
Uncovering Novel QTLs and Candidate Genes for Salt Tolerance at the Bud Burst Stage in Rice through Genome-Wide Association Study

 and
and
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Variation in the 211 Rice Varieties
2.2. Identification of QTLs for STB via GWAS
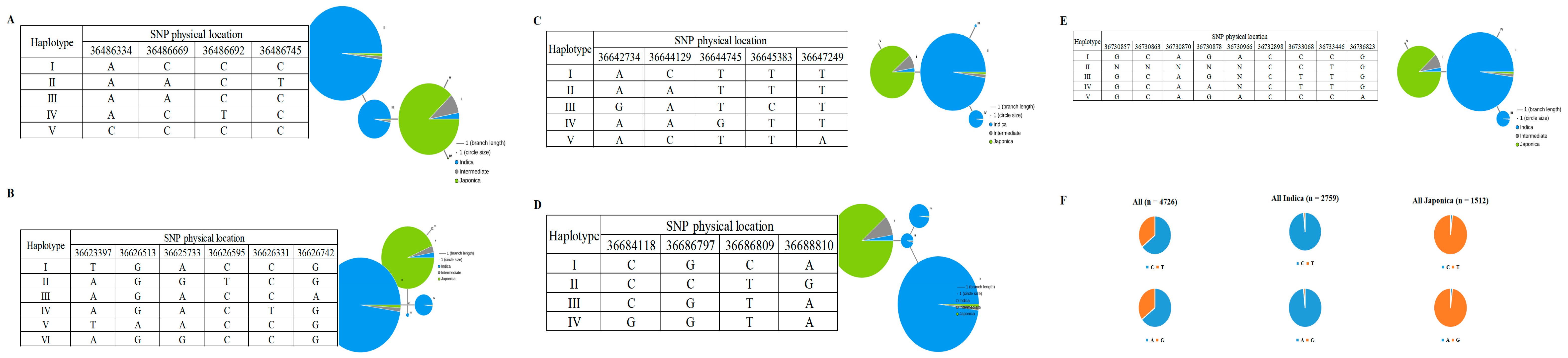
2.3. Candidate Genes Screen in qRSL1-2
2.4. Natural Allelic Variation in Candidate Genes Contributes to RSL
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Salt Tolerance Evaluation at the Bud Burst Stage
4.3. Genome-Wide Association Study
4.4. Candidate Gene Analyses
4.5. Genomic DNA Extraction, Sequencing and Genotyping
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mather, K.A.; Caicedo, A.L.; Polato, N.R.; Olsen, K.M.; McCouch, S.; Purugganan, M.D. The extent of linkage disequilibrium in rice (Oryza sativa L.). Genetics 2007, 177, 2223–2232. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Takehisa, H.; Shimodate, T.; Fukuta, Y.; Ueda, T.; Yano, M.; Yamaya, T.; Kameya, T.; Sato, T. Identification of quantitative trait loci for plant growth of rice in paddy field flooded with salt water. Field Crop. Res. 2004, 89, 85–95. [Google Scholar] [CrossRef]
- Yu, J.; Zhao, W.; Tong, W.; He, Q.; Yoon, M.Y.; Li, F.P.; Choi, B.; Heo, E.B.; Kim, K.W.; Park, Y.J. A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage. Int. J. Mol. Sci. 2018, 19, 3145. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R.; Hyman, J.R.; Lin, G.Y. Relationships between cold- and salt-tolerance during seed germination in tomato: Analysis of response and correlated response to selection. Plant Breed. 1999, 118, 49–52. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mendioro, M.S.; Diaz, G.Q.; Gregorio, G.B.; Singh, R.K. Genetic analysis of salt tolerance at seedling and reproductive stages in rice (Oryza sativa). Plant Breed. 2014, 133, 548–559. [Google Scholar] [CrossRef]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, X.H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef]
- Lei, L.; Zheng, H.; Bi, Y.; Yang, L.; Liu, H.; Wang, J.; Sun, J.; Zhao, H.; Li, X.; Li, J.; et al. Identification of a Major QTL and Candidate Gene Analysis of Salt Tolerance at the Bud Burst Stage in Rice (Oryza sativa L.) Using QTL-Seq and RNA-Seq. Rice 2020, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Kota, S.; Flowers, T.J. Salt tolerance in rice: Seedling and reproductive stage QTL mapping come of age. Theor. Appl. Genet. 2021, 134, 3495–3533. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, J.; Bao, Y.; Wu, Y.; Zhang, H. Quantitative trait loci controlling rice seed germination under salt stress. Euphytica 2011, 178, 297–307. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, B.; Zhao, H.; Wang, J.; Liu, H.; Sun, J.; Xing, J.; Zou, D. Identification of QTLs for salt tolerance at the germination and early seedling stage using linkage and association analysis in japonica rice. Chin. J. Rice Sci. 2014, 28, 358–366. [Google Scholar]
- Mardani, Z.; Rabiei, B.; Sabouri, H.; Sabouri, A. Identification of molecular markers linked to salt-tolerant genes at germination stage of rice. Plant Breed. 2014, 133, 196–202. [Google Scholar] [CrossRef]
- Luo, X.; Deng, H.; Wang, P.; Zhang, X.; Li, C.; Li, C.; Tan, J.; Wu, G.; Wang, Y.; Cheng, Q.; et al. Genetic analysis of germinating ability under alkaline and neutral salt stress by a high-density bin genetic map in rice. Euphytica 2020, 216, 107. [Google Scholar] [CrossRef]
- Cheng, J.; He, Y.; Yang, B.; Lai, Y.; Wang, Z.; Zhang, H. Association mapping of seed germination and seedling growth at three conditions in indica rice (Oryza sativa L.). Euphytica 2015, 206, 103–115. [Google Scholar] [CrossRef]
- Shi, Y.; Gao, L.; Wu, Z.; Zhang, X.; Wang, M.; Zhang, C.; Zhang, F.; Zhou, Y.; Li, Z. Genome-wide association study of salt tolerance at the seed germination stage in rice. BMC Plant Biol. 2017, 17, 92. [Google Scholar] [CrossRef] [PubMed]
- Lekklar, C.; Pongpanich, M.; Suriya-arunroj, D.; Chinpongpanich, A.; Tsai, H.; Comai, L.; Chadchawan, S.; Buaboocha, T. Genome-wide association study for salinity tolerance at the flowering stage in a panel of rice accessions from Thailand. BMC Genom. 2019, 20, 76. [Google Scholar] [CrossRef] [PubMed]
- Warraich, A.S.; Krishnamurthy, S.L.; Sooch, B.S.; Vinaykumar, N.M.; Dushyanthkumar, B.M.; Bose, J.; Sharma, P.C. Rice GWAS reveals key genomic regions essential for salinity tolerance at reproductive stage. Acta Physiol. Plant. 2020, 42, 134. [Google Scholar] [CrossRef]
- Islam, M.R.; Naveed, S.A.; Zhang, Y.; Li, Z.; Zhao, X.; Fiaz, S.; Zhang, F.; Wu, Z.; Hu, Z.; Fu, B.; et al. Identification of Candidate Genes for Salinity and Anaerobic Tolerance at the Germination Stage in Rice by Genome-Wide Association Analyses. Front. Genet. 2022, 13, 822516. [Google Scholar] [CrossRef]
- Zou, J.; Liu, C.; Liu, A.; Zou, D.; Chen, X. Overexpression of OsHsp17.0 and OsHsp23.7 enhances drought and salt tolerance in rice. J. Plant Physiol. 2012, 169, 628–635. [Google Scholar] [CrossRef]
- Chen, H.C.; Cheng, W.H.; Hong, C.Y.; Chang, Y.S.; Chang, M.C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA-dependent and ABA-independent pathways, respectively. Rice 2018, 11, 50. [Google Scholar] [CrossRef]
- He, Y.; Yang, B.; He, Y.; Zhan, C.; Cheng, Y.; Zhang, J.; Zhang, H.; Cheng, J.; Wang, Z. A quantitative trait locus, qSE3, promotes seed germination and seedling establishment under salinity stress in rice. Plant J. 2019, 97, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. OsNAC45 is Involved in ABA Response and Salt Tolerance in Rice. Rice 2020, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Shen, L.; Shen, Z.; Jing, W.; Ge, H.; Zhao, J.; Zhang, W. The potassium transporter OsHAK21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Tamiru, M.; Abe, A.; Yoshida, K.; Uemura, A.; Yaegashi, H.; Obara, T.; Oikawa, K.; Utsushi, H.; Kanzaki, E.; et al. MutMap accelerates breeding of a salt-tolerant rice cultivar. Nat. Biotechnol. 2015, 33, 445–449. [Google Scholar] [CrossRef]
- Huang, X.Y.; Chao, D.Y.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K. A major gene for low temperature germinability in rice (Oryza sativa L.). Euphytica 2004, 136, 63–68. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, S.; Zhao, Q.; Zhou, L.; Zhao, L.; Yao, S.; Zhang, Y.; Wang, C. Mapping of QTLs for Bud-stage Salinity Tolerance Based on Chromosome Segment Substitution Line in Rice. Acta Agric. Boreali-Sin. 2017, 32, 106–111. [Google Scholar]
- Lei, L.; Han, Z.; Cui, B.; Yang, L.; Liu, H.; Wang, J.; Zhao, H.; Xin, W.; Li, X.; Li, J.; et al. Mapping of a major QTL for salinity tolerance at the bud burst stage in rice (Oryza sativa L) using a high-density genetic map. Euphytica 2021, 217, 167. [Google Scholar] [CrossRef]
- Nan, N.; Wang, J.; Shi, Y.; Qian, Y.; Jiang, L.; Huang, S.; Liu, Y.; Wu, Y.; Liu, B.; Xu, Z.Y. Rice plastidial NAD-dependent malate dehydrogenase 1 negatively regulates salt stress response by reducing the vitamin B6 content. Plant Biotechnol. J. 2019, 18, 172–184. [Google Scholar] [CrossRef]
- Le, T.D.; Gathignol, F.; Vu, H.T.; Nguyen, K.L.; Tran, L.H.; Vu, H.T.T.; Dinh, T.X.; Lazennec, F.; Pham, X.H.; Véry, A.; et al. Genome-Wide Association Mapping of Salinity Tolerance at the Seedling Stage in a Panel of Vietnamese Landraces Reveals New Valuable QTLs for Salinity Stress Tolerance Breeding in Rice. Plants 2021, 10, 1088. [Google Scholar] [CrossRef]
- Fang, H.; Meng, Q.; Xu, J.; Tang, H.; Tang, S.; Zhang, H.; Huang, J. Knock-down of stress inducible OsSRFP1 encoding an E3 ubiquitin ligase with transcriptional activation activity confers abiotic stress tolerance through enhancing antioxidant protection in rice. Plant Mol. Biol. 2015, 87, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, K.; Izui, K. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 2000, 23, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Z.; Xiao, Y.; Liu, X.; Chen, Y.; Zhang, Z.; Kang, H.; Wang, X.; Jiang, S.; Peng, S.; et al. Association Mapping and Functional Analysis of Rice Cold Tolerance QTLs at the Bud Burst Stage. Rice 2021, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhu, W.; Zhang, Y.; Zhu, Z.; Zhao, L.; Chen, T.; Zhao, Q.; Zhou, L.; Fang, X.; Wang, Y.; et al. Detection of QTL for Cold Tolerance at Bud Bursting Stage Using Chromosome Segment Substitution Lines in Rice (Oryza sativa). Rice Sci. 2011, 18, 71–74. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, H.; Zhang, D.; Li, J.; Xiong, H.; Yu, J.; Li, J.; Rashid, M.A.R.; Li, G.; Ma, X.; et al. Genetic Analysis of Cold Tolerance at the Germination and Booting Stages in Rice by Association Mapping. PLoS ONE 2015, 10, e120590. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lei, L.; Li, P.; Wang, J.; Wang, C.; Yang, F.; Chen, J.; Liu, H.; Zheng, H.; Xin, W.; et al. Identification of Candidate Genes Conferring Cold Tolerance to Rice (Oryza sativa L.) at the Bud-Bursting Stage Using Bulk Segregant Analysis Sequencing and Linkage Mapping. Front. Plant Sci. 2021, 12, 647239. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, D.; Liu, H.; Liu, Y.; Huang, M.; Wang, H.; Chen, Z.; Guo, T. Identification of QTLs involved in cold tolerance during the germination and bud stages of rice (Oryza sativa L.) via a high-density genetic map. Breed. Sci. 2020, 70, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ye, J.; Xu, Q.; Feng, Y.; Yuan, X.; Yu, H.; Wang, Y.; Wei, X.; Yang, Y. Genome-wide association study of cold tolerance of Chinese indica rice varieties at the bud burst stage. Plant Cell Rep. 2018, 37, 529–539. [Google Scholar] [CrossRef]
- Li, C.; Liu, J.; Bian, J.; Jin, T.; Zou, B.; Liu, S.; Zhang, X.; Wang, P.; Tan, J.; Wu, G.; et al. Identification of cold tolerance QTLs at the bud burst stage in 211 rice landraces by GWAS. BMC Plant Biol. 2021, 21, 542. [Google Scholar] [CrossRef]
- Li, X.; Zheng, H.; Wu, W.; Liu, H.; Wang, J.; Jia, Y.; Li, J.; Yang, L.; Lei, L.; Zou, D.; et al. QTL Mapping and Candidate Gene Analysis for Alkali Tolerance in Japonica Rice at the bud Stage Based on Linkage Mapping and Genome-Wide Association Study. Rice 2020, 13, 48. [Google Scholar] [CrossRef]
- Chung, P.J.; Jung, H.; Choi, Y.D.; Kim, J. Genome-wide analyses of direct target genes of four rice NAC-domain transcription factors involved in drought tolerance. BMC Genom. 2018, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Woo, Y.; Ryu, S.; Shin, Y.; Kim, W.T.; Park, K.Y.; Lee, I.; An, G. Further Characterization of a Rice AGL12 Group MADS-Box Gene, OsMADS26. Plant Physiol. 2008, 147, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Yuenyong, W.; Chinpongpanich, A.; Comai, L.; Chadchawan, S.; Buaboocha, T. Downstream components of the calmodulin signaling pathway in the rice salt stress response revealed by transcriptome profiling and target identification. BMC Plant Biol. 2018, 18, 335. [Google Scholar] [CrossRef] [PubMed]
- Vij, S.; Giri, J.; Dansana, P.K.; Kapoor, S.; Tyagi, A.K. The Receptor-Like Cytoplasmic Kinase (OsRLCK) Gene Family in Rice: Organization, Phylogenetic Relationship, and Expression during Development and Stress. Mol. Plant 2008, 1, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, H.; Long, Y.; Shu, Y.; Zhai, J. Plant Public RNA-seq Database: A comprehensive online database for expression analysis of ~45 000 plant public RNA-Seq libraries. Plant Biotechnol. J. 2022, 20, 806–808. [Google Scholar] [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; Xing, Y.; Chen, L.; Xie, W. RiceVarMap: A comprehensive database of rice genomic variations. Nucleic Acids Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | 211 Accessions | 130 indica Accessions | 81 japonica Accessions | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | CV | Mean | Range | CV | Mean | Range | CV | |
| RSL | 0.415 ± 0.078 | 0.231–0.696 | 18.7% | 0.443 ± 0.078 | 0.231–0.696 | 17.8% | 0.371 ± 0.051 | 0.288–0.578 | 13.9% |
| RRL | 0.276 ± 0.141 | 0.035–0.910 | 51.0% | 0.251 ± 0.131 | 0.071–0.775 | 52.1% | 0.316 ± 0.147 | 0.035–0.910 | 46.5% |
| RRN | 0.584 ± 0.097 | 0.318–0.860 | 16.6% | 0.580 ± 0.107 | 0.318–0.860 | 18.4% | 0.591 ± 0.080 | 0.388–0.782 | 13.5% |
| Trait | RSL | RRL | RRN |
|---|---|---|---|
| RSL | 1 | 0.267 ** | 0.432 ** |
| RRL | 0.267 ** | 1 | 0.204 ** |
| RRN | 0.432 ** | 0.204 ** | 1 |
| QTL ID | Trait | Chr. | Peak SNPs | p Value | R2 | Previous QTL/Genes |
|---|---|---|---|---|---|---|
| qRSL1-1 | RSL | 1 | 35691376 | 3.20 × 10−4 | 0.091007605 | OsMDH1 |
| qRSL1-2 | RSL | 1 | 36789943 | 2.65 × 10−4 | 0.082702247 | |
| qRSL1-3 | RSL | 1 | 36988603 | 7.26 × 10−4 | 0.072572863 | |
| qRSL3 | RSL | 3 | 13014108 | 1.13 × 10−4 | 0.092421651 | OsSRFP1 |
| qRSL4 | RSL | 4 | 29820476 | 4.53 × 10−4 | 0.077107238 | OsCDPK7 |
| qRRL4 | RRL | 4 | 5864214 | 5.18 × 10−4 | 0.076378954 | |
| qRRL5 | RRL | 5 | 18874396 | 6.00 × 10−4 | 0.074780842 | |
| qRRN1 | RRN | 1 | 32234318 | 5.06 × 10−4 | 0.060777188 | QTL_2 |
| qRRN4 | RRN | 4 | 29820476 | 1.15 × 10−4 | 0.092366087 | OsCDPK7 |
| Gene ID | Leaves-20 Days | Post-Emergence Inflorescence | Pre-Emergence Inflorescence | Anther | Pistil | Seed-5 DAP | Embryo-25 DAP | Endosperm-25 DAP | Seed-10 DAP | Endosperm- 25 DAP (Replicate) | Leaves-20 days (Replicate) | Shoots |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LOC_Os01g62950 | 2.25939 | 10.0462 | 5.71393 | 216.92 | 8.45013 | 9.64699 | 2.03138 | 4.22441 | 3.58312 | 3.50467 | 3.7757 | 25.0827 |
| LOC_Os01g62980 | 8.97772 | 55.7145 | 17.5485 | 1.64909 | 39.8088 | 5.31876 | 1.02588 | 0.550286 | 0.329246 | 0.332092 | 11.3404 | 45.3305 |
| LOC_Os01g63060 | 7.51192 | 17.6248 | 11.2939 | 16.5555 | 40.2667 | 16.0646 | 8.30178 | 2.95839 | 4.79123 | 3.31343 | 7.28362 | 94.1526 |
| LOC_Os01g63190 | 58.0209 | 39.3723 | 0.776449 | 0.709715 | 0 | 10.945 | 0 | 0 | 0.444664 | 0 | 11.7839 | 11.9054 |
| LOC_Os01g63230 | 13.7457 | 6.46319 | 1.00233 | 5.34358 | 1.40058 | 0.672047 | 1.80477 | 13.6685 | 5.3863 | 13.6636 | 11.384 | 12.8333 |
| LOC_Os01g63280 | 5.23959 | 1.04796 | 0.869788 | 1.10318 | 1.08601 | 0.832929 | 0 | 0 | 0 | 0 | 4.495 | 7.77339 |
| LOC_Os01g63400 | 4.35858 | 5.53168 | 7.76593 | 0.69816 | 4.32159 | 1.941 | 3.98117 | 0.544953 | 0.73063 | 0.598556 | 1.50912 | 6.61778 |
| LOC_Os01g63460 | 5.14934 | 7.66854 | 15.1861 | 1.18708 | 7.16601 | 4.26658 | 1.06111 | 0.32016 | 0 | 0 | 2.64417 | 6.98541 |
| LOC_Os01g63540 | 0 | 0 | 3.28173 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2.85494 |
| LOC_Os01g63580 | 0.42975 | 78.9638 | 117.25 | 11.0498 | 44.6477 | 77.7596 | 6.86909 | 3.76347 | 3.26988 | 3.11506 | 1.19418 | 25.4433 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Lu, C.; Yang, M.; Wu, G.; Nyasulu, M.; He, H.; He, X.; Bian, J. Uncovering Novel QTLs and Candidate Genes for Salt Tolerance at the Bud Burst Stage in Rice through Genome-Wide Association Study. Plants 2024, 13, 174. https://doi.org/10.3390/plants13020174
Li C, Lu C, Yang M, Wu G, Nyasulu M, He H, He X, Bian J. Uncovering Novel QTLs and Candidate Genes for Salt Tolerance at the Bud Burst Stage in Rice through Genome-Wide Association Study. Plants. 2024; 13(2):174. https://doi.org/10.3390/plants13020174
Chicago/Turabian StyleLi, Caijing, Changsheng Lu, Mengmeng Yang, Guangliang Wu, Mvuyeni Nyasulu, Haohua He, Xiaopeng He, and Jianmin Bian. 2024. "Uncovering Novel QTLs and Candidate Genes for Salt Tolerance at the Bud Burst Stage in Rice through Genome-Wide Association Study" Plants 13, no. 2: 174. https://doi.org/10.3390/plants13020174