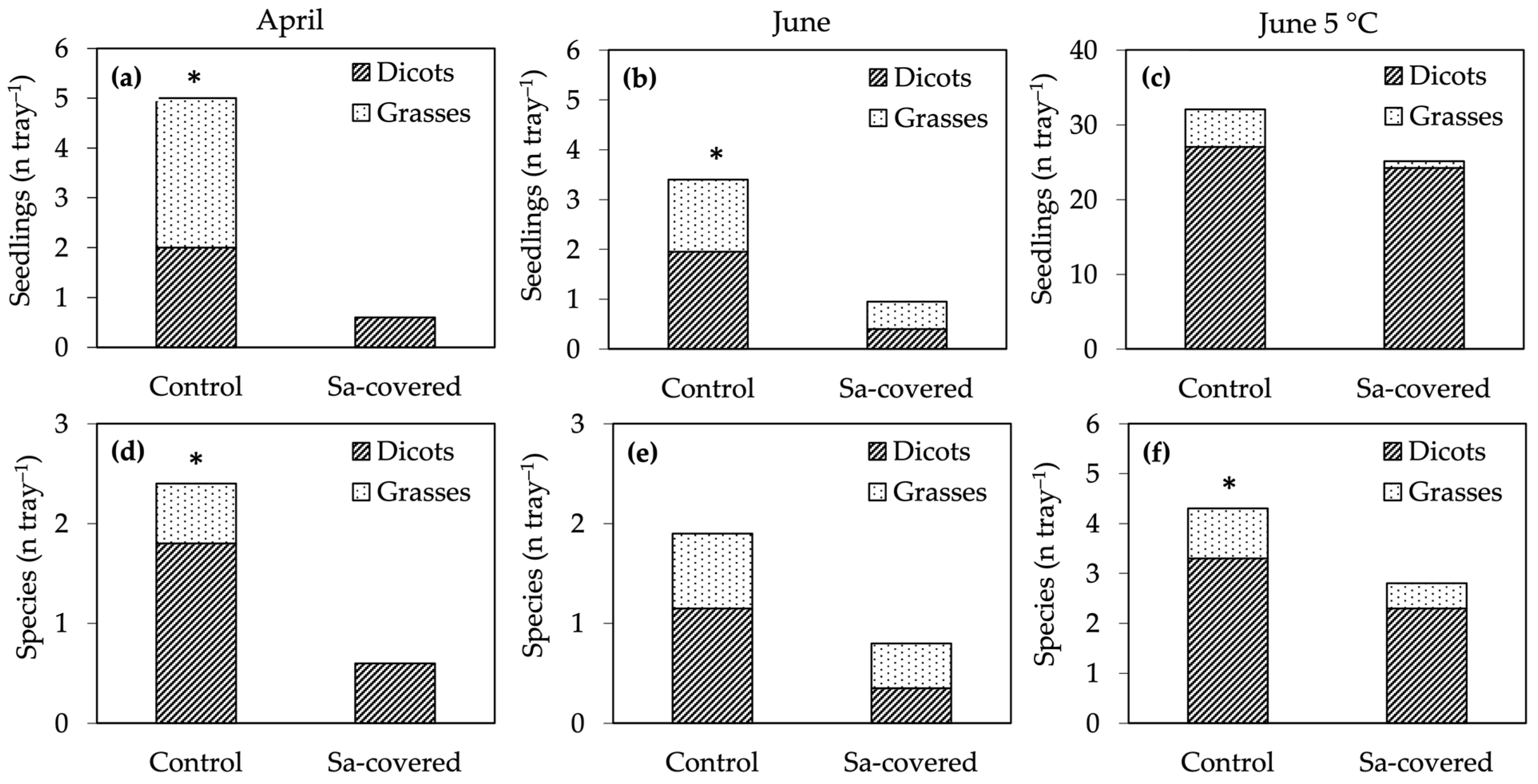
Figure 6.
Long-term-effect experiment: (a–c) number of seed bank seedlings, and (d–f) number of species that emerged from soils collected under established S. origanifolia populations (Sa-covered) and at a 2 m distance (control). Soils were collected in April (a,d) and in June, tested immediately (b,e) and after cold pre-treatment (c,f). Values are means of five replicates (April) and ten replicates (June, June 5 °C). * indicates significant differences for the total SB (Dicots + Grasses) at p ≤ 0.05, t-test.
Figure 6.
Long-term-effect experiment: (a–c) number of seed bank seedlings, and (d–f) number of species that emerged from soils collected under established S. origanifolia populations (Sa-covered) and at a 2 m distance (control). Soils were collected in April (a,d) and in June, tested immediately (b,e) and after cold pre-treatment (c,f). Values are means of five replicates (April) and ten replicates (June, June 5 °C). * indicates significant differences for the total SB (Dicots + Grasses) at p ≤ 0.05, t-test.
Figure 7.
Geographical location of the research area within the Nature Reserve of Migliarino, San Rossore, Massaciuccoli (Pisa, Italy): 1, The Forest of San Rossore; 2, The special protected Forest of San Bartolomeo; 3, The area invaded by S. origanifolia where soil samples were collected.
Figure 7.
Geographical location of the research area within the Nature Reserve of Migliarino, San Rossore, Massaciuccoli (Pisa, Italy): 1, The Forest of San Rossore; 2, The special protected Forest of San Bartolomeo; 3, The area invaded by S. origanifolia where soil samples were collected.
Figure 8.
Long-term experiment: (a) facility for germination tests in the laboratory; (b) the germination test on cold pre-treated soils (June 5 °C) after four weeks.
Figure 8.
Long-term experiment: (a) facility for germination tests in the laboratory; (b) the germination test on cold pre-treated soils (June 5 °C) after four weeks.
Table 1.
Short-term-effect experiment: biomass, and N and P concentrations and contents of S. origanifolia plants sprouted from rhizomes (Sa adults), as affected by the Site and Illumination mean effects. Within each treatment, values followed by different letters in a column are significantly different at p ≤ 0.05, Tukey test (n = 6). LD, low density site; HD, high density site; FS, full sun; CS, canopy shade.
Table 1.
Short-term-effect experiment: biomass, and N and P concentrations and contents of S. origanifolia plants sprouted from rhizomes (Sa adults), as affected by the Site and Illumination mean effects. Within each treatment, values followed by different letters in a column are significantly different at p ≤ 0.05, Tukey test (n = 6). LD, low density site; HD, high density site; FS, full sun; CS, canopy shade.
| Treatment | Sa Adults Biomass | Concentration | Content |
|---|
| N | P | N | P |
|---|
| | g dw pot−1 | mg g−1 | mg g−1 | mg pot−1 | mg pot−1 |
| Site | | | | | |
| LD | 24.7 a | 13.3 a | 1.5 a | 326.7 b | 37.7 a |
| HD | 30.7 a | 15.6 a | 1.6 a | 478.8 a | 50.0 a |
| Illumination | | | | | |
| FS | 24.3 a | 13.1 b | 1.5 a | 317.5 b | 37.7 a |
| CS | 31.0 a | 15.7 a | 1.6 a | 488.1 a | 50.0 a |
Table 2.
Short-term-effect experiment: leaf parameters of S. origanifolia adult plants as affected by the Illumination treatment mean effect. Data are means of two sites and three replicates (n = 6). Values followed by the same letter within a column are not significantly different at p ≤ 0.05, Tukey test. FS, full sun; CS, canopy shade; SLA, Specific Leaf Area.
Table 2.
Short-term-effect experiment: leaf parameters of S. origanifolia adult plants as affected by the Illumination treatment mean effect. Data are means of two sites and three replicates (n = 6). Values followed by the same letter within a column are not significantly different at p ≤ 0.05, Tukey test. FS, full sun; CS, canopy shade; SLA, Specific Leaf Area.
| Illumination | Leaf Dry Weight | Leaf Area | SLA | Chlorophyll Concentration |
|---|
| | g pot−1 | mg leaf−1 | cm2 leaf−1 | mm2 mg−1 | μg cm−2 | mg g−1 |
| FS | 4.3 b | 14.7 b | 3.0 b | 20.0 a | 10.9 b | 0.22 b |
| CS | 6.9 a | 22.2 a | 4.9 a | 21.9 a | 18.1 a | 0.41 a |
Table 3.
Short-term-effect experiment: species recorded at least twice in the seed bank of soils with a low (LD) and high (HD) density of S. origanifolia. x, present; -, absent. Relative abundance of growth forms, and of S. origanifolia, as affected by the site mean effect. For each growth form, values followed by the same letter are not significantly different at p ≤ 0.05, Tukey test (n = 12). Soils were collected in the San Bartolomeo Forest on 5 March.
Table 3.
Short-term-effect experiment: species recorded at least twice in the seed bank of soils with a low (LD) and high (HD) density of S. origanifolia. x, present; -, absent. Relative abundance of growth forms, and of S. origanifolia, as affected by the site mean effect. For each growth form, values followed by the same letter are not significantly different at p ≤ 0.05, Tukey test (n = 12). Soils were collected in the San Bartolomeo Forest on 5 March.
| Growth Form/Species | Family | LD | HD |
|---|
| Trees | | | |
| Laurus nobilis | Lauraceae | - | x |
| Fraxinus angustifolia | Oleaceae | x | - |
| Ulmus sp. | Ulmaceae | - | x |
| Climbers | | | |
| Clematis vitalba | Ranunculaceae | x | x |
| Hedera helix | Araliaceae | - | x |
| Periploca graeca | Apocynaceae | x | - |
| Rubus sp. | Rosaceae | x | x |
| Salpichroa origanifolia | Solanaceae | x | x |
| Dicotyledon herbs | | | |
| unknown cfr Sonchus sp. | Asteraceae | x | x |
| Cerastium sp. | Caryophyllaceae | x | - |
| Moehringia trinervia | Caryophyllaceae | x | x |
| Stellaria media | Caryophyllaceae | x | x |
| Trifolium sp. | Fabaceae | x | x |
| Oxalis corniculata | Oxalidaceae | - | x |
| unknown cfr Veronica sp. | Plantaginaceae | x | - |
| Persicaria hydropiper | Polygonaceae | x | x |
| unknown | Solanaceae | - | x |
| Grasses | | | |
| unknown cfr. Poa annua | Poaceae | x | x |
| Carex sp. | Cyperaceae | x | x |
| Relative abundance (% of total seedlings) | | |
| Trees | | 0.9 a | 1.7 a |
| Climbers | | 7.9 b | 54.2 a |
| Dicot herbs | | 46.5 a | 8.7 b |
| Grasses | | 44.7 a | 35.4 a |
| Salpichroa origanifolia | | 0.9 b | 12.2 a |
Table 4.
Short-term-effect experiment: number, biomass, and N and P content of resident, S. origanifolia, and total seed bank (SB) seedlings, and percentage of residents on total SB values, as affected by the Illumination × Sa presence interaction. Data are means of two sites and three replicates (n = 6). Within each row, values followed by the same letter are not significantly different at p ≤ 0.05, Tukey test. +Sa, presence of Sa adults; −Sa, absence of Sa adults.
Table 4.
Short-term-effect experiment: number, biomass, and N and P content of resident, S. origanifolia, and total seed bank (SB) seedlings, and percentage of residents on total SB values, as affected by the Illumination × Sa presence interaction. Data are means of two sites and three replicates (n = 6). Within each row, values followed by the same letter are not significantly different at p ≤ 0.05, Tukey test. +Sa, presence of Sa adults; −Sa, absence of Sa adults.
| | Full Sun | Canopy Shade |
|---|
| SB Seedlings | +Sa | −Sa | +Sa | −Sa |
|---|
| Number (n pot−1) | | | |
| Residents | 81.5 a | 45.5 b | 50.5 b | 55.5 ab |
| S. origanifolia | 5.5 a | 2.5 a | 6.0 a | 4.5 a |
| Total | 87.0 a | 48.0 b | 56.5 b | 60.0 ab |
| Residents n° (%) | 93.7 a | 94.8 a | 89.4 a | 92.5 a |
| Biomass (g dw pot−1) | | | |
| Residents | 8.0 a | 9.6 a | 3.8 a | 9.9 a |
| S. origanifolia | 1.8 b | 6.9 a | 1.9 b | 8.7 a |
| Total | 9.8 b | 16.5 a | 5.7 b | 18.6 a |
| Residents dw (%) | 81.8 a | 58.3 b | 66.9 ab | 53.2 d |
| N content (mg pot−1) | | | |
| Residents | 97.8 ab | 105.3 ab | 54.2 b | 160.2 a |
| S. origanifolia | 14.8 b | 88.2 a | 17.8 b | 108.7 a |
| Total | 112.6 bc | 193.5 ab | 72.0 c | 268.9 a |
| Residents N (%) | 86.9 a | 54.4 b | 75.2 a | 59.6 b |
| P content (mg pot−1) | | | |
| Residents | 25.3 a | 27.9 a | 11.6 b | 32.1 a |
| S. origanifolia | 3.8 b | 16.3 a | 3.9 b | 18.2 a |
| Total | 29.1 a | 44.2 a | 15.5 b | 50.3 a |
| Residents P (%) | 86.9 a | 63.2 b | 74.8 ab | 63.9 b |
Table 5.
Long-term-effect experiment: species or types that emerged from soil samples collected in April and June under established populations of S. origanifolia (Sa) and at a 2 m distance (Ctrl). A subset of soil collected in June was cold pre-treated (J 5 °C) for three months. x, species recorded in at least one tray.
Table 5.
Long-term-effect experiment: species or types that emerged from soil samples collected in April and June under established populations of S. origanifolia (Sa) and at a 2 m distance (Ctrl). A subset of soil collected in June was cold pre-treated (J 5 °C) for three months. x, species recorded in at least one tray.
| | | April | June | June 5 °C |
|---|
| Species/Type | Family | Ctrl | Sa | Ctrl | Sa | Ctrl | Sa |
|---|
| Dysphania botrys | Amaranthaceae | x | - | - | - | - | x |
| Periploca graeca | Apocynaceae | x | - | - | - | - | - |
| unknown | Asteraceae | - | - | - | - | x | - |
| Erigeron sp. | Asteraceae | - | - | - | - | x | - |
| Myosotis sp. cfr | Boraginaceae | - | - | - | - | x | - |
| Cardamine hirsuta | Brassicaceae | - | - | - | - | x | x |
| Cerastium glomeratum | Caryophyllaceae | - | - | x | - | x | x |
| Moheringia trinervia | Caryophyllaceae | - | - | - | - | x | x |
| Polycarpon tetraphyllum | Caryophyllaceae | x | - | - | - | x | - |
| Silene sp. | Caryophyllaceae | - | - | x | - | x | - |
| Stellaria media | Caryophyllaceae | - | - | - | - | x | x |
| Carex sp. 1 | Cyperaceae | x | - | x | - | x | - |
| Carex sp. 2 | Cyperaceae | - | - | x | x | x | x |
| Erica sp. | Ericaceae | x | - | - | - | - | - |
| Euphorbia peplus | Euphorbiaceae | - | - | x | x | x | x |
| Lotus sp. | Fabaceae | - | - | - | x | - | - |
| Trifolium sp. | Fabaceae | - | - | x | - | - | - |
| Juncus inflexus | Juncaceae | - | - | - | - | x | x |
| Oxalis corniculata | Oxalidaceae | - | - | x | - | x | x |
| Veronica arvensis | Plantaginaceae | - | - | x | x | x | x |
| Poa annua | Poaceae | - | - | x | x | x | x |
| unknown | Poaceae | - | x | x | x | x | x |
| Clematis vitalba | Ranunculaceae | x | x | - | - | - | - |
| Ranunculus cfr muricatus | Ranunculaceae | - | - | - | - | x | x |
| Solanum nigrum | Solanaceae | - | - | - | - | x | - |
| unknown | Solanaceae | - | x | x | x | x | - |
| Urtica dioica | Urticaceae | - | - | - | - | - | x |
| Total n° of species | | 6 | 3 | 11 | 7 | 20 | 14 |
Table 6.
Short-term-effect experiment: main physical and chemical soil properties of the sites with low (LD) and high (HD) coverage of S. origanifolia in the San Bartolomeo Forest. Soil samples were collected on 5 March from the 0–10 cm profile.
Table 6.
Short-term-effect experiment: main physical and chemical soil properties of the sites with low (LD) and high (HD) coverage of S. origanifolia in the San Bartolomeo Forest. Soil samples were collected on 5 March from the 0–10 cm profile.
| Soil Properties | Unit | LD | HD |
|---|
| Sand (2 mm > Ø > 0.05 mm) | % | 77.6 | 87.7 |
| Silt (0.05 mm > Ø > 0.002 mm) | % | 16.3 | 6.9 |
| Clay (Ø < 0.002 mm) | % | 6.1 | 5.4 |
| Soil Organic C | % | 13.9 | 9.3 |
| pH | - | 6.6 | 6.6 |
| Total N | g kg−1 | 18.7 | 11.2 |
| C/N | - | 7.3 | 8.1 |
| Cation Exchange Capacity | µS cm−1 | 349.5 | 266.0 |
Table 7.
Short-term-effect experiment: illuminance (lx) and % illuminance reduction in CS, recorded in the full sun (FS) and canopy shade (CS) illumination treatments at leaf onset (21 March) and at complete leaf unfolding (2 May) of the plane tree.
Table 7.
Short-term-effect experiment: illuminance (lx) and % illuminance reduction in CS, recorded in the full sun (FS) and canopy shade (CS) illumination treatments at leaf onset (21 March) and at complete leaf unfolding (2 May) of the plane tree.
| Day Time (h) | Illumination | 21 March | 2 May |
|---|
| 09:00 a.m. | FS | 12,372 | 42,540 |
| | CS | 5720 | 847 |
| | CS reduction (%) | 54 | 98 |
| 12:00 a.m. | FS | 37,733 | 59,333 |
| | CS | 18,067 | 9000 |
| | CS reduction (%) | 52 | 85 |













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}