
Influence of Orographic Factors on the Distribution of Lichens in the Franz Josef Land Archipelago

Abstract
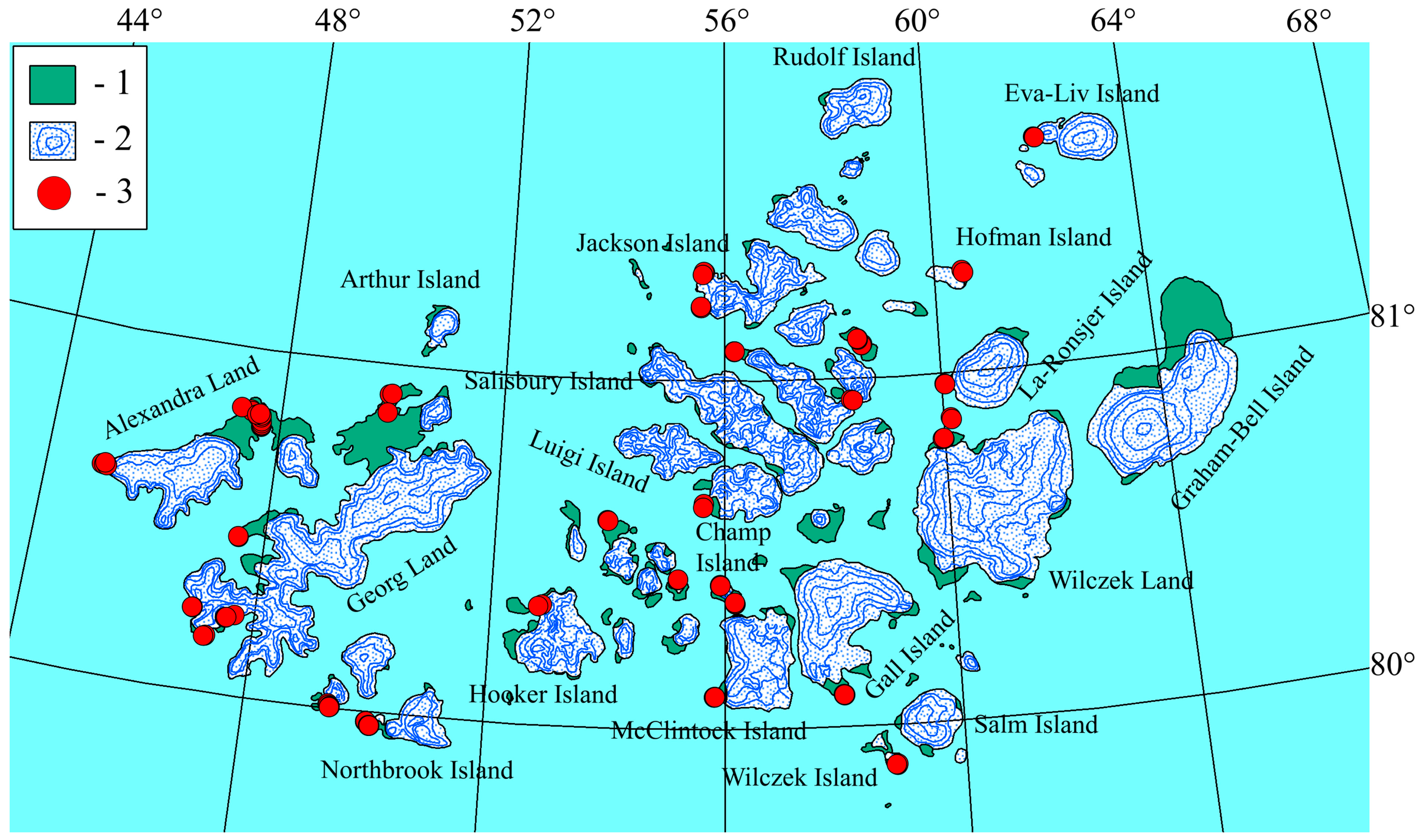
:1. Introduction
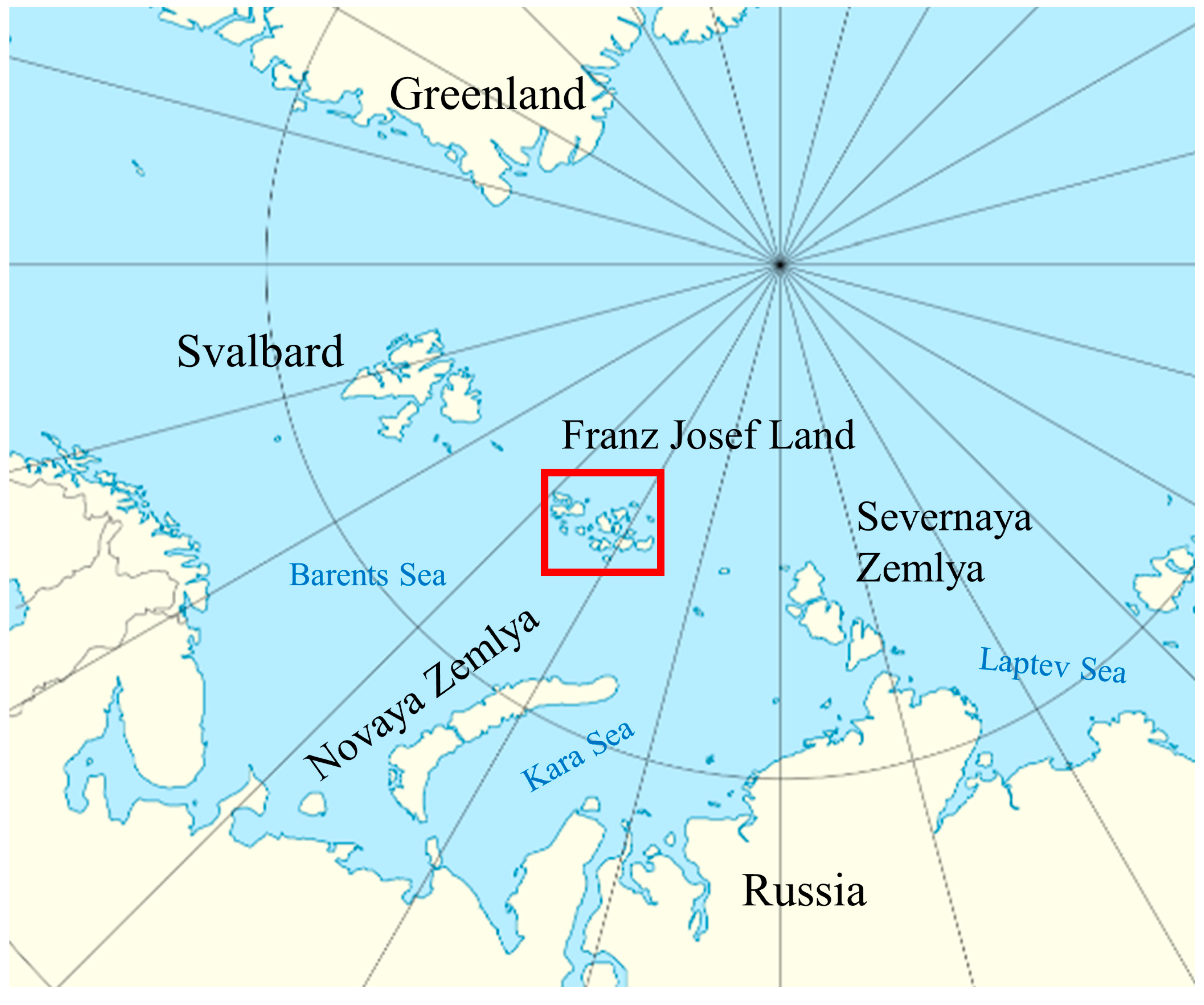
The Natural Conditions of the Research Area
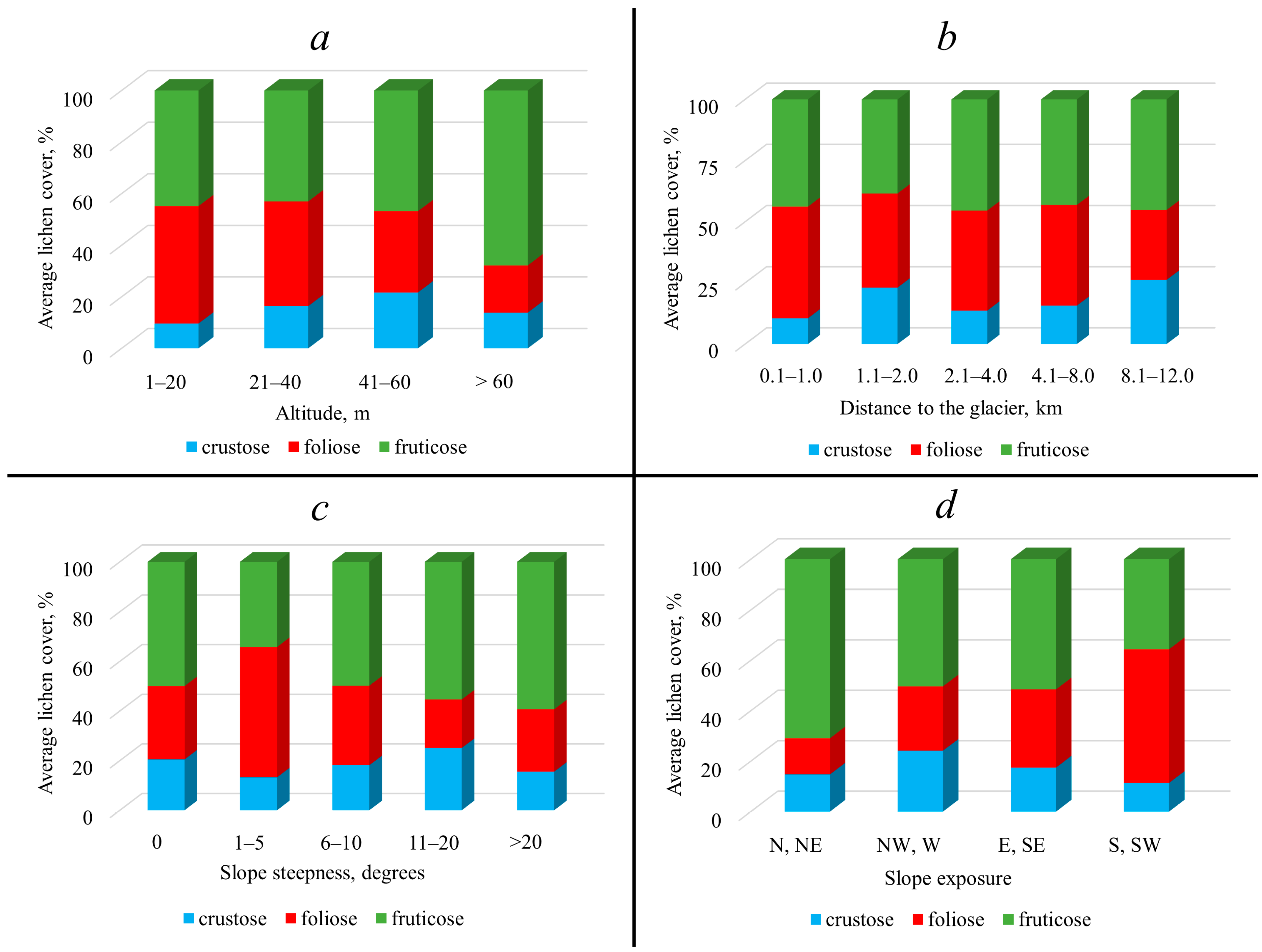
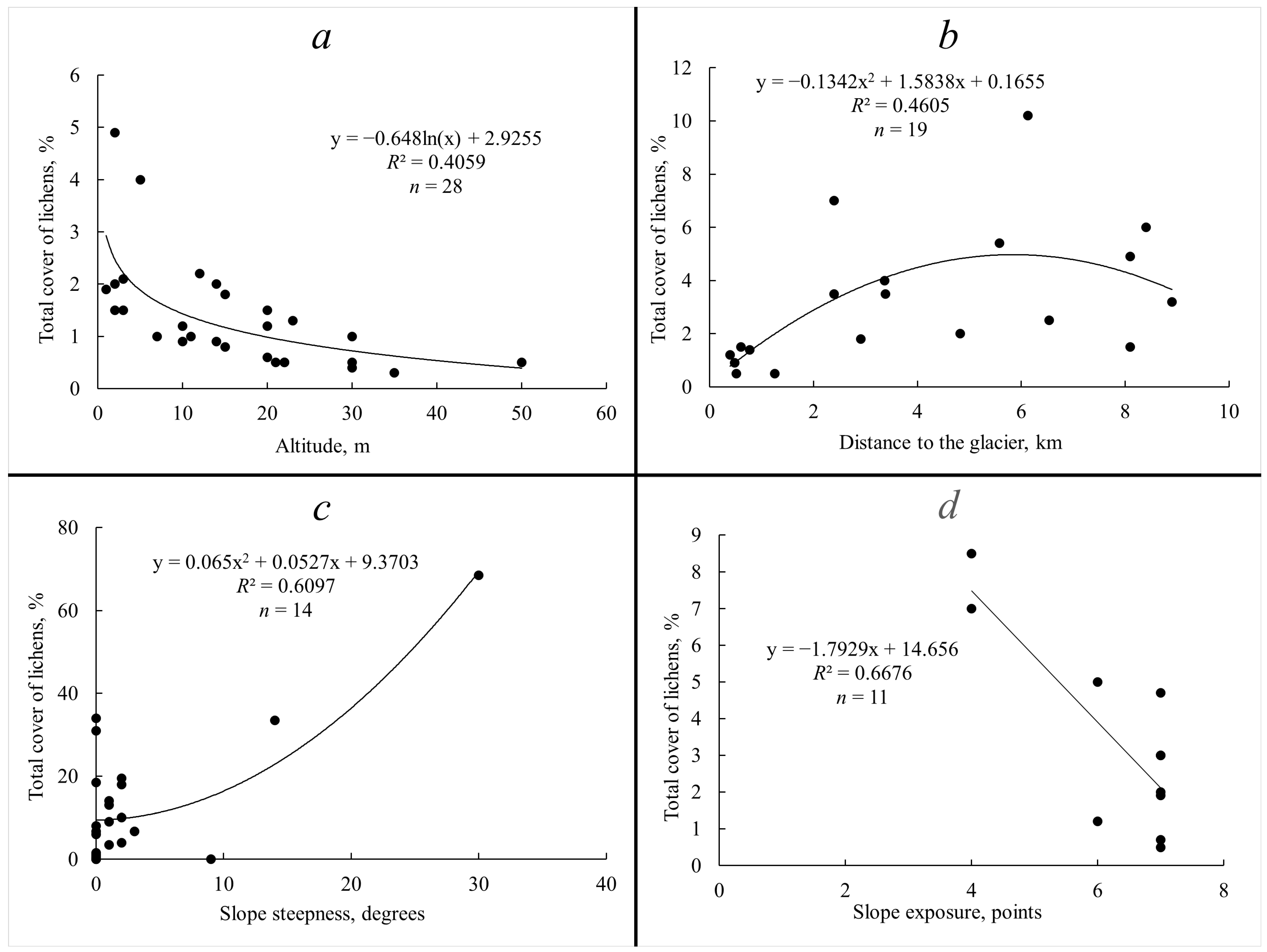
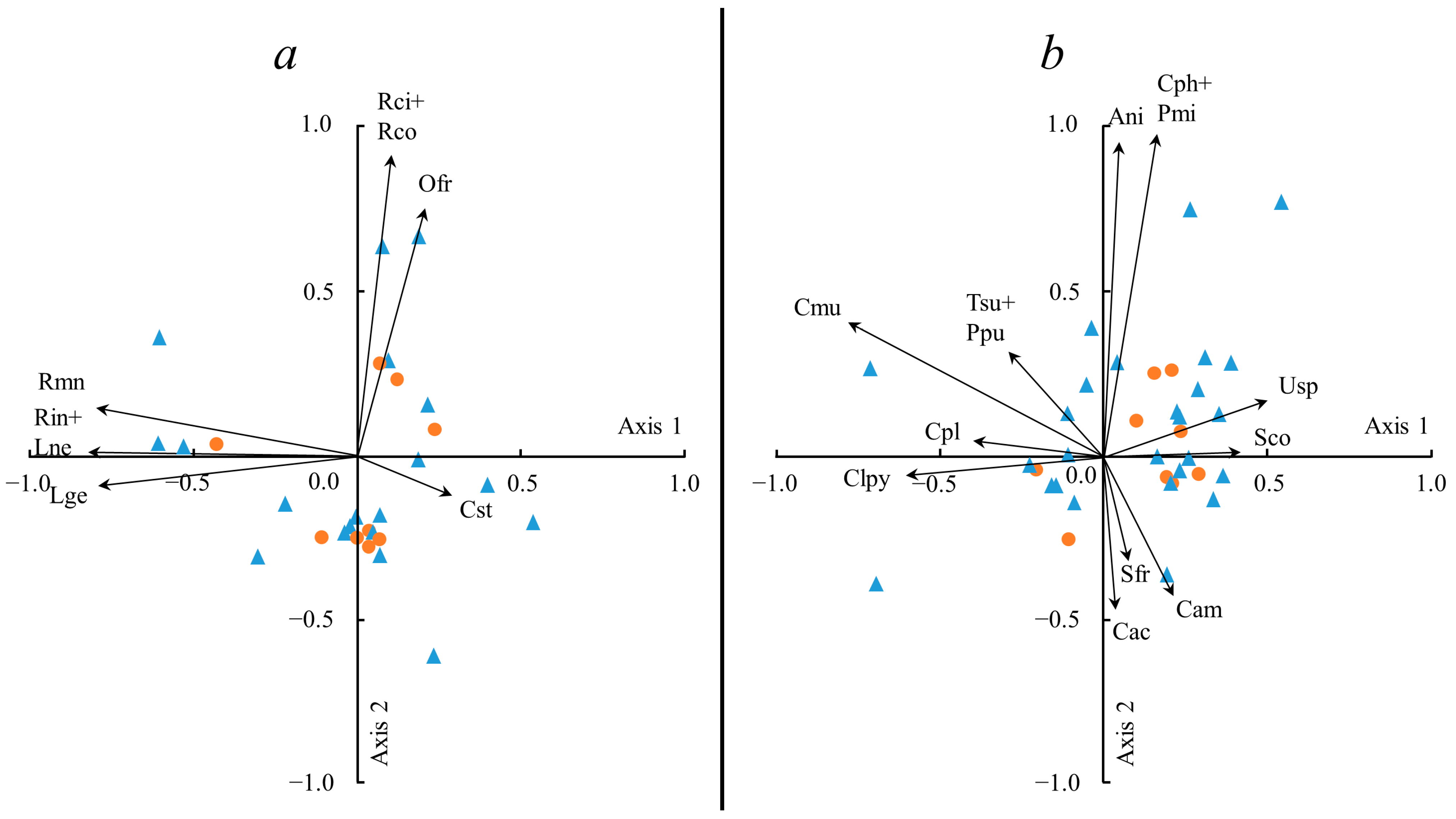
2. Results
3. Discussion
4. Materials and Methods
4.1. Lichen Collection and Laboratory Studies
4.2. Geobotanical Research
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahzelt, D.; Maycockj, P.; Svoboda, J. Initial establishment of saxicolous lichens following recent glacial recession in Sverdrup Pass, Ellesmere Island, Canada. Lichenologist 1988, 20, 253–268. [Google Scholar] [CrossRef]
- Buschbom, J.; Kappen, L. The role of microclimate for the lichen vegetation pattern on rock surfaces in the Subarctic. Sauteria 1998, 9, 79–94. [Google Scholar]
- Daniëls, F.J.A. Vegetation of the Angmagssalik district, southeast Greenland. III. Epilithic macrolichen communities. Meddelelser Om Grønland 1975, 198, 1–32. [Google Scholar]
- John, E.; Dale, M.R.T. Environmental correlates of species distributions in a saxicolous lichen community. J. Veg. Sci. 1990, 1, 385–392. [Google Scholar] [CrossRef]
- Link, S.O.; Nash, T.H. An analysis of an arctic lichen community with respect to slope on silicious rocks at Anaktuvuk Pass, Alaska. Bryologist 1984, 87, 162–166. [Google Scholar] [CrossRef]
- Grytnes, J.A.; Heegaard, E.; Ihlen, P.G. Species richness of vascular plants, bryophytes, and lichens along an altitudinal gradient in western Norway. Acta Oecologica 2006, 29, 241–246. [Google Scholar] [CrossRef]
- Pinokiyo, A.; Singh, K.P.; Singh, J.S. Diversity and distribution of lichens in relation to altitude within a protected biodiversity hot spot, north-east India. Lichenologist 2008, 40, 47–62. [Google Scholar] [CrossRef]
- Baniya, C.B.; Rai, H.; Upreti, D.K. Terricolous lichens in Himalauas: Patterns of species richness along elevation gradient. In Terricolous lichens in India. V. 1. Diversity Patterns and Distribution Ecology; Rai, H., Upreti, D.K., Eds.; Springer: New York, NY, USA, 2013; pp. 33–52. [Google Scholar]
- Gupta, S.; Khare, R.; Rai, H.; Upreti, D.K.; Gupta, R.K.; Sharma, P.K.; Srivastava, K.; Bhattacharya, P. Influence of macro-scale environmental variables on diversity and distribution pattern of lichens in Badrinath valley, Western Himalaya. Mycosphere 2014, 5, 229–243. [Google Scholar] [CrossRef]
- Brodo, I.M. Substrate ecology. In The Lichens; Ahmajian, V., Hale, M.E., Eds.; Academic Press: New York, NY, USA, 1978; pp. 401–441. [Google Scholar]
- Belnap, J.; Lange, O.L. (Eds.) Biological Soil Crusts: Structure, Function and Management; Springer: Berlin/Heidelberg, Germany, 2001; 503p. [Google Scholar]
- Breen, K.; Levesque, E. Proglacial succession of biological soil crusts and vascular plants: Biotic interactions in the High Arctic. Can. J. Bot. 2006, 84, 1714–1731. [Google Scholar] [CrossRef]
- Elven, R. Association analysis of moraine vegetation at the glacier Hardangerjokulen, Finse, South Norway. Norweg. J. Bot. 1978, 25, 172–191. [Google Scholar]
- Smith, R.I.L. Colonization by lichens and development of lichen-dominated communities in the maritime Antarctic. Lichenologist 1995, 27, 473–483. [Google Scholar] [CrossRef]
- Dzhenyuk, S.L. Climate-forming factors and climatic specific features of the Franz-Josef Land Region. Trudy Kol’skogo nauchnogo tsentra RAN 2014, 4, 61–69. (In Russian) [Google Scholar]
- Boyarskiy, P.V. (Ed.) Franz Josef Land; Paulson: Moscow, Russia, 2013; 680p. (In Russian) [Google Scholar]
- Groswald, M.G.; Krenke, A.N.; Vinogradov, A.N.; Markin, V.A.; Psareva, T.V.; Razumeiko, N.G.; Sukhodrovsky, V.L. Glaciation of Franz Josef Land; Nauka: Moscow, Russia, 1973; 352p. (In Russian) [Google Scholar]
- Govorukha, L.S. Landscape and geographical characteristics of Franz Josef Land. Trudy AANII 1968, 285, 86–117. (In Russian) [Google Scholar]
- Milovanova, M.S.; Novikov, V.Y.; Demyanov, A.A. Study of the dynamics of changes in the coastlines of the islands of the Franz Josef Land archipelago based on satellite imagery materials. Izv. Vuzov “Geod. Aerophotosurveying” 2012, 1, 18–22. (In Russian) [Google Scholar]
- Überla, K. Faktorenanalise; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1980; 398p. [Google Scholar]
- Kappen, L. Chapter II. B. 2: Ecophysiological relationships in different climatic regions. In Handbook of Lichenology; Galun, M., Ed.; CRC Press: Boca Rato, FL, USA, 1988; Volume 2, pp. 37–94. [Google Scholar]
- Kukwa, M.; Zhurbenko, M.P. Notes on the lichen genus Lepraria from the Arctic. Graph. Scr. 2010, 22, 3–8. [Google Scholar]
- Hansen, E.S. A contribution to the lichen flora of J. A. D. Jensens Nunatakker, Frederikshåb district, South-West Greenland. Cryptog. Mycol. 2010, 31, 201–210. [Google Scholar]
- Węgrzin, M. New records of lichens and lichenicolous fungi from the polish Tatra mountains. Polish Bot. J. 2008, 53, 163–168. [Google Scholar]
- Malíček, J.; Palice, Z.; Bouda, F.; Knudsen, K.; Šoun, J.; Vondrák, J.; Novotný, P. Atlas of Czech Lichens. 2023. Available online: https://dalib.cz/en/ (accessed on 13 November 2023).
- Armstrong, R.A. Adaptation of lichens to extreme conditions. In Plant Adaptation Strategies in Changing Environments; Shukla, V., Kumar, S., Kumar, N., Eds.; Springer Nature: Singapore, 2017; pp. 1–27. [Google Scholar]
- Rai, H.; Khare, R.; Nayaka, S.; Upreti, D.K. The influence of water variables on the distribution of terricolous lichens in Garhwal Himalayas. In Souvenir, Water and Biodiversity; Kumar, P., Singh, P., Srivastava, R.J., Eds.; Uttar Pradesh State biodiversity board: Lucknow, India, 2013; Volume 7, pp. 75–83. [Google Scholar]
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. (Eds.) The Lichens of Great Britain and Ireland; The British Lichen Society: London, UK, 2009; 1046p. [Google Scholar]
- Stepanchikova, I.S.; Gagarina, L.V. Chapter 8. Collection, identification and storage of lichenological collections. In The Lichen Flora of Russia. Biology, Ecology, Diversity, Distribution and Methods to Study Lichens; Andreev, M.P., Himelbrant, D.E., Eds.; KMK Scientific Press Ltd.: Moscow/St. Petersburg, Russia, 2014; pp. 204–219. (In Russian) [Google Scholar]
- Westberg, M.; Moberg, R.; Myrdal, M.; Nordin, A.; Ekman, S. Santesson′s Checklist of Fennoscandian Lichen-Forming and Lichenicolous Fungi; Museum of Evolution, Uppsala University: Uppsala, Sweden, 2021; 933p. [Google Scholar]
- Lumbsch, H.T.; Huhndorf, S.M. Myconet Volume 14. Part One. Outline of Ascomycota—2009. Part Two. Notes on Ascomycete Systematics. Nos 4751–5113. Fieldiana Life Earth Sci. 2010, 1, 1–64. [Google Scholar] [CrossRef]
- Chesnokov, S.V.; Davydov, E.A.; Konoreva, L.A.; Prokopiev, I.A.; Poryadina, L.N.; Zheludeva, E.V.; Shavarda, A.L. The monotypic genus Flavocetraria and two new genera: Cladocetraria and Foveolaria, in the cetrarioid core. Plant Syst. Evol. 2023, 309, 24. [Google Scholar] [CrossRef]
- Rychagov, G.I. General Geomorphology: Textbook; Moscow State University named after M.V. Lomonosov: Moscow, Russia, 2006; 416p. (In Russian) [Google Scholar]
- Ramenskii, L.G. Selected works. In Problems and Methods of Studying Vegetation Cover; Nauka: St. Petersburg, Russian, 1971; 335p. (In Russian) [Google Scholar]
- Shaukat, S.S.; Rao, T.A.; Khan, M.A. Impact of sample size on principal component analysis ordination of an environmental data set: Effects on eigenstructure. Ekológia 2016, 35, 173–190. [Google Scholar] [CrossRef]
- Puzachenko, Y.G. Mathematical Methods in Environmental and Geographical Research: A Textbook for Students; Universities Academia: Moscow, Russia, 2004; 416p. (In Russian) [Google Scholar]
- Ter Braak, C.J.F.; Prentice, I.C. A theory of gradient analysis. Adv. Ecol. Res. 2004, 34, 235–282. [Google Scholar] [CrossRef]
- Kempton, R.A. The use of biplots in interpreting variety by environment interactions. J. Agric. Sci. 1984, 103, 123–135. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/IML® Studio 14.2: User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2016; 576p. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name\Parameter | The Shape of the Mesorelief, within Which the Microrelief Is Formed | The Steepness of the Slope Elements, Degrees | Diameter, m | The Excess of Relief Elements over Each Other, m | Granulometric Composition, Processes of Rock Destruction | Nanorelief Elements, Secondary Polygons, Diameter, m | Elements of Micro/Nanorelief, to Which Lichens Are Confined |
|---|---|---|---|---|---|---|---|
| Stony–gravelly polygons | High-sea and postglacial plains | 0–3 | 0.6–1.2 | 0.1–0.3 | Small rubble and gravel | 0.2–0.3 | Contact zones of the polygon and stone rim |
| Polygons–meshes | Marine terraces of different levels | 0 | 0.2–2.0 | – | Small boulders | – | Small boulders in meshes and depressions |
| Fine-grained and gravelly polygons | Accumulative plains with flattened surfaces | 0 | 0.2–0.3 | – | Sandy loam, light loam; fine crushed stone, gravel | – | Cracks between polygons |
| Gravelly polygons | Low marine terraces | 35–40 | 25–40 | 0.7–0.8 | Rubble | 5–15 | Cracks between polygons |
| Loamy polygons | Marine or postglacial accumulative hollow–humped plains | 1–3 | 0.15–0.35 | 0.02–0.05 | Medium loam | – | Cracks between polygons |
| Spots in the moss carpets | Marine-accumulative (hilly ridge) and abrasive-accumulative plains | 0–5 | 0.3–1.0 | 0.02–0.04 | Medium or heavy loam | – | Contact areas of spot and moss cover |
| Spots in the liverwort carpets | Marine-accumulative (hilly ridge) and abrasive-accumulative plains | 0–5 | 0.8–1.2, irregular shaped spots | 0.03–0.06 | Sand, sandy loam, light loam with an admixture of fine crushed stone | – | Contact areas of spot and liverwort carpet |
| Slope strips | Slopes of basalt plateaus | 15–25 | 0.2–1.0 | 0–0.15 | Loam (light and medium), crushed stone; flagstone, small blocks | – | Contact zones between strips with different granulometric composition |
| Slope steps | Slopes of basalt plateaus | 30–40—the main surface; 10–12—terrace platform-steps | 0.3–1.0 | 0.4–0.7 | Medium loam, crushed stone, flagstone | – | The edge part of the steps, the side of the underlying step |
| Loam–peat mounds | Sea-accumulative plains | 1–35 | 3.0–4.0 | 0.5 (exceeding the top over the foot) | Peat, medium loam; formation of frost-breaking cracks | Small peat fragments | Top, slopes and foot of a peat hillock |
| Blocks of basalt | Marine abrasive-accumulative terraces | 0–90 | 1.0–2.5 | 1.2–1.5 (exceeding the top over the foot) | Basalts; flaking of rock fragments, laying of cracks | – | Cracks in the rock |
| Altitude, m | Distance to the Glacier, km | Steepness Range, Degrees | Slope Exposure | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–20 | 21–40 | 41–60 | >60 | 0.1–1.0 | 1.1–2.0 | 2.1–4.0 | 4.1–8.0 | 8.1–12.0 | 0 | 1–5 | 6–10 | 11–20 | >20 | N, NE | W, NW | E, SE | S, SW | |
| Crustose lichens | ||||||||||||||||||
| Agonimia gelatinosa (Ach.) M. Brand & Diederich | + | + | + | + | ||||||||||||||
| Arthrorhaphis alpina (Schaer.) R. Sant. | + | + | + | + | + | + | ||||||||||||
| Baeomyces carneus Flörke | 1.0 ± 0.7 | + | + | 4.1 ± 2.8 | 4.3 ± 3.2 | + | + | + | 1.0 ± 0.7 | + | ||||||||
| Biatora ementiens (Nyl.) Printzen | + | + | + | + | + | + | + | + | + | + | ||||||||
| Blastenia ammiospila (Wahlenb.) Arup et al. | + | + | + | + | + | + | + | + | + | + | ||||||||
| Bryoplaca jungermanniae (Vahl) Søchting et al. | + | + | + | + | + | |||||||||||||
| Caloplaca stillicidiorum (Vahl) Lynge | + | + | 1.0 ± 0.3 | + | + | + | + | + | + | + | 1.0 ± 0.3 | + | + | |||||
| Candelariella aurella (Hoffm.) Zahlbr. | + | + | + | + | + | + | + | + | ||||||||||
| C. cf. canadensis H. Magn. | 1.1 ± 0.7 | + | 2.0 ± 0.6 | 2.0 ± 1.3 | + | 1.1 ± 0.7 | ||||||||||||
| Helocarpon crassipes Th. Fr. | + | + | + | + | + | + | + | + | + | |||||||||
| Japewia tornoënsis (Nyl.) Tønsberg | + | + | + | + | ||||||||||||||
| Lecanora epibryon (Ach.) Ach. | + | + | 1.0 ± 0.4 | + | + | 2.0 ± 0.7 | + | + | + | + | 1.0 ± 0.7 | + | 1.0 ± 0.6 | + | ||||
| L. polytropa (Ehrh. ex Hoffm.) Rabenh. | + | + | + | + | + | + | + | + | + | |||||||||
| Lecidea lapicida (Ach.) Ach. | + | + | + | + | + | + | + | |||||||||||
| L. ramulosa Th. Fr. | + | + | + | + | + | + | + | |||||||||||
| Lepraria caesioalba (B. de Lesd.) J. R. Laundon | + | + | + | |||||||||||||||
| L. gelida Tønsberg & Zhurb. | + | + | 1.1 ± 0.5 | + | + | + | + | + | + | + | 1.0 ± 0.7 | + | + | + | + | |||
| L. neglecta (Nyl.) Lettau | + | + | + | + | + | + | + | + | + | + | + | |||||||
| Megaspora verrucosa (Ach.) Hafellner & V. Wirth | + | + | + | + | + | + | + | + | ||||||||||
| Micarea incrassata Hedl. | + | + | + | + | + | + | + | + | ||||||||||
| M. lignaria (Ach.) Hedl. | + | + | + | + | + | + | ||||||||||||
| Miriquidica lulensis (Hellb.) Hertel & Rambold | + | + | + | + | ||||||||||||||
| Ochrolechia frigida (Sw.) Lynge | 2.2 ± 0.4 | 2.5 ± 0.6 | 2.6 ± 1.8 | 3.5 ± 2.3 | 1.2 ± 0.2 | 1.7 ± 0.9 | 3.2 ± 1.4 | 3.8 ± 0.7 | 4.4 ± 1.3 | 3.2 ± 0.7 | 2.0 ± 0.4 | 1.7 ± 0.5 | 3.9 ± 2.8 | 1.0 ± 0.2 | 1.9 ± 1.0 | 2.9 ± 1.5 | 2.1 ± 0.6 | 1.6 ± 0.4 |
| Parvoplaca tiroliensis (Zahlbr.) Arup et al. | + | + | 1.0 ± 0.4 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Pertusaria geminipara (Th. Fr.) C. Knight ex Brodo | + | 2.0 ± 1.4 | 1.6 ± 1.3 | 1.0 ± 0.4 | + | 2.0 ± 1.4 | 1.0 ± 0.4 | 2.0 ± 1.4 | 1.0 ± 0.4 | |||||||||
| Polyblastia gothica Th. Fr. | + | + | + | + | ||||||||||||||
| Porpidia melinodes (Körb.) Gowan & Ahti | + | + | + | + | ||||||||||||||
| Protomicarea limosa (Ach.) Hafellner | + | + | + | + | + | + | ||||||||||||
| Protopannaria pezizoides (Weber) P. M. Jørg. & S. Ekman | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Protothelenella sphinctrinoidella (Nyl.) H. Mayrhofer & Poelt | + | + | + | + | + | |||||||||||||
| Psoroma hypnorum (Vahl) Gray | + | 1.0 ± 0.4 | + | + | + | + | + | + | 1.3 ± 0.8 | + | + | + | 1.8 ± 1.2 | + | + | 1.4 ± 0.6 | + | + |
| Rhizocarpon cinereovirens (Müll. Arg.) Vain. | 1.0 ± 0.7 | 1.0 ± 0.5 | 1.0 ± 0.6 | |||||||||||||||
| R. copelandii (Körb.) Th. Fr. | + | + | + | + | + | + | + | |||||||||||
| R. geminatum Körb. | 1.0 ± 0.6 | + | + | + | + | + | + | + | + | |||||||||
| R. geographicum (L.) DC. | + | + | + | + | ||||||||||||||
| R. inarense (Vain.) Vain. | 1.0 ± 0.6 | + | 1.0 ± 0.6 | + | + | 1.0 ± 0.3 | + | 1.0 ± 0.5 | ||||||||||
| Rhizoplaca melanophthalma (DC.) Leuckert & Poelt | + | + | + | + | + | + | + | + | ||||||||||
| Rinodina mniaroea (Ach.) Körb. | + | + | + | + | ||||||||||||||
| R. olivaceobrunnea C. W. Dodge & G. E. Baker | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||
| R. terrestris Tomin | + | + | + | + | ||||||||||||||
| R. turfacea (Wahlenb.) Körb. | + | + | + | + | 1.0 ± 0.6 | + | + | + | + | + | + | + | 1.4 ± 0.9 | + | + | + | ||
| Rostania ceranisca (Nyl.) Otálora et al. | + | + | + | + | + | + | + | + | + | |||||||||
| Sporastatia testudinea (Ach.) A. Massal. | + | + | ||||||||||||||||
| Tetramelas insignis (Nägeli ex Hepp) Kalb | + | + | + | + | 1.0 ± 0.4 | + | + | + | + | 1.0 ± 0.3 | + | + | + | |||||
| T. geophilus (Flörke ex Sommerf.) Norman | + | + | + | + | + | |||||||||||||
| T. papillatus (Sommerf.) Kalb | 2.0 ± 0.9 | 2.0 ± 0.8 | 2.0 ± 0.9 | 2.0 ± 0.9 | ||||||||||||||
| Tremolecia atrata (Ach.) Hertel | 1.0 ± 0.7 | + | 1.0 ± 0.6 | + | + | + | + | 1.0 ± 0.4 | + | 1.0 ± 0.8 | + | |||||||
| Number of crustose species | 34 | 32 | 18 | 7 | 34 | 17 | 29 | 11 | 17 | 30 | 30 | 16 | 14 | 18 | 12 | 19 | 26 | 31 |
| Foliose lichens | ||||||||||||||||||
| Arctocetraria nigricascens (Nyl.) Kärnefelt & A. Thell | 1.0 ± 0.6 | + | 1.0 ± 0.4 | 1.0 ± 0.6 | + | 1.0 ± 0.6 | + | |||||||||||
| Cetraria ericetorum Opiz | + | 1.0 ± 0.6 | + | 1.3 ± 0.8 | 1.0 ± 0.6 | + | 1.0 ± 0.6 | + | + | + | 1.3 ± 1.0 | |||||||
| C. islandica (L.) Ach. | 2.2 ± 0.3 | 2.3 ± 0.5 | 2.0 ± 1.2 | 1.9 ± 0.8 | 2.1 ± 0.4 | 1.1 ± 0.5 | 2.4 ± 0.5 | 3.2 ± 1.1 | 2.2 ± 0.9 | 2.6 ± 0.5 | 2.1 ± 0.3 | 1.7 ± 0.5 | 1.6 ± 0.6 | + | 3.3 ± 1.8 | 1.6 ± 0.6 | 1.8 ± 1.2 | 1.9 ± 0.3 |
| Cetrariella delisei (Bory ex Schaer.) Kärnefelt & A. Thell | 6.5 ± 1.6 | 3.8 ± 1.7 | 1.0 ± 0.6 | + | 3.0 ± 1.5 | 2.3 ± 1.1 | 8.8 ± 2.8 | 5.5 ± 2.1 | 3.1 ± 1.1 | 6.5 ± 2.0 | 5.0 ± 1.7 | 2.5 ± 1.3 | 1.4 ± 0.7 | 1.0 ± 0.3 | 6.5 ± 3.7 | 1.3 ± 0.6 | 2.7 ± 1.7 | 4.9 ± 2.2 |
| C. fastigiata (Delise ex Nyl.) Kärnefelt & A. Thell | + | + | ||||||||||||||||
| Flavocetraria cucullata (Bellardi) Kärnefelt & A. Thell | 3.7 ± 0.6 | 2.8 ± 0.4 | 5.0 ± 3.1 | 5.3 ± 1.1 | 3.7 ± 0.7 | 1.7 ± 1.0 | 3.8 ± 0.7 | 3.4 ± 1.2 | 2.1 ± 0.9 | 3.3 ± 0.6 | 3.8 ± 0.7 | 2.1 ± 0.6 | 4.5 ± 2.7 | 3.0 ± 0.7 | 8.8 ± 3.3 | 2.3 ± 1.1 | 2.1 ± 1.3 | 3.4 ± 0.6 |
| Foveolaria nivalis (L.) S. Chesnokov et al. | 3.5 ± 1.5 | 2.6 ± 1.2 | 1.3 ± 0.9 | 3.3 ± 1.5 | 3.0 ± 2.2 | 2.6 ± 1.6 | 4.2 ± 2.2 | 1.8 ± 1.2 | 3.0 ± 1.2 | 3.9 ± 2.4 | 2.3 ± 1.5 | 1.5 ± 0.7 | 1.3 ± 1.0 | 2.9 ± 1.6 | 3.2 ± 2.0 | 1.0 ± 0.6 | 1.0 ± 0.4 | |
| Melanelia hepatizon (Ach.) A. Thell | 1.2 ± 0.3 | 1.0 ± 0.2 | 2.8 ± 2.2 | 1.3 ± 0.8 | 1.1 ± 0.3 | + | 1.6 ± 0.5 | 1.2 ± 0.4 | + | 1.4 ± 0.4 | 1.1 ± 0.3 | 1.3 ± 0.8 | + | + | 1.0 ± 0.7 | + | + | 1.3 ± 0.4 |
| M. stygia (L.) Essl. | 2.8 ± 1.6 | + | + | + | 4.0 ± 1.0 | 2.8 ± 1.6 | + | + | ||||||||||
| Parmelia omphalodes (L.) Ach. | + | + | + | + | + | + | + | + | + | |||||||||
| P. saxatilis (L.) Ach. | + | + | + | + | + | + | + | + | ||||||||||
| P. skultii Hale | 1.1 ± 0.4 | + | + | 1.0 ± 0.4 | + | 1.5 ± 0.7 | + | 1.9 ± 1.2 | + | 1.4 ± 0.6 | + | 1.0 ± 0.6 | + | + | + | + | + | + |
| Peltigera aphthosa (L.) Willd. | + | + | + | 1.0 ± 0.6 | + | + | 1.0 ± 0.5 | + | + | 1.0 ± 0.6 | 1.0 ± 0.6 | + | ||||||
| P. canina (L.) Willd. | + | + | + | + | + | + | + | + | + | + | + | |||||||
| P. elisabethae Gyeln. | + | + | + | |||||||||||||||
| P. leucophlebia (Nyl.) Gyeln. | + | + | + | + | + | + | + | + | 2.0 ± 1.3 | + | + | + | + | + | ||||
| P. malacea (Ach.) Funck | + | + | + | + | + | + | + | |||||||||||
| P. polydactylon (Neck.) Hoffm. | + | + | + | |||||||||||||||
| P. ponojensis Gyeln. | + | 1.0 ± 0.3 | + | + | 1.0 ± 0.6 | + | + | + | + | + | + | + | ||||||
| P. rufescens (Weiss) Humb. | + | + | + | + | + | + | + | + | + | + | ||||||||
| Physcia caesia (Hoffm.) Fürnr. | + | + | + | |||||||||||||||
| Physconia muscigena (Ach.) Poelt | + | 1.0 ± 0.6 | + | + | 1.0 ± 0.6 | + | + | + | 1.0 ± 0.5 | + | + | 1.0 ± 0.5 | ||||||
| Rusavskia elegans (Link) S. Y. Kondr. & Kärnefelt | + | + | + | + | ||||||||||||||
| Solorina bispora Nyl. | 1.5 ± 0.7 | 2.0 ± 0.9 | 1.0 ± 0.6 | 1.5 ± 0.7 | 2.0 ± 1.4 | 1.0 ± 0.7 | ||||||||||||
| S. crocea (L.) Ach. | + | 2.0 ± 1.4 | + | + | 1.3 ± 0.6 | + | 1.3 ± 0.7 | 1.0 ± 0.6 | + | 1.4 ± 0.6 | ||||||||
| S. saccata (L.) Ach. | 1.0 ± 0.7 | 1.0 ± 0.5 | 1.0 ± 0,4 | 1.0 ± 0.7 | ||||||||||||||
| Umbilicaria aprina Nyl. | + | + | + | + | + | + | ||||||||||||
| U. arctica (Ach.) Nyl. | 15.3 ± 10.8 | 30.0 ± 12.9 | + | + | 3.0 ± 1.8 | 12.0 ± 8.9 | ||||||||||||
| U. cylindrica (L.) Delise ex Duby | 15.0 ± 10.3 | + | 15.0 ± 7.4 | + | 7.8 ± 5.2 | + | + | |||||||||||
| U. decussata (Vill.) Zahlbr. | 1.6 ± 0.8 | + | 1.3 ± 1.0 | 1.8 ± 0.7 | + | + | 1.0 ± 0.6 | 1.6 ± 0.8 | + | + | 1.8 ± 0.9 | |||||||
| U. hyperborea (Ach.) Hoffm. | 2.8 ± 1.3 | 3.7 ± 2.7 | 1.3 ± 0.8 | 2.7 ± 1.2 | 1.1 ± 0.3 | + | 5.9 ± 2.1 | + | 3.9 ± 1.5 | + | + | 1.0 ± 0.6 | + | + | 1.0 ± 0.6 | |||
| U. cf. lyngei Schol. | + | + | + | + | ||||||||||||||
| U. proboscidea (L.) Schrad. | 1.6 ± 0.8 | 2.3 ± 1.2 | 1.2 ± 0.7 | + | + | 2.8 ± 1.0 | 2.4 ± 1.2 | 1.8 ± 1.3 | + | + | + | 1.3 ± 0.8 | ||||||
| U. torrefacta (Lightf.) Schrad. | 8.5 ± 5.2 | 1.0 ± 0.5 | + | 8.0 ± 4.2 | + | 2.0 ± 0.9 | 1.3 ± 1.0 | 8.0 ± 6.9 | + | 8.0 ± 4.8 | 2.0 ± 1.6 | |||||||
| U. virginis Schaer. | + | + | + | + | + | + | + | + | + | + | ||||||||
| Number of foliose species | 28 | 21 | 19 | 11 | 30 | 19 | 25 | 11 | 10 | 27 | 26 | 9 | 13 | 20 | 13 | 16 | 23 | 24 |
| Fruticose lichens | ||||||||||||||||||
| Alectoria ochroleuca (Hoffm.) A. Massal. | 2.1 ± 1.1 | 1.1 ± 0.3 | 1.0 ± 0.4 | 1.3 ± 0.8 | + | + | 2.7 ± 1.4 | 2.0 ± 1.2 | 2.6 ± 1.0 | + | + | + | + | + | ||||
| A. nigricans (Ach.) Nyl. | 2.4 ± 0.5 | 1.4 ± 0.3 | 3.1 ± 1.7 | 2.2 ± 1.7 | 2.1 ± 0.6 | 1.2 ± 0.7 | 2.3 ± 0.6 | 3.1 ± 1.0 | 1.4 ± 0.8 | 2.4 ± 0.5 | 1.7 ± 0.3 | 1.0 ± 0.4 | + | 4.8 ± 3.2 | 2.5 ± 1.8 | 1.3 ± 0.6 | 3.2 ± 1.6 | 1.5 ± 0.4 |
| Bryocaulon divergens (Ach.) Kärnefelt | 2.9 ± 0.7 | 2.2 ± 0.5 | 3.6 ± 1.3 | 4.1 ± 2.7 | 3.1 ± 0.7 | 1.0 ± 0.4 | 2.6 ± 0.7 | 2.5 ± 1.7 | 1.7 ± 1.0 | 2.8 ± 1.0 | 2.7 ± 0.5 | 1.4 ± 0.7 | 1.0 ± 0.6 | 5.0 ± 2.9 | 2.0 ± 0.7 | 1.3 ± 0.6 | 4.3 ± 2.7 | 2.5 ± 0.6 |
| Bryoria chalybeiformis (L.) Brodo & D. Hawksw. | + | + | + | + | + | + | + | + | + | + | ||||||||
| Cetraria aculeata (Schreb.) Fr. | 1.0 ± 0.7 | + | 2.0 ± 1.2 | 1.0 ± 0.5 | + | 2.0 ± 1.2 | + | 1.0 ± 0.5 | + | |||||||||
| C. muricata (Ach.) Eckfeldt | 1.9 ± 1.0 | + | + | 1.8 ± 1.1 | + | 1.8 ± 1.3 | 1.5 ± 0.8 | 2.0 ± 1.4 | + | 5.0 ± 2.6 | + | + | ||||||
| Cladonia amaurocraea (Flörke) Schaer. | 1.0 ± 0.4 | 1.0 ± 0.7 | 1.0 ± 0.4 | 1.0 ± 0.6 | 1.0 ± 0.4 | 1.0 ± 0.5 | ||||||||||||
| C. borealis S. Stenroos | + | + | + | + | + | + | + | + | ||||||||||
| C. carneola (Fr.) Fr. | + | + | + | + | + | |||||||||||||
| C. chlorophaea (Flörke ex Sommerf.) Spreng. | 1.0 ± 0.4 | 1.0 ± 0.7 | 1.1 ± 0.3 | + | 1.2 ± 0.5 | + | 1.0 ± 0.7 | 1.0 ± 0.3 | ||||||||||
| C. coccifera (L.) Willd. | 1.5 ± 0.7 | 2.5 ± 0.7 | 1.5 ± 0.5 | 2.5 ± 0.5 | 2.3 ± 0.4 | 1.0 ± 0.7 | 2.0 ± 1.2 | 2.0 ± 0.7 | ||||||||||
| C. gracilis (L.) Willd. | 4.6 ± 3.2 | + | + | + | 3.5 ± 2.3 | + | 3.4 ± 2.4 | + | + | + | 1.0 ± 0.7 | + | ||||||
| C. macroceras (Delise) Hav. | 1.3 ± 0.9 | 1.3 ± 0.7 | + | 2.0 ± 0.9 | 1.3 ± 0.9 | |||||||||||||
| C. phyllophora Hoffm. | 1.0 ± 0.7 | + | 1.0 ± 0.6 | + | + | 1.0 ± 0.7 | + | 1.0 ± 0.6 | ||||||||||
| C. pleurota (Flörke) Schaer. | + | + | + | + | + | + | + | |||||||||||
| C. pocillum (Ach.) Grognot | + | + | 1.0 ± 0.5 | + | + | + | 1.2 ± 0.8 | 1.0 ± 0.6 | + | + | 1.6 ± 0.9 | + | ||||||
| C. pyxidata (L.) Hoffm. | 1.5 ± 0.4 | + | + | + | 1.0 ± 0.2 | + | 1.7 ± 0.7 | + | 1.5 ± 0.8 | 1.0 ± 0.2 | 1.4 ± 0.5 | 1.0 ± 0.6 | + | + | 4.3 ± 2.3 | + | + | 1.1 ± 0.3 |
| C. stricta (Nyl.) Nyl. | 1.9 ± 1.5 | + | + | + | 6.0 ± 3.8 | 1.0 ± 0.6 | 1.3 ± 0.7 | + | + | 1.3 ± 1.0 | ||||||||
| Pseudephebe minuscula (Nyl. ex Arnold) Brodo & D. Hawksw. | 17.7 ± 12.4 | + | 25.0 ± 11.8 | + | 1.8 ± 1.3 | + | 2.8 ± 1.6 | + | 12.8 ± 9.9 | + | 12.0 ± 6.2 | |||||||
| P. pubescens (L.) M. Choisy | 1.8 ± 0.6 | + | 2.8 ± 1.6 | 1.3 ± 0.5 | + | 1.9 ± 0.7 | 2.8 ± 1.7 | + | 2.0 ± 0.6 | 1.0 ± 0.6 | + | + | + | + | 1.6 ± 1.1 | |||
| Sphaerophorus fragilis (L.) Pers. | 5.7 ± 3.2 | 1.0 ± 0.6 | + | + | 6.6 ± 3.2 | + | 3.4 ± 2.0 | 2.0 ± 1.2 | + | 5.7 ± 3.2 | + | + | + | 1.3 ± 1.0 | + | 1.3 ± 0.9 | + | |
| S. globosus (Huds.) Vain. | 2.9 ± 0.8 | 2.8 ± 1.5 | + | 2.7 ± 1.4 | + | 3.9 ± 0.8 | 1.3 ± 1.1 | + | 3.1 ± 0.9 | 1.7 ± 0.7 | + | + | 2.2 ± 1.7 | + | 2.0 ± 1.2 | |||
| Stereocaulon alpinum Laurer | 2.4 ± 0.6 | 5.3 ± 2.7 | + | 2.0 ± 1.3 | 1.2 ± 0.8 | 1.9 ± 0.8 | 2.1 ± 0.5 | 7.5 ± 4.9 | 11.5 ± 2.7 | 3.7 ± 2.5 | 2.7 ± 0.7 | 1.8 ± 1.3 | 11.0 ± 7.7 | 1.6 ± 0.9 | 4.0 ± 2.5 | 7.9 ± 5.2 | 2.1 ± 1.0 | 3.5 ± 1.0 |
| S. botryosum Ach. | 3.1 ± 1.6 | 4.8 ± 1.8 | 5.0 ± 2.7 | 2.5 ± 1.3 | 4.6 ± 3.5 | 8.0 ± 3.9 | 2.0 ± 1.3 | 5.0 ± 1.3 | 6.0 ± 2.2 | 3.5 ± 1.9 | 2.5 ± 0.7 | 1.4 ± 0.7 | 1.2 ± 0.8 | 2.0 ± 1.2 | 3.5 ± 2.5 | |||
| S. condensatum Hoffm. | 2.0 ± 0.8 | 1.0 ± 0.4 | 2.0 ± 1.4 | 1.0 ± 0.8 | 1.0 ± 0.7 | 1.5 ± 0.3 | ||||||||||||
| S. depressum (Frey) I. M. Lamb | 1.0 ± 0.3 | 1.0 ± 0.4 | 1.0 ± 0.6 | 1.0 ± 0.6 | 1.0 ± 0.8 | 1.0 ± 0.3 | ||||||||||||
| S. glareosum (Savicz) H. Magn. | 2.3 ± 1.1 | 4.0 ± 2.3 | + | 2.3 ± 1.5 | ||||||||||||||
| Thamnolia vermicularis (Sw.) Schaer. | 2.0 ± 0.3 | 2.0 ± 0.3 | 2.3 ± 0.7 | 3.2 ± 1.8 | 1.9 ± 0.3 | 1.3 ± 0.3 | 2.6 ± 0.5 | 2.8 ± 1.0 | 1.5 ± 0.6 | 2.3 ± 0.4 | 2.0 ± 0.3 | 2.1 ± 0.6 | 1.4 ± 0.8 | 1.4 ± 0.6 | 2.5 ± 1.4 | 1.8 ± 0.5 | 1.6 ± 0.4 | 2.1 ± 0.3 |
| Usnea sphacelata R. Br. | + | + | + | + | + | + | + | + | + | + | + | + | ||||||
| Number of fruticose species | 26 | 24 | 14 | 7 | 26 | 16 | 25 | 16 | 14 | 24 | 26 | 12 | 11 | 14 | 14 | 15 | 21 | 20 |
| Components | Eigenvalue | % of Total Variance | Cumulative Eigenvalue | Cumulative, % |
|---|---|---|---|---|
| Crustose lichens | ||||
| 1 | 6.8074 | 13.0911 | 6.8074 | 13.0911 |
| 2 | 4.0709 | 7.8286 | 10.8782 | 20.9197 |
| 3 | 3.0108 | 5.7901 | 13.8891 | 26.7098 |
| 4 | 2.8326 | 5.4473 | 16.7216 | 32.1570 |
| 5 | 2.6450 | 5.0861 | 19.3667 | 37.2436 |
| 6 | 2.3047 | 4.4321 | 21.6714 | 41.6757 |
| 7 | 2.0824 | 4.0045 | 23.7537 | 45.6802 |
| 8 | 1.9932 | 3.8331 | 25.7470 | 49.5134 |
| 9 | 1.6285 | 3.1317 | 27.3755 | 52.6451 |
| 10 | 1.5972 | 3.0714 | 28.9726 | 55.7166 |
| Fruticose lichens | ||||
| 1 | 3.5761 | 12.7720 | 3.5761 | 12.7720 |
| 2 | 2.3682 | 8.4578 | 5.9443 | 21.2298 |
| 3 | 1.9041 | 6.8004 | 7.8484 | 28.0302 |
| 4 | 1.5328 | 5.4741 | 9.3812 | 33.5043 |
| 5 | 1.4521 | 5.1860 | 10.8333 | 38.6903 |
| 6 | 1.3175 | 4.7052 | 12.1508 | 43.3956 |
| 7 | 1.2920 | 4.6142 | 13.4427 | 48.0097 |
| 8 | 1.1600 | 4.1427 | 14.6027 | 52.1525 |
| 9 | 1.1200 | 4.0001 | 15.7227 | 56.1524 |
| 10 | 1.0510 | 3.7537 | 16.7737 | 59.9062 |
| Orographic Factor | b* | s* | b | s | t | p |
|---|---|---|---|---|---|---|
| Crustose lichens (R2 = 0.3425) | ||||||
| Altitude | 0.5691 | 0.1489 | 0.0399 | 0.0127 | 3.1514 | 0.0024 |
| Distance to the glacier | 0.4655 | 0.1233 | 0.1285 | 0.0381 | 3.3696 | 0.0013 |
| Slope angle | −0.3766 | 0.1508 | −0.0395 | 0.0158 | −2.4976 | 0.0150 |
| Foliose lichens (R2 = 0.2218) | ||||||
| Slope angle | −0.4323 | 0.1421 | −0.0229 | 0.0085 | −2.6911 | 0.0089 |
| Fruticose lichens (R2 = 0.3876) | ||||||
| Exposure | 0.4974 | 0.1297 | 0.1974 | 0.0678 | 2.9109 | 0.0048 |
| Slope angle | 0.4131 | 0.1442 | 0.0256 | 0.0118 | 2.1709 | 0.0333 |
| Altitude | −0.3990 | 0.1423 | −0.0228 | 0.0109 | −2.1009 | 0.0392 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kholod, S.; Konoreva, L.; Chesnokov, S. Influence of Orographic Factors on the Distribution of Lichens in the Franz Josef Land Archipelago. Plants 2024, 13, 193. https://doi.org/10.3390/plants13020193
Kholod S, Konoreva L, Chesnokov S. Influence of Orographic Factors on the Distribution of Lichens in the Franz Josef Land Archipelago. Plants. 2024; 13(2):193. https://doi.org/10.3390/plants13020193
Chicago/Turabian StyleKholod, Sergey, Liudmila Konoreva, and Sergey Chesnokov. 2024. "Influence of Orographic Factors on the Distribution of Lichens in the Franz Josef Land Archipelago" Plants 13, no. 2: 193. https://doi.org/10.3390/plants13020193