
Conserving Marine Forests: Assessing the Effectiveness of a Marine Protected Area for Cystoseira sensu lato Populations in the Central Mediterranean Sea

Abstract
:1. Introduction
2. Results
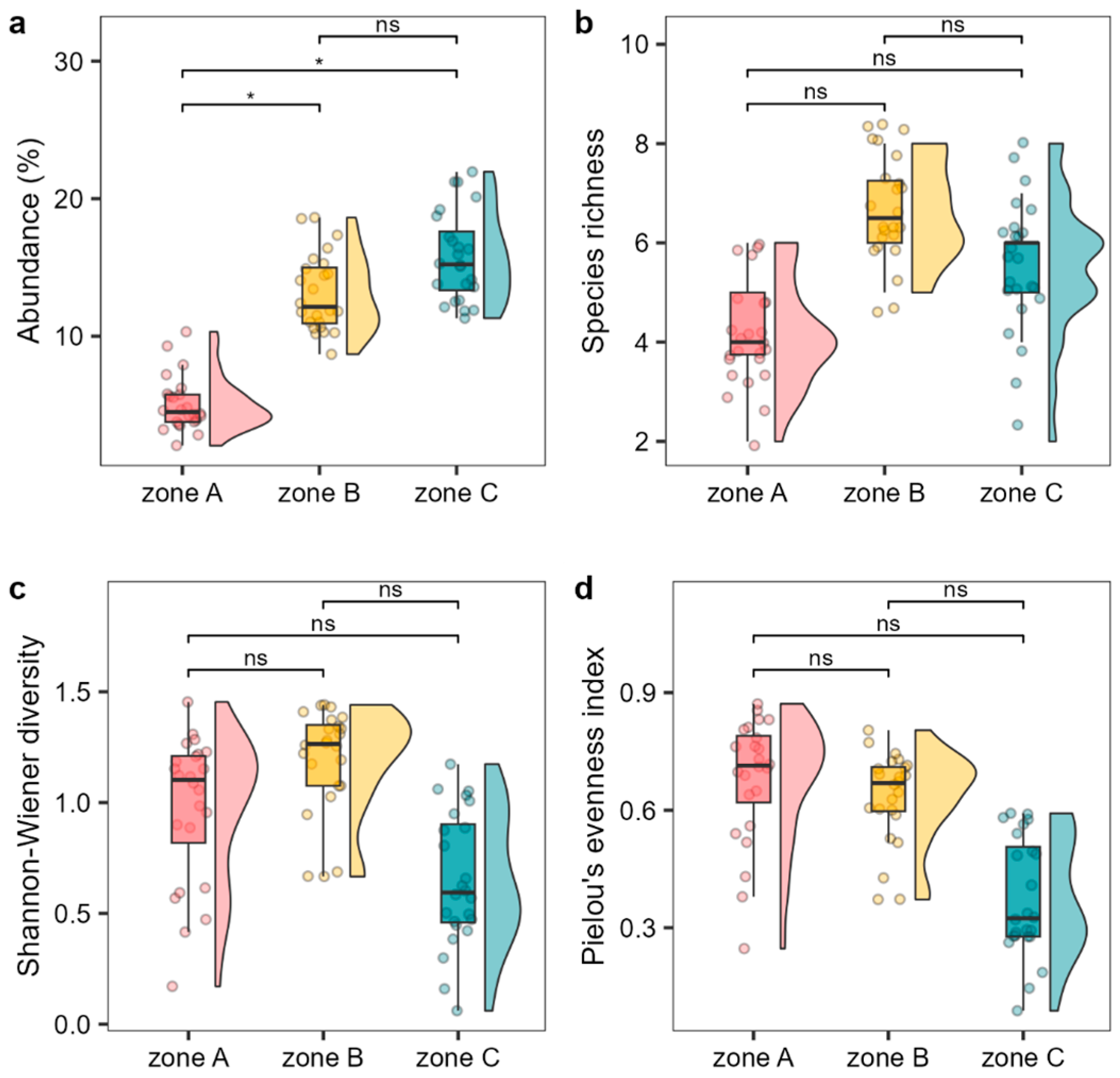
2.1. Cystoseira s.l. Assemblage within the MPA
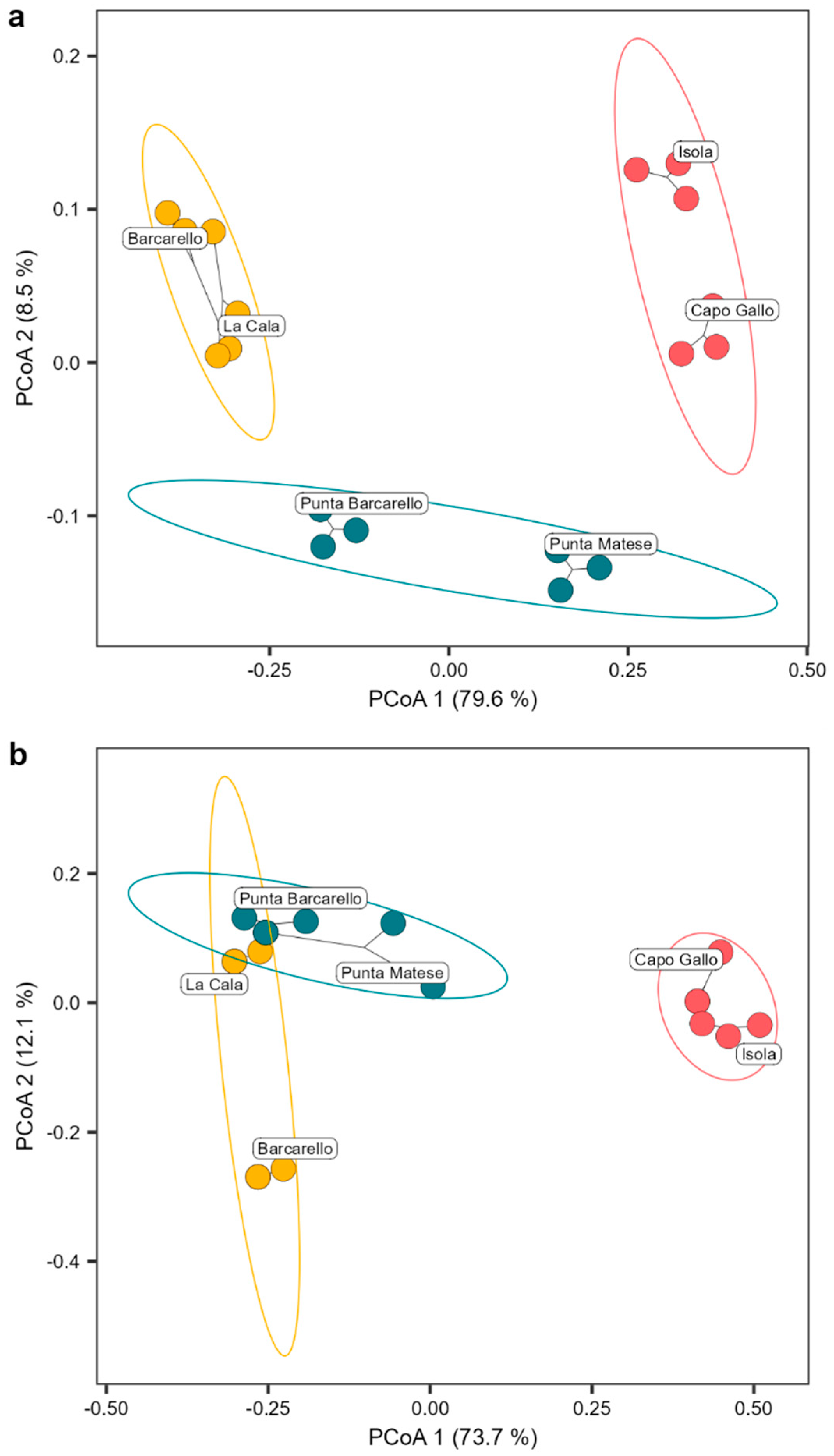
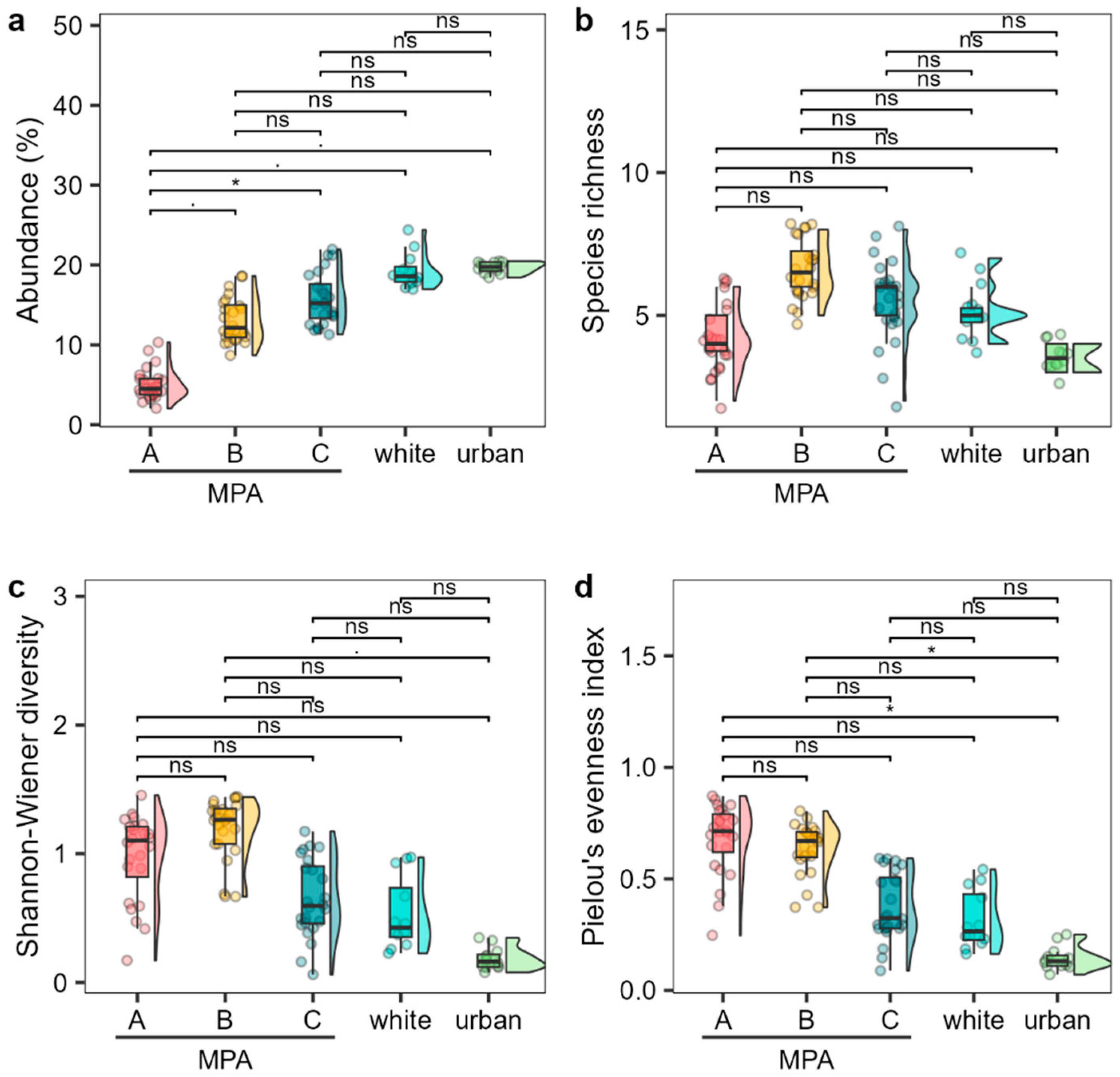
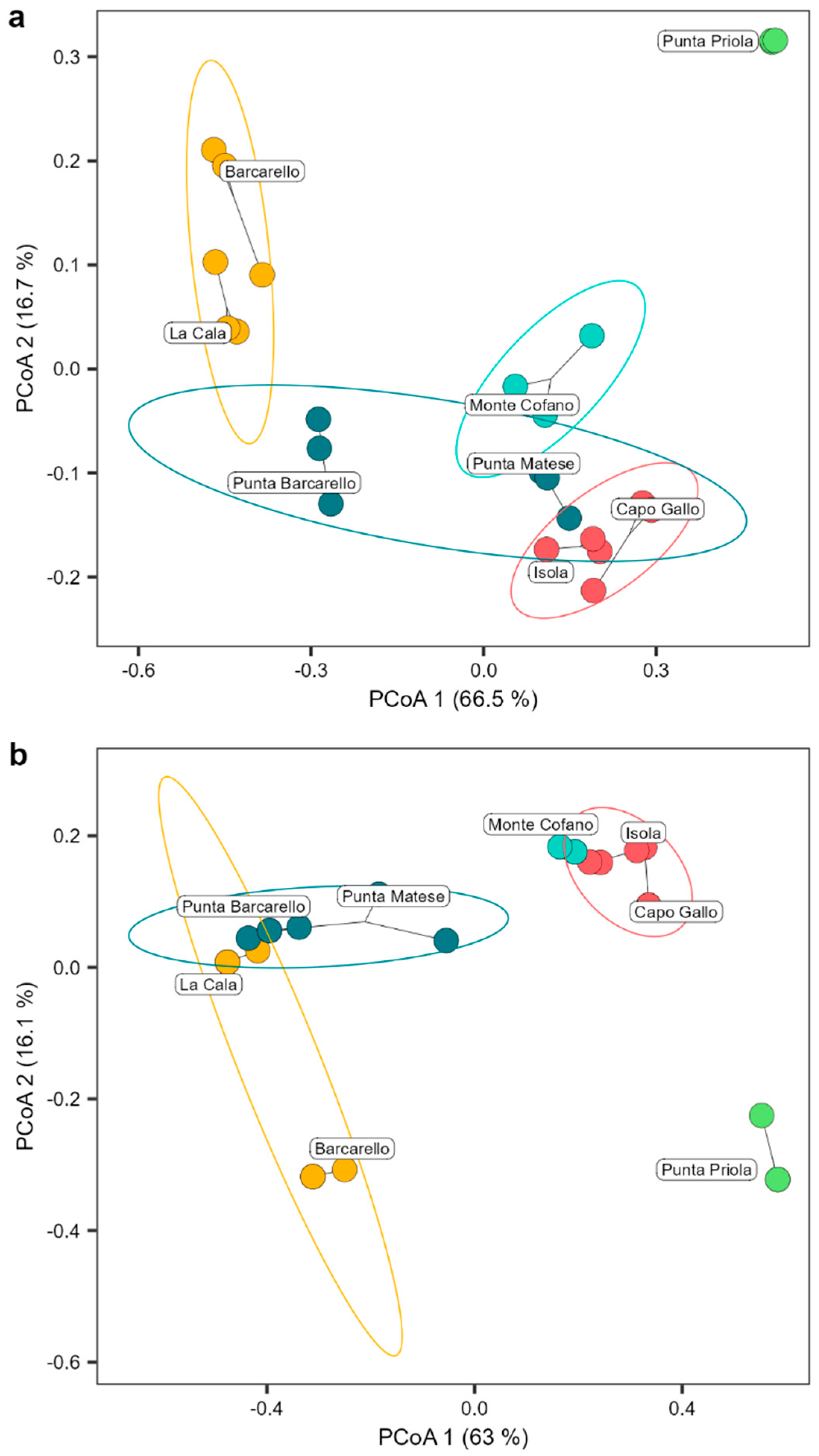
2.2. Comparison of Cystoseira s.l. Assemblage between MPA and Unprotected Sites
2.3. Non-Indigenous Seaweeds
3. Discussion
4. Materials and Methods
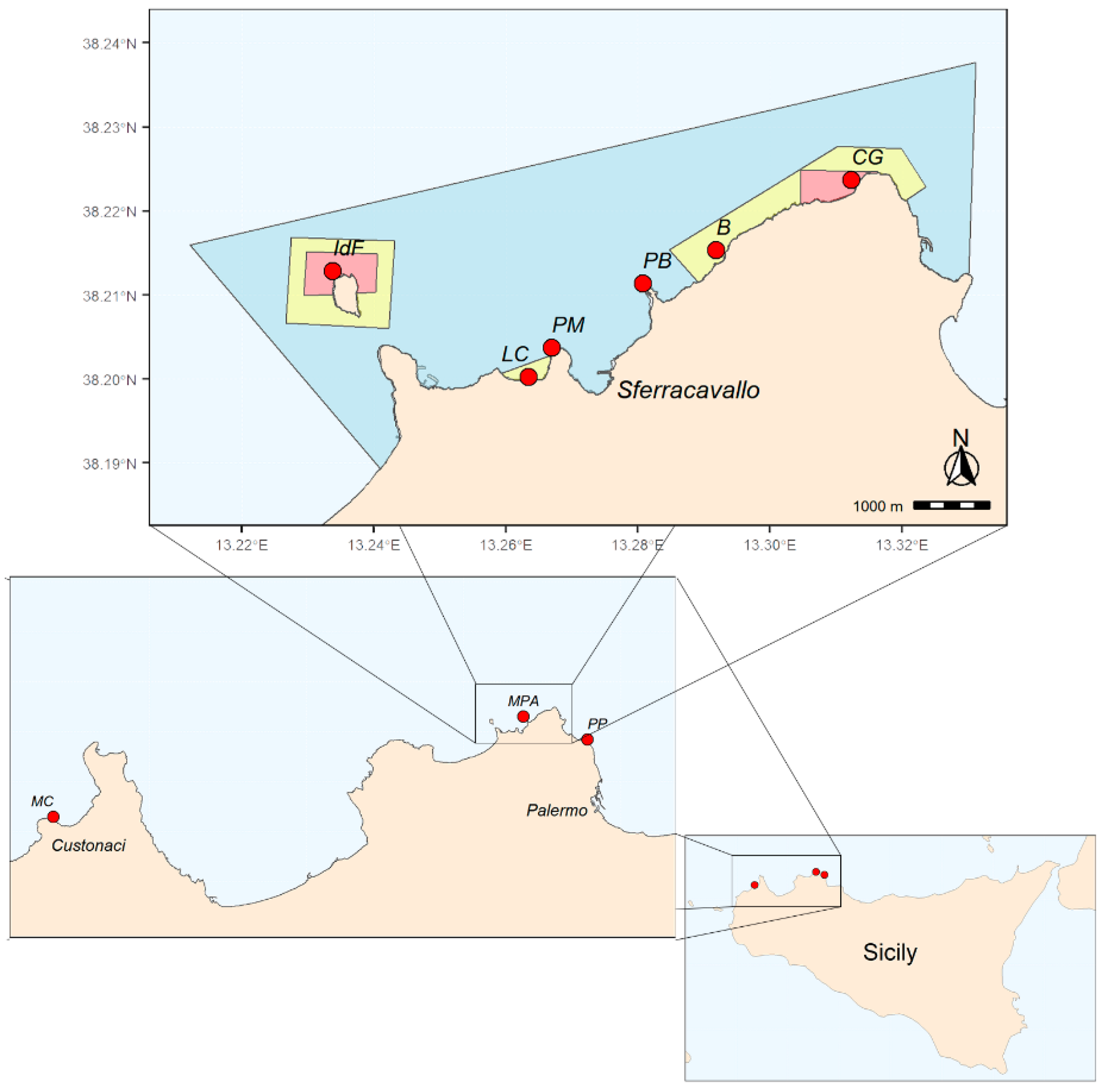
4.1. The “Capo Gallo-Isola delle Femmine” MPA
4.2. Unprotected Sites
4.3. Survey of Cystoseira s.l. Populations
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global Patterns of Kelp Forest Change over the Past Half-Century. Proc. Natl. Acad. Sci. USA 2016, 113, 13785–13790. [Google Scholar] [CrossRef] [PubMed]
- Mineur, F.; Arenas, F.; Assis, J.; Davies, A.J.; Engelen, A.H.; Fernandes, F.; Malta, E.; Thibaut, T.; Van Nguyen, T.; Vaz-Pinto, F.; et al. European Seaweeds under Pressure: Consequences for Communities and Ecosystem Functioning. J. Sea Res. 2015, 98, 91–108. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp Forest Ecosystems: Biodiversity, Stability, Resilience and Future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Coleman, M.A.; Wernberg, T. Forgotten Underwater Forests: The Key Role of Fucoids on Australian Temperate Reefs. Ecol. Evol. 2017, 7, 8406–8418. [Google Scholar] [CrossRef] [PubMed]
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira Sensu Lato (Fucales, Phaeophyceae) in the Eastern Atlantic and Mediterranean Based on Morphological and DNA Evidence, Including Carpodesmia Gen. Emend. and Treptacantha Gen. Emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Molinari Novoa, E.A.; Guiry, M.D. Reinstatement of the Genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Not. Algarum 2020, 172, 1–10. [Google Scholar]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.M.; Francour, P. Nursery Value of Cystoseira Forests for Mediterranean Rocky Reef Fishes. J. Exp. Mar. Bio. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Schiel, D.R.; Foster, M.S. The Population Biology of Large Brown Seaweeds: Ecological Consequences of Multiphase Life Histories in Dynamic Coastal Environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Giaccone, G.; Alongi, G.; Pizzuto, F.; Cossu, A.V.L. La Vegetazione Marina Bentonica Fotofila Del Mediterraneo: 2. Infralitorale e Circalitorale: Proposte Di Aggiornamento. Boll. dell’Accad. Gioenia di Sci. Nat. 1994, 27, 111–157. [Google Scholar]
- Falace, A.; Bressan, G. Seasonal Variations of Cystoseira Barbata (Stackhouse) C. Agardh Frond Architecture. Hydrobiologia 2006, 555, 193–206. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Acunto, S.; Cinelli, F.; Hawkins, S.J. The Influence of Canopy Algae on Vertical Patterns of Distribution of Low-Shore Assemblages on Rocky Coasts in the Northwest Mediterranean. J. Exp. Mar. Bio. Ecol. 2002, 267, 89–106. [Google Scholar] [CrossRef]
- Ballesteros, E.; Garrabou, J.; Hereu, B.; Zabala, M.; Cebrian, E.; Sala, E. Deep-Water Stands of Cystoseira Zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: Insights into Assemblage Structure and Population Dynamics. Estuar. Coast. Shelf Sci. 2009, 82, 477–484. [Google Scholar] [CrossRef]
- European Commission Water Framework Directive (2000/60/EC); European Environment Agency: Copenhagen, Denmark, 2000; Volume L 269, pp. 1–15.
- MSFD. Directive 2008/56/EC of the European Parliament and the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive). 2008. Available online: http://ec.europa.eu/environment/water/marine/index_en.htm (accessed on 23 December 2023).
- Khailov, K.M.; Firsov, Y.K. The Relationships between Weight, Length, Age and Intensity of Photosynthesis and Organotrophy in the Thallus of Cystoseira Barbata from the Black Sea. Bot. Mar. 1976, 19, 329–334. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Pannacciulli, F.; Bulleri, F.; Moschella, P.S.; Airoldi, L.; Relini, G.; Cinelli, F. Predicting the Consequences of Anthropogenic Disturbance: Large-Scale Effects of Loss of Canopy Algae on Rocky Shores. Mar. Ecol. Prog. Ser. 2001, 214, 137–150. [Google Scholar] [CrossRef]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-Term Decline of the Populations of Fucales (Cystoseira Spp. and Sargassum Spp.) in the Albères Coast (France, North-Western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef] [PubMed]
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Loss of Fucoid Algae along a Gradient of Urbanisation, and Structure of Benthic Assemblages. Mar. Ecol. Prog. Ser. 2008, 358, 63–74. [Google Scholar] [CrossRef]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The Ups and Downs of a Canopy-Forming Seaweed over a Span of More than One Century. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of Vulnerable Cystoseira Populations along the Italian Infralittoral Fringe, and Relationships with Environmental and Anthropogenic Variables. Mar. Pollut. Bull. 2018, 129, 762–771. [Google Scholar] [CrossRef]
- Strain, E.M.A.; Thomson, R.J.; Micheli, F.; Mancuso, F.P.; Airoldi, L. Identifying the Interacting Roles of Stressors in Driving the Global Loss of Canopy-Forming to Mat-Forming Algae in Marine Ecosystems. Glob. Chang. Biol. 2014, 20, 3300–3312. [Google Scholar] [CrossRef]
- Arevalo, R.; Pinedo, S.; Ballesteros, E.; Arévalo, R.; Pinedo, S.; Ballesteros, E. Changes in the Composition and Structure of Mediterranean Rocky-Shore Communities Following a Gradient of Nutrient Enrichment: Descriptive Study and Test of Proposed Methods to Assess Water Quality Regarding Macroalgae. Mar. Pollut. Bull. 2007, 55, 104–113. [Google Scholar] [CrossRef]
- Falace, A.; Alongi, G.; Cormaci, M.; Furnari, G.; Curiel, D.; Cecere, E.; Petrocelli, A. Changes in the Benthic Algae along the Adriatic Sea in the Last Three Decades. Chem. Ecol. 2010, 26, 77–90. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfune, A.; Boudouresque, C.F.; Verlaque, M. Decline and Local Extinction of Fucales in French Riviera: The Harbinger of Future Extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Connell, S.D.; Foster, M.S.; Airoldi, L. What Are Algal Turfs? Towards a Better Description of Turfs. Mar. Ecol. Prog. Ser. 2014, 495, 299–307. [Google Scholar] [CrossRef]
- Agnetta, D.; Badalamenti, F.; Ceccherelli, G.; Di Trapani, F.; Bonaviri, C.; Gianguzza, P. Role of Two Co-Occurring Mediterranean Sea Urchins in the Formation of Barren from Cystoseira Canopy. Estuar. Coast. Shelf Sci. 2015, 152, 73–77. [Google Scholar] [CrossRef]
- Buonomo, R.; Chefaoui, R.M.; Lacida, R.B.; Engelen, A.H.; Serrão, E.A.; Airoldi, L. Predicted Extinction of Unique Genetic Diversity in Marine Forests of Cystoseira spp. Mar. Environ. Res. 2018, 138, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.E.; Halpern, B.S.; Grorud-Colvert, K.; Lubchenco, J.; Ruttenberg, B.I.; Gaines, S.D.; Airamé, S.; Warner, R.R. Biological Effects within No-Take Marine Reserves: A Global Synthesis. Mar. Ecol. Prog. Ser. 2009, 384, 33–46. [Google Scholar] [CrossRef]
- Gianni, F.; Bartolini, F.; Airoldi, L.; Ballesteros, E.; Francour, P.; Guidetti, P.; Meinesz, A.; Thibaut, T.; Mangialajo, L. Conservation and Restoration of Marine Forests in the Mediterranean Sea and the Potential Role of Marine Protected Areas. Adv. Oceanogr. Limnol. 2013, 4, 83–101. [Google Scholar] [CrossRef]
- Blowes, S.A.; Chase, J.M.; Di Franco, A.; Frid, O.; Gotelli, N.J.; Guidetti, P.; Knight, T.M.; May, F.; McGlinn, D.J.; Micheli, F.; et al. Mediterranean Marine Protected Areas Have Higher Biodiversity via Increased Evenness, Not Abundance. J. Appl. Ecol. 2020, 57, 578–589. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Cottalorda, J.M.; Hereu, B.; Susini, M.L.; Verlaque, M. Unexpected Temporal Stability of Cystoseira and Sargassum Forests in Port-Cros, One of the Oldest Mediterranean Marine National Parks. Cryptogam. Algol. 2016, 37, 61–90. [Google Scholar] [CrossRef]
- Sala, E.; Ballesteros, E.; Dendrinos, P.; Di Franco, A.; Ferretti, F.; Foley, D.; Fraschetti, S.; Friedlander, A.; Garrabou, J.; Güçlüsoy, H.; et al. The Structure of Mediterranean Rocky Reef Ecosystems across Environmental and Human Gradients, and Conservation Implications. PLoS ONE 2012, 7, e32742. [Google Scholar] [CrossRef]
- Falace, A.; Kaleb, S.; De La Fuente, G.; Asnaghi, V.; Chiantore, M. Ex Situ Cultivation Protocol for Cystoseira Amentacea Var. Stricta (Fucales, Phaeophyceae) from a Restoration Perspective. PLoS ONE 2018, 13, e0193011. [Google Scholar] [CrossRef] [PubMed]
- Verdura, J.; Sales, M.; Ballesteros, E.; Cefalì, M.E.; Cebrian, E. Restoration of a Canopy-Forming Alga Based on Recruitment Enhancement: Methods and Long-Term Success Assessment. Front. Plant Sci. 2018, 9, 1832. [Google Scholar] [CrossRef] [PubMed]
- Decreto Ministeriale 24 Luglio 2002, Istituzione dell’Area Marina Protetta denominata Capo Gallo—Isola delle Femmine; Governo Italiano: Romoe, Italy, 2002.
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 22 August 2022).
- Meinesz, A.; Blanfuné, A. 1983–2013: Development of Marine Protected Areas along the French Mediterranean Coasts and Perspectives for Achievement of the Aichi Target. Mar. Policy 2015, 54, 10–16. [Google Scholar] [CrossRef]
- Riggio, S.; Raimondo, F.M. Proposta Di Una Riserva Costiera per La Tutela e La Valorizzazione Dei Biotopi Di Isola Delle Femmine e Di Monte Gallo (Palermo). Quad. di Bot. Appl. 1991, 2, 59–96. [Google Scholar]
- Lucido, M. Geomorfologia Della Piattaforma Continentale Tra Torre Del Pozzillo e Torre Mondello (Sicilia Nord-Occidentale). Nat. Sicil. 1992, 16, 91–107. [Google Scholar]
- Ramos, E.; Díaz de Terán, J.R.; Puente, A.; Juanes, J.A. The Role of Geomorphology in the Distribution of Intertidal Rocky Macroalgae in the NE Atlantic Region. Estuar. Coast. Shelf Sci. 2016, 179, 90–98. [Google Scholar] [CrossRef]
- Tsiamis, K.; Salomidi, M.; Gerakaris, V.; Mogg, A.O.M.; Porter, E.S.; Sayer, M.D.J.; Küpper, F.C. Macroalgal Vegetation on a North European Artificial Reef (Loch Linnhe, Scotland): Biodiversity, Community Types and Role of Abiotic Factors. J. Appl. Phycol. 2020, 32, 1353–1363. [Google Scholar] [CrossRef]
- Muth, A.F. Effects of Zoospore Aggregation and Substrate Rugosity on Kelp Recruitment Success. J. Phycol. 2012, 48, 1374–1379. [Google Scholar] [CrossRef]
- Gaylord, B.; Reed, D.C.; Washburn, L.; Raimondi, P.T. Physical–Biological Coupling in Spore Dispersal of Kelp Forest Macroalgae. J. Mar. Syst. 2004, 49, 19–39. [Google Scholar] [CrossRef]
- Giaccone, G.; Sortino, M. Flora e Vegetazione Algale Di Isola Delle Femmine. Lav. dell’Istit. Bot. Giard. Colon. Palermo 1964, 21, 140–164. [Google Scholar]
- Giaccone, G.; Colonna, P.; Graziano, C.; Mannino, A.M.; Tornatore, E.; Cormaci, M.; Furnari, G.; Scammacca, B. Revisione Della Flora Marina Di Sicilia e Isole Minori. Boll. dell’Accad. Gioenia di Sci. Nat. 1985, 18, 537–781. [Google Scholar]
- Mannino, A.M. Studio Fitosociologico Della Vegetazione Mesolitorale a Lithophyllum Lichenoides Philippi (Rhodophyceae, Corallinales). Nat. Sicil. 1992, 16, 3–25. [Google Scholar]
- Mannino, A.M.; Mancuso, F.P. Guida All’identificazione Delle Cistoseire (Area Marina Protetta “Capo Gallo-Isola Delle Femmine”); Università degli Studi di Palermo: Palermo, Italy, 2009. [Google Scholar]
- Perkol-Finkel, S.; Airoldi, L. Loss and Recovery Potential of Marine Habitats: An Experimental Study of Factors Maintaining Resilience in Subtidal Algal Forests at the Adriatic Sea. PLoS ONE 2010, 5, e10791. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Ito, M.; Matsumoto, A.; Arakawa, H. Effects of Sediment Influx on the Settlement and Survival of Canopy-Forming Macrophytes. Sci. Rep. 2016, 6, 18677. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, L.; Virgilio, M. Responses of Turf-Forming Algae to Spatial Variations in the Deposition of Sediments. Mar. Ecol. Prog. Ser. 1998, 165, 12. [Google Scholar] [CrossRef]
- Eriksson, B.K.; Johansson, G. Effects of Sedimentation on Macroalgae: Species-Specific Responses Are Related to Reproductive Traits. Oecologia 2005, 143, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A New Methodology Based on Littoral Community Cartography Dominated by Macroalgae for the Implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef]
- Sales, M.; Cebrian, E.; Tomas, F.; Ballesteros, E. Pollution Impacts and Recovery Potential in Three Species of the Genus Cystoseira (Fucales, Heterokontophyta). Estuar. Coast. Shelf Sci. 2011, 92, 347–357. [Google Scholar] [CrossRef]
- Zenetos, A.; Galanidi, M. Mediterranean Non Indigenous Species at the Start of the 2020s: Recent Changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean—The 100 “worst Invasives” and Their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Agostaro, R.D.; Milazzo, M.; Badalamenti, F. The Invasive Seaweed Asparagopsis Taxiformis Erodes the Habitat Structure and Biodiversity of Native Algal Forests in the Mediterranean Sea. Mar. Environ. Res. 2022, 173, 105515. [Google Scholar] [CrossRef] [PubMed]
- Mannino, A.M.; Balistreri, P. Citizen Science: A Successful Tool for Monitoring Invasive Alien Species (IAS) in Marine Protected Areas. The Case Study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 2018, 8386, 42–48. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P. Invasive Alien Species in Mediterranean Marine Protected Areas: The Egadi Islands (Italy) Case Study. Biodiversity 2021, 22, 13–23. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Issaris, Y.; Poursanidis, D.; Thessalou-Legaki, M. Vulnerability of Marine Habitats to the Invasive Green Alga Caulerpa Racemosa Var. Cylindracea within a Marine Protected Area. Mar. Environ. Res. 2010, 70, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, F.P.; Chemello, R.; Mannino, A.M. The Effects of Non-Indigenous Macrophytes on Native Biodiversity: Case Studies from Sicily. J. Mar. Sci. Eng. 2023, 11, 1389. [Google Scholar] [CrossRef]
- Holmer, M.; Marbà, N.; Lamote, M.; Duarte, C.M. Deterioration of Sediment Quality in Seagrass Meadows (Posidonia Oceanica) Invaded by Macroalgae (Caulerpa Sp.). Estuaries Coasts 2009, 32, 456–466. [Google Scholar] [CrossRef]
- Arias, A.; Giangrande, A.; Gambi, M.C.; Anadón, N. Biology and New Records of the Invasive Species Branchiomma Bairdi (Annelida: Sabellidae) in the Mediterranean Sea. Mediterr. Mar. Sci. 2013, 14, 162–171. [Google Scholar] [CrossRef]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Castelluccio, F.; Pozone, F.; Roussis, V.; Templado, J.; Ghiselin, M.T.; Cimino, G. Factors Promoting Marine Invasions: A Chemoecological Approach. Proc. Natl. Acad. Sci. USA 2008, 105, 4582–4586. [Google Scholar] [CrossRef]
- Ballesteros, E.; Cebrian, E.; Alcoverro, T. Mortality of Shoots of Posidonia Oceanica Following Meadow Invasion by the Red Alga Lophocladia Lallemandii. Bot. Mar. 2007, 50, 8–13. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti-Cecchi, L.; Morri, C.; Pasini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Hard Bottoms. In Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study; Gambi, M.C., Dappiano, M., Eds.; SIBM: Genova, Italy, 2004; Volume 11, pp. 185–215. [Google Scholar]
- Dethier, M.; Graham, E.; Cohen, S.; Tear, L. Visual versus Random-Point Percent Cover Estimations: “objective” Is Not Always Better. Mar. Ecol. Prog. Ser. 1993, 96, 93–100. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Milazzo, M.; Chemello, R. Decreasing in Patch-Size of Cystoseira Forests Reduces the Diversity of Their Associated Molluscan Assemblage in Mediterranean Rocky Reefs. Estuar. Coast. Shelf Sci. 2021, 250, 107163. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988; Volume 51, ISBN 978-94-015-7360-3. [Google Scholar]
- Blondel, V.D.; Guillaume, J.L.; Lambiotte, R.; Lefebvre, E. Fast Unfolding of Communities in Large Networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; ISBN 0-12-044620-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MPA Zones | Unprotected Sites | |||||
|---|---|---|---|---|---|---|
| A | B | C | White | Urban | ||
| Species | ||||||
| Cystoseira compressa | (Esper) Gerloff & Nizamuddin 1975 | + | + | + | + | + |
| Cystoseira compressa f. rosetta | (Ercegovic) Cormaci, G. Furnari, Giaccone, B. Scammacca & Serio | + | + | |||
| Cystoseira foeniculacea | (Linnaeus) Greville | + | + | |||
| Cystoseira foeniculacea f. latiramosa | (Ercegovic) A. Gómez Garreta, M.C. Barceló, M.A. Ribera & J.R. Lluch 2001 | + | + | |||
| Cystoseira foeniculacea f. tenuiramosa | (Ercegovic) A. Gómez Garreta, M.C. Barceló, M.A. Ribera & J. Rull Lluch | + | + | |||
| Cystoseira humilis | Schousboe ex Kützing | + | + | + | ||
| Cystoseira humilis var. myriophylloides | (Sauvageau) J.H. Price & D.M. John | + | + | + | ||
| Ericaria amentacea | (C. Agardh) Molinari & Guiry | + | + | + | + | |
| Ericaria brachycarpa | (J. Agardh) Molinari & Guiry | + | + | + | + | + |
| Ericaria brachycarpa var. claudiae | Boudouresque, Perret-Boudouresque & Blanfuné | + | + | + | ||
| Ericaria crinita | (Duby) Molinari & Guiry | + | + | + | ||
| Ericaria dubia | (Valiante) Neiva & Serrão 2022 | + | + | |||
| Ericaria funkii | (Schiffner ex Gerloff & Nizamuddin) Molinari & Guiry | + | + | |||
| Ericaria mediterranea | (Sauvageau) Molinari & Guiry | + | ||||
| Gongolaria montagnei | (J. Agardh) Kuntze | + | + | + | + | |
| Gongolaria montagnei var. compressa | (Ercegovic) Verlaque, Blanfuné, Boudouresque & Thibaut | + | + | + | + | |
| Gongolaria montagnei var. tenuior | (Ercegovic) Molinari & Guiry | + | + | + | ||
| Gongolaria sauvageauana | (Hamel) Molinari & Guiry | + | ||||
| Gongolaria squarrosa | (De Notaris) Kuntze | + | ||||
| MPA Zones | Unprotected Sites | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | White | Urban | |||||||||||||
| Depth Range | Species | N | F% | D% | N | F% | D% | N | F% | D% | N | F% | D% | N | F% | D% | |
| 0–5 m | Cystoseira compressa | 3.5 | 41.7 | 3.8 | 8.3 | 50.0 | 4.7 | 5.4 | 37.5 | 5.1 | 7.9 | 50.0 | 6.7 | 11.7 | 50.0 | 51.9 | |
| Cystoseira compressa f. rosetta | 1.0 | 12.5 | 1.1 | 0.4 | 4.2 | 0.4 | |||||||||||
| Cystoseira humilis | 15.2 | 54.2 | 8.6 | 4.4 | 25.0 | 4.1 | 13.3 | 50.0 | 11.3 | ||||||||
| Ericaria amentacea | 50.8 | 100.0 | 54.5 | 82.3 | 100.0 | 46.3 | 58.1 | 100.0 | 54.8 | 60.0 | 100.0 | 50.7 | |||||
| Ericaria brachycarpa | 37.9 | 95.8 | 40.6 | 71.9 | 100.0 | 40.4 | 37.7 | 87.5 | 35.6 | 31.7 | 83.3 | 26.8 | 10.8 | 50.0 | 48.1 | ||
| Ericaria mediterranea | 5.4 | 25.0 | 4.6 | ||||||||||||||
| 5–10 m | Cystoseira humilis var. myriophylloides | 3.1 | 12.5 | 11.0 | 8.3 | 20.8 | 8.0 | 4.4 | 16.7 | 6.3 | |||||||
| Cystoseira humilis | 9.0 | 37.5 | 8.6 | 6.0 | 29.2 | 8.7 | 3.3 | 25.0 | 7.3 | ||||||||
| Cystoseira foeniculacea f. tenuiramosa | 8.1 | 37.5 | 7.8 | 7.5 | 29.2 | 10.8 | |||||||||||
| Cystoseira foeniculacea | 3.3 | 16.7 | 3.2 | 5.0 | 33.3 | 54.5 | |||||||||||
| Ericaria crinita | 19.2 | 70.8 | 67.6 | 34.8 | 79.2 | 33.5 | 23.5 | 58.3 | 34.1 | ||||||||
| Gongolaria montagnei | 6.0 | 33.3 | 21.3 | 3.1 | 16.7 | 4.5 | 6.7 | 16.7 | 14.7 | 4.2 | 33.3 | 45.5 | |||||
| Gongolaria montagnei var. tenuior | 40.2 | 83.3 | 38.8 | 24.6 | 62.5 | 35.6 | 25.0 | 75.0 | 55.0 | ||||||||
| Gongolaria sauvageauana | 10.4 | 41.7 | 22.9 | ||||||||||||||
| 10–15 m | Cystoseira foeniculacea f. tenuiramosa | 11.2 | 50.0 | 12.3 | 5.2 | 25.0 | 14.5 | ||||||||||
| Cystoseira humilis | 10.6 | 37.5 | 11.6 | ||||||||||||||
| Cystoseira foeniculacea | 5.6 | 25.0 | 6.2 | 2.1 | 8.3 | 26.3 | |||||||||||
| Ericaria crinita | 12.7 | 45.8 | 43.9 | 14.2 | 45.8 | 15.5 | 6.7 | 33.3 | 18.6 | ||||||||
| Ericaria brachycarpa var. claudiae | 1.2 | 8.3 | 4.3 | 6.0 | 16.7 | 6.6 | 4.2 | 16.7 | 52.6 | ||||||||
| Ericaria funkii | 14.0 | 45.8 | 15.3 | 6.7 | 25.0 | 18.6 | |||||||||||
| Gongolaria montagnei var. compressa | 5.4 | 25.0 | 18.7 | 4.0 | 12.5 | 4.3 | 1.9 | 12.5 | 5.2 | 0.8 | 8.3 | 3.2 | |||||
| Gongolaria montagnei var. tenuior | 25.8 | 58.3 | 28.2 | 11.2 | 50.0 | 31.4 | 10.8 | 25.0 | 41.9 | ||||||||
| 15–20 m | Cystoseira foeniculacea f. latiramosa | 7.9 | 37.5 | 11.9 | 2.7 | 20.8 | 10.2 | ||||||||||
| Cystoseira foeniculacea f. tenuiramosa | 7.7 | 33.3 | 11.6 | 1.0 | 4.2 | 3.9 | |||||||||||
| Cystoseira humilis | 3.5 | 16.7 | 5.3 | ||||||||||||||
| Ericaria dubia | 4.6 | 25.0 | 6.9 | 1.7 | 12.5 | 6.2 | |||||||||||
| Ericaria funkii | 14.8 | 37.5 | 22.3 | 4.2 | 16.7 | 15.6 | |||||||||||
| Gongolaria montagnei | 15.6 | 41.7 | 41.0 | 10.0 | 41.7 | 37.5 | 8.3 | 33.3 | 52.6 | 5.4 | 33.3 | 100.0 | |||||
| Gongolaria montagnei var. compressa | 14.8 | 70.8 | 38.8 | 5.0 | 16.7 | 7.5 | 3.5 | 20.8 | 13.3 | 4.2 | 33.3 | 26.3 | |||||
| Gongolaria squarrosa | 7.7 | 29.2 | 20.2 | ||||||||||||||
| Gongolaria montagnei var. tenuior | 22.9 | 54.2 | 34.5 | 3.5 | 12.5 | 13.3 | |||||||||||
| Gongolaria sauvageauana | 3.3 | 16.7 | 21.1 | ||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, F.P.; Sarà, G.; Mannino, A.M. Conserving Marine Forests: Assessing the Effectiveness of a Marine Protected Area for Cystoseira sensu lato Populations in the Central Mediterranean Sea. Plants 2024, 13, 162. https://doi.org/10.3390/plants13020162
Mancuso FP, Sarà G, Mannino AM. Conserving Marine Forests: Assessing the Effectiveness of a Marine Protected Area for Cystoseira sensu lato Populations in the Central Mediterranean Sea. Plants. 2024; 13(2):162. https://doi.org/10.3390/plants13020162
Chicago/Turabian StyleMancuso, Francesco Paolo, Gianluca Sarà, and Anna Maria Mannino. 2024. "Conserving Marine Forests: Assessing the Effectiveness of a Marine Protected Area for Cystoseira sensu lato Populations in the Central Mediterranean Sea" Plants 13, no. 2: 162. https://doi.org/10.3390/plants13020162