

Phytohormone Production by the Endophyte Bacillus safensis TS3 Increases Plant Yield and Alleviates Salt Stress
 , ,
, ,
Abstract
:1. Introduction
2. Results
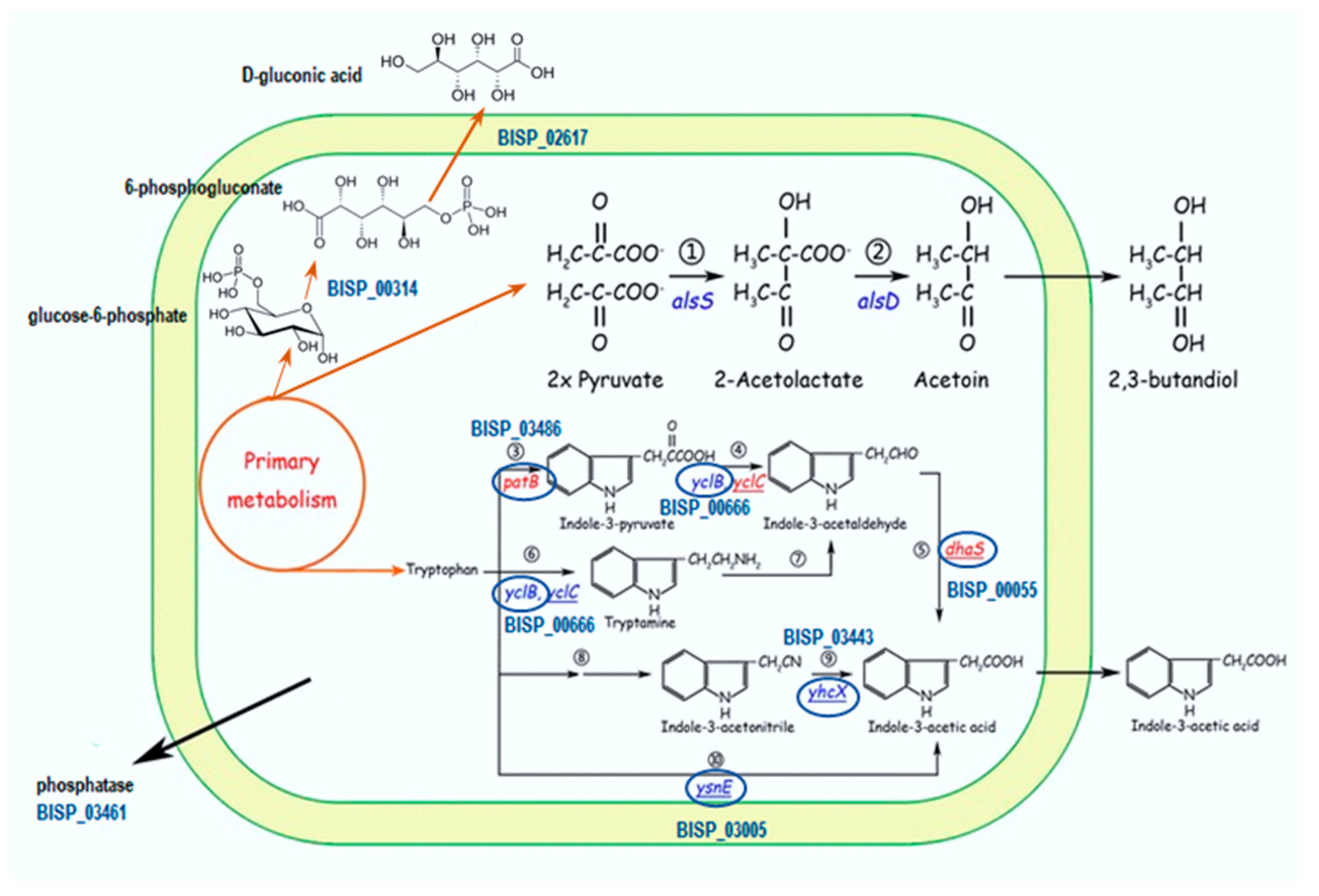
2.1. Production of Enzymes and Phytohormones by the Strain TS3
2.2. Colonization Ability of Bacillus safensis TS3
2.3. The Effect of Different Concentrations of NaCl on the Growth of TS3
2.4. The Growth-Stimulating Activity of Strain TS3 under the Condition of an Increased NaCl Concentration Using Gnotobiotic Systems
2.5. Pot Experiments with Strain TS3 on Salinated Soil
2.6. Field Tests with Potato and Cabbage Plants
2.7. Genome Analysis
3. Discussion
4. Materials and Methods
4.1. Estimation of Cellulase, Amylase, Protease, and Lipase Activity
4.2. Analysis of Bacterial Phytohormones
4.3. Determination of the Colonization Ability of Bacillus safensis TS3
4.4. Root Colonization Study by Using Fluorescence In Situ Hybridization and Confocal Laser Scanning Microscopy
4.5. The Effect of Different Concentrations of NaCl on the Growth and Cell Number of TS3
4.6. The Growth-Stimulating Activity of the TS3 Strain under the Condition of Increased NaCl Concentrations Using Gnotobiotic Systems
4.7. Pot Experiments with the Strain TS3 in Salinated Soil
4.8. Field Tests with Potato and Cabbage Plants
- Control: N100P110K130;
- N100P110K130 + preplanting treatment of tubers, with consumption of the bacterial suspension (1.0 L/t for consumption of the working solution and 10.0 L/t for spraying of the plants in the vegetative growth stage first and in the flowering stage second); consumption of bacterial suspension = 1.0 L/ha; consumption of working solution = 300.0 L/ha;
- N100P110K130 + preplanting treatment of tubers, with consumption of the bacterial suspension (1.0 L/t for consumption of the working solution and 10.0 L/t for spraying of the plants in the vegetative growth stage first and in the flowering stage second); consumption of bacterial suspension = 2.0 L/ha; consumption of working solution = 300.0 L/ha;
- N100P110K130 + preplanting treatment of tubers, with consumption of the bacterial suspension (1.0 L/t for consumption of working solution and 10.0 L/t for spraying of the plants in the vegetative growth stage first and in the flowering stage second); consumption of bacterial suspension = 3.0 L/ha; consumption of working solution = 300.0 L/ha.
- Control = N120P120K180;
- N120P120K180 + spraying of plants before planting seedlings (consumption of bacterial suspension = 1.0 L/ha, with spraying of plants first 15 days after planting seedlings and second in the phase of forming the head); consumption of bacterial suspension = 1.5 L/ha; consumption of working solution = 300 L/ha;
- N120P120K180 + spraying of plants before planting seedlings (consumption of bacterial suspension = 1.0 L/ha, with spraying of plants first 15 days after planting seedlings and second in the phase of forming the head); consumption of bacterial suspension = 3.0 L/ha; consumption of working solution = 300 L/ha;
- N120P120K180 + spraying of plants before planting seedlings (consumption of bacterial suspension = 1.0 L/ha, with spraying of plants first 15 days after planting seedlings and second in the phase of forming the head); consumption of bacterial suspension = 4.5 L/ha; consumption of working solution = 300 L/ha.
4.9. Genome Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate saltinduced damages. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M.; Ahmad, P.; Chandna, R.; Prasad, M.N.V.; Ozturk, M. Enhancing plant productivity under salt stress—Relevance of poly-omics. In Salt Stress in Plants: Omics, Signaling and Responses; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: Berlin/Heildeberg, Germany, 2013; pp. 113–156. [Google Scholar]
- Khan, M.; Rolly, N.K.; Al Azzawi, T.N.I.; Imran, M.; Mun, B.-G.; Lee, I.-J.; Yun, B.-W. Lead (Pb)-induced oxi-dative stress alters the morphological and physio-biochemical properties of rice (Oryza sativa L.). Agronomy 2021, 11, 409. [Google Scholar] [CrossRef]
- FAO. Global Network on Integrated Soil Management for Sustain-Able Use of Salt-Affected Soils. Available online: http://www.fao.org/ag/agl/agll/spush (accessed on 27 September 2023).
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef] [PubMed]
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystem. In Salinity: Environment-Plants-Molecules; Läuchli, A., Lüttge, U., Eds.; Kluwer Academic: Dodrecht, The Netherlands, 2002; p. 3. [Google Scholar]
- Choudhary, D.K.; Sharma, A.K.; Agarwal, P.; Varma, A.; Tuteja, N. Volatiles and Food Security; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Gondek, M.; Weindorf, D.C.; Thiel, C.; Kleinheinz, G. Soluble salts in compost and their effects on soil and plants: A Review. Compost Sci. Util. 2020, 28, 59–75. [Google Scholar] [CrossRef]
- Reints, J.; Dinar, A.; Crowley, D. Dealing with water scarcity and salinity: Adoption of water efficient technologies and management practices by california avocado growers. Sustainability 2020, 12, 3555. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting Rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Slama, H.B.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Baranov, O.; Ahadi, R.; Belbahri, L. Impacts of salt stress on the rhizosphere and endophytic bacterial role in plant salt alleviation. Int. J. Plant Biol. 2023, 14, 361–376. [Google Scholar] [CrossRef]
- Manishankar, P.; Wang, N.; Köster, P.; Alatar, A.A.; Kudla, J. Calcium signaling during salt stress and in the regulation of ion homeostasis. J. Exp. Bot. 2018, 69, 4215–4226. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Bhardwaj, A.K.; Mishra, V.K.; Singh, A.K.; Arora, S.; Srivastava, S.; Singh, Y.P.; Sharma, D.K. Soil salinity and land use-land cover interactions with soil carbon in a salt-affected irrigation canal command of indo-gangetic plain. Catena 2019, 180, 392–400. [Google Scholar] [CrossRef]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Wirth, S.; Egamberdieva, D. Comparing symbiotic performance and physiological responses of two soybean cultivars to arbuscular mycorrhizal fungi under salt stress. Saudi J. Biol. Sci. 2019, 26, 38–48. [Google Scholar] [CrossRef]
- Redman, R.S.; Kim, Y.O.; Woodward, C.J.; Greer, C.; Espino, L.; Doty, S.L.; Rodriguez, R.J. Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: A strategy for mitigating impacts of climate change. PLoS ONE 2011, 6, e14823. [Google Scholar] [CrossRef]
- Chebotar, V.K.; Malfanova, N.V.; Scherbakov, A.V.; Ahtemova, G.A.; Borisov, A.Y.; Lugtenberg, B.; Tikhonovich, I.A. Endophytic bacteria in microbial preparations that improve plant development. Appl. Biochem. Microbiol. 2015, 51, 271–277. [Google Scholar] [CrossRef]
- Szymańska, S.; Borruso, L.; Brusetti, L.; Hulisz, P.; Furtado, B.; Hrynkiewicz, K. Bacterial microbiome of root-associated endophytes of Salicornia europaea in correspondence to different levels of salinity. Environ. Sci. Pollut. Res. 2018, 25, 25420–25431. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Tkacz, A.; Poole, P. Role of root microbiota in plant productivity. J. Exp. Bot. 2015, 66, 2167–2175. [Google Scholar] [CrossRef]
- Vaishnav, A.; Shukla, A.K.; Sharma, A.; Kumar, R.; Choudhary, D.K. Endophytic bacteria in plant salt stress tolerance: Current and future prospects. J. Plant Growth Regul. 2019, 38, 650–668. [Google Scholar] [CrossRef]
- Naveed, M.; Hafeez, S.; Rafique, M.; Mumtaz, M.Z.; Subhani, Z.; Holatko, J.; Hammerschmiedt, T.; Malicek, O.; Mustafa, A.; Kintl, A.; et al. Plant-endophyte mediated improvement in physiological and bio-protective abilities of marigold (Tagetes patula). Front. Plant Sci. 2022, 13, 993130. [Google Scholar] [CrossRef]
- Mulholland, B.J.; Taylor, I.B.; Jackson, A.C.; Thornpson, A.J. Can ABA mediate responses of salinity stressed tomato. Environ. Exp. Bot. 2003, 50, 17–28. [Google Scholar] [CrossRef]
- Egamberdieva, D. The role of phytohormone producing bacteria in alleviating salt stress in crop plants. In Biotechnological Techniques of Stress Tolerance in Plants; Miransari, M., Ed.; Stadium Press LLC.: Cedar City, UT, USA, 2013; pp. 21–39. [Google Scholar]
- Hoffman, M.T.; Gunatilaka, M.K.; Wijeratne, K.; Gunatilaka, L.; Arnold, A.E. Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PLoS ONE 2013, 8, 31–33. [Google Scholar] [CrossRef]
- Soltani, J.; Samavati, R.; Jalili, B.; Bagheri, H.; Hamzei, J. Halotolerant endophytic bacteria from desert-adapted halophyte plants alleviate salinity stress in germinating seeds of the common wheat Triticum aestivum L. Cereal Res. Commun. 2023, 11, 1–11. [Google Scholar] [CrossRef]
- Kang, S.M.; Ul Hoque, M.I.; Woo, J.I.; Lee, I.J. Mitigation of salinity stress on soybean seedlings using indole acetic acid-producing Acinetobacter pittii YNA40. Agriculture 2023, 13, 1021. [Google Scholar] [CrossRef]
- Saleem, S.; Iqbal, A.; Ahmed, F.; Ahmad, M. Phytobeneficial and salt stress mitigating efficacy of IAA producing salt tolerant strains in Gossypium hirsutum. Saudi J. Biol. Sci. 2021, 28, 5317–5324. [Google Scholar] [CrossRef] [PubMed]
- Arif, S.; Munis, M.F.H.; Liaquat, F.; Gulzar, S.; Haroon, U.; Zhao, L.A.; Zhang, Y.D. Trichoderma viride establishes biodefense against clubroot (Plasmodiophora brassicae) and fosters plant growth via colonizing root hairs in pak choi (Brassica campestris spp. chinesnsis). Biol. Control 2023, 183, 105265. [Google Scholar] [CrossRef]
- Feng, Q.J.; Cao, S.L.; Liao, S.J.; Wassie, M.; Sun, X.Y.; Chen, L.; Xie, Y. Fusarium equiseti—Inoculation altered rhizosphere soil microbial community, potentially driving perennial ryegrass growth and salt tolerance. Sci. Total Environ. 2023, 871, 162153. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Li, Y.; Zhang, L.; Mu, J.; Jiang, Y.; Fu, H.; Zhang, Y.; Cui, H.; Yu, X.; Ye, Z. Biosynthetic pathways and functions of indole-3-acetic acid in microorganisms. Microorganisms 2023, 11, 2077. [Google Scholar] [CrossRef] [PubMed]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signaling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Medina, M.J.; Steinkellner, S.; Vieheilig, H.; Bote, J.A.O.; Garrido, J.M.G. Abscisic acid determines arbuscule development and functionality in the tomato arbuscular mycorrhiza. New Phytol. 2007, 175, 554–564. [Google Scholar] [CrossRef] [PubMed]
- De Hita, D.; Fuentes, M.; Zamarreño, A.M.; Ruiz, Y.; Garcia-Mina, J.M. Culturable bacterial endophytes from sedimentary humic acid-treated plants. Front. Plant Sci. 2020, 11, 837. [Google Scholar] [CrossRef]
- Satomi, M.; La Duc, M.T.; Venkateswaran, K. Bacillus safensis sp. nov., isolated from spacecraft and assembly-facility surfaces. Internat J. Syst. Evolut. Microbiol. 2006, 56, 1735–1740. [Google Scholar] [CrossRef]
- Kothari, V.V.; Kothari, R.K.; Kothari, C.R.; Bhatt, V.D.; Nathani, N.M.; Koringa, P.G.; Joshi, C.G.; Vyas, B.R.M. Genome sequence of salt-tolerant Bacillus safensis strain VK, isolated from saline desert area of Gujarat, India. Genome Announc. 2013, 1, e00671-13. [Google Scholar] [CrossRef]
- Raja, C.E.; Omine, K. Arsenic, boron and salt resistant Bacillus safensis MS11 isolated from Mongolia desert soil. Afr. J. Biotechnol. 2012, 11, 2267–2275. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.N.; Dey, P.L.; Chakraborty, A.P. Bacillus safensis from wheat rhizosphere promotes growth and ameliorates salinity stress in wheat. Indian J. Biotechnol. 2018, 17, 466–479. [Google Scholar]
- Chebotar, V.K.; Gancheva, M.S.; Voshol, G.P.; Malfanova, N.V.; Karasev, E.S.; Chizhevskaya, E.P.; Zaplatkin, A.N.; Khiutti, A.V.; Lazarev, A.M.; Gadjiev, N.M.; et al. Draft genome sequence of the tomato stem endophyte Bacillus safensis TS3. Microbiol. Resour. Announc. 2022, 17, e00816-22. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; van der Bij, A.J.; Brand, J.; de Weger, L.A.; Wijffelman, C.A.; Lugtenberg, B.J.J. Gnotobiotic system for studying rhizosphere colonisation by plant growth-promoting Pseudomonas bacteria. Mol. Plant-Microbe Interact. 1996, 9, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Li, S.; Zhang, N.; Cui, X.; Zhou, X.; Zhang, G.; Shen, Q.; Zhang, R. Analysis and cloning of the synthetic pathway of the phytohormone indole-3-acetic acid in the plant-beneficial Bacillus amyloliquefaciens SQR9. Microb. Cell Fact. 2015, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Fadiji, A.E.; Babalola, O.O. Elucidating Mechanisms of Endophytes Used in Plant Protection and Other Bioactivities With Multifunctional Prospects. Front. Bioeng. Biotechnol. 2020, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Slama, H.; Cherif-Silini, H.; Chenari Bouket, A.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.; Luptakova, L.; Triki, M.; Vallat, A.; et al. Screening for Fusarium antagonistic bacteria from contrasting niches designated the endophyte Bacillus halotolerans as plant warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef]
- Andrés-Barrao, C.; Lafi, F.F.; Alam, I.; De Zélicourt, A.; Eida, A.A.; Bokhari, A.; Alzubaidy, H.; Bajic, V.B.; Hirt, H.; Saad, M.M. Complete genome sequence analysis of Enterobacter sp. SA187, a plant multi-stress tolerance promoting endophytic bacterium. Front. Microbiol. 2017, 8, 2023. [Google Scholar] [CrossRef]
- Mehnaz, D.; Mukhtar, S.; Ishaq, A.; Hassan, S.; Abdulla, K.; Mirza, M.S. Comparison of microbial communities associated with halophyte (Salsola stocksii) and non-halophyte (Triticum aestivum) using culture-independent approaches. Pol. J. Microbiol. 2017, 66, 353–364. [Google Scholar] [CrossRef]
- Abbas, H.; Patel, R.M.; Parekh, V.B. Culturable endophytic bacteria from halotolerant Salicornia brachata L.: Isolation and plant growth promoting traits. Indian J. Appl. Microbiol. 2018, 21, 10–21. [Google Scholar] [CrossRef]
- Chebotar, V.K.; Chizhevskaya, E.P.; Baganova, M.E.; Keleinikova, O.V.; Yuzikhin, O.S.; Zaplatkin, A.N.; Khonina, O.V.; Kostitsin, R.D.; Lapenko, N.G. Endophytes from halotolerant plants aimed to overcome salinity and draught. Plants 2022, 11, 2992. [Google Scholar] [CrossRef] [PubMed]
- Mirskaya, G.V.; Khomyakov, Y.V.; Rushina, N.A.; Vertebny, V.E.; Chizhevskaya, E.P.; Chebotar, V.K.; Chesnokov, Y.V.; Pishchik, V.N. Plant development of early-maturing spring wheat (Triticum aestivum L.) under inoculation with Bacillus sp. V2026. Plants 2022, 11, 1817. [Google Scholar] [CrossRef] [PubMed]
- Chebotar, V.K.; Chizhevskaya, E.P.; Vorobyov, N.I.; Bobkova, V.V.; Pomyaksheva, L.V.; Khomyakov, Y.V.; Konovalov, S.N. The quality and productivity of strawberry (Fragaria × ananassa Duch.) improved by the inoculation of PGPR Bacillus velezensis BS89 in field experiments. Agronomy 2022, 12, 2600. [Google Scholar] [CrossRef]
- Chebotar, V.K.; Chizhevskaya, E.P.; Andronov, E.E.; Vorobyov, N.I.; Keleinikova, O.V.; Baganova, M.E.; Konovalov, S.N.; Filippova, P.S.; Pishchik, V.N. Assessment of the rhizosphere bacterial community under maize growth using various agricultural technologies with biomodified mineral fertilizers. Agronomy 2023, 13, 1855. [Google Scholar] [CrossRef]
- Timofeeva, T.A.; Chebotar, V.K.; Demidov, D.V.; Gaidukova, S.E.; Yakovleva, I.V.; Kamionskaya, A.M. Effects of apatite concentrate in combination with phosphate-solubilizing microorganisms on the yield of ryegrass cultivar Izorskiy. Agronomy 2023, 13, 1568. [Google Scholar] [CrossRef]
- Chebotar, V.K.; Zaplatkin, A.N.; Balakina, S.V.; Gadzhiev, N.M.; Lebedeva, V.A.; Khiutti, A.V.; Chizhevskaya, E.P.; Filippova, P.S.; Keleinikova, O.V.; Baganova, M.E.; et al. The effect of endophytic bacteria Bacillus thuringiensis W65 and B. amyloliquefaciens P20 on the yield and the incidence of potato rhizoctoniosis and late blight. Sel’skokhozyaistvennaya Biol. 2023, 58, 429–446. [Google Scholar] [CrossRef]
- Bokhari, A.; Essack, M.; Lafi, F.F.; Andres-Barrao, C.; Jalal, R.; Alamoudi, S.; Razali, R.; Alzubaidy, H.; Shah, K.H.; Siddique, S.; et al. Bioprospecting desert plant Bacillus endophytic strains for their potential to enhance plant stress tolerance. Sci. Rep. 2019, 9, 18154. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- National Library of Medicine; Natilnal Centre for Biotechnology Information. Tax-Onomy Browser. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi (accessed on 16 December 2023).
- Ishag, A.E.S.A.; Abdelbagi, A.O.; Hammad, A.M.A.; Elsheikh, E.A.E.; Elsaid, O.E.; Hur, J.H.; Laing, M.D. Biodegradation of chlorpyrifos, malathion, and dimethoate by three strains of bacteria isolated from pesticide-polluted soils in sudan. J. Agricult. Food Chem. 2016, 64, 8491–8498. [Google Scholar] [CrossRef]
- Wahla, A.Q.; Iqbal, S.; Anwar, S.; Firdous, S.; Mueller, J.A. Optimizing the metribuzin degrading potential of a novel bacterial consortium based on Taguchi design of experiment. J. Hazard. Mater. 2019, 366, 1–9. [Google Scholar] [CrossRef]
- Mayer, F.L.; Kronstad, J.W. Disarming fungal pathogens: Bacillus safensis inhibits virulence factor production and biofilm formation by Cryptococcus neoformans and Candida albicans. MBio 2017, 8, e01537-17. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Chakraborty, B.; Chakraborty, U. Plant Growth promoting rhizobacteria protect wheat plants against temperature stress through antioxidant signalling and reducing chloroplast and membrane injury. J. Plant Growth Regul. 2018, 37, 1396–1412. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Liu, J.; Guo, W.-H.; Li, X.-B.; Xia, J.-B.; Xie, W.-J.; Yao, Z.-G.; Zhang, Y.-M.; Wang, R.-Q. Characterization and initial application of endophytic Bacillus safensis strain ZY16 for improving phytoremediation of oil-contaminated saline soils. Front. Microbiol. 2019, 10, 991. [Google Scholar] [CrossRef] [PubMed]
- Lanna-Filho, R.; Pozzebon, B.C.; de Oliveira, A.M.R. Endophytic bacteria Bacillus safensis and Pseudomonas hibiscicola and their ability to increase rice seedling growth. Arq. Inst. Biol. 2022, 89, e00162021. [Google Scholar] [CrossRef]
- Kazan, K. Auxin and the integration of environmental signals into plant root development. Ann. Bot. 2013, 112, 1655–1665. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76–85. [Google Scholar] [CrossRef]
- Jung, J.; Park, C. Auxin modulation of salt stress signaling in Arabidopsis seed germination. Plant Signal. Behav. 2011, 6, 1198–1200. [Google Scholar] [CrossRef] [PubMed]
- Schmülling, T. New insights into the functions of cytokinins in plant development. J. Plant Growth Regul. 2002, 21, 40–49. [Google Scholar] [CrossRef]
- Borrell, A.K.; Hammer, G.L.; Henzell, R.G. Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci. 2000, 40, 1037–1048. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, W.Q.; Zhang, G.L.; Kaminek, M.; Dobrev, P.; Xu, J.; Gruissem, W. Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J. Integr. Plant Biol. 2010, 52, 653–669. [Google Scholar] [CrossRef]
- Wilkinson, S.; Kudoyarova, G.R.; Veselov, D.S.; Arkhipova, T.N.; Davies, W.J. Plant hormone interactions: Innovative targets for crop breeding and management. J. Exp. Bot. 2012, 63, 3499–3509. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, T.N.; Veselov, S.U.; Melentiev, A.I.; Kudoyarova, G.R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil. 2005, 272, 201–215. [Google Scholar] [CrossRef]
- Pal, K.K.; Dey, R.; Sherathia, D.N.; Devidayal; Mangalassery, S.; Kumar, A.; Rupapara, R.B.; Mandaliya, M.; Rawal, P.; Bhadania, R.A.; et al. Alleviation of salinity stress in peanut by application of endophytic bacteria. Front. Microbiol. 2021, 12, 650771. [Google Scholar] [CrossRef] [PubMed]
- Sorty, A.M.; Meena, K.K.; Choudhary, K.; Bitla, U.M.; Minhas, P.S.; Krishnani, K.K. Effect of plant growth promoting bacteria associated with halophytic weed (Psoralea corylifolia L.) on germination and seedling growth of wheat under saline conditions. Appl. Biochem. Biotechnol. 2016, 180, 872–882. [Google Scholar] [CrossRef]
- Cherif-Silini, H.; Thissera, B.; Bouket, A.C.; Saadaoui, N.; Silini, A.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Yahiaoui, B. Durum wheat stress tolerance induced by endophyte Pantoea agglomerans with genes contributing to plant functions and sec-ondary metabolite arsenal. Int. J. Mol. Sci. 2019, 20, 3989. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Jha, P.N. A halotolerant bacterium Bacillus licheniformis HSW-16 augments induced systemic tolerance to salt stress in wheat plant (Triticum aestivum). Front. Plant Sci. 2016, 7, 1890. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Kucharova, Z.; Davranov, K.; Berg, G.; Makarova, N.; Azarova, T.; Chebotar, V.; Tikhonovich, I.; Kamilova, F.; Validov, S.Z.; et al. Bacteria able to control foot and root rot and to promote growth of cucumber in salinated soils. Biol. Fertil. Soils 2011, 47, 197–205. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Alimov, J.; Shurigin, V.; Alaylar, B.; Wirth, S.; Bellingrath-Kimura, S.D. Diversity and plant growth-promoting ability of endophytic, halotolerant bacteria associated with Tetragonia tetragonioides (Pall.) Kuntze. Plants 2022, 11, 49. [Google Scholar] [CrossRef]
- Mohamad, O.A.A.; Ma, J.-B.; Liu, Y.-H.; Zhang, D.; Hua, S.; Bhute, S.; Hedlund, B.P.; Li, W.-J.; Li, L. Beneficial endophytic bacterial populations associated with medicinal plant Thymus vulgaris alleviate salt stress and confer resistance to Fusarium oxysporum. Front. Plant Sci. 2020, 11, 47. [Google Scholar] [CrossRef]
- Naz, I.; Bano, A.; Ul-Hassan, T. Isolation of phytohormones producing plant growth promoting rhizobacteria from weeds growing in Khewra salt range, Pakistan and their implication in providing salt tolerance to Glycine max L. Afr. J. Biotechnol. 2009, 8, 5762–5766. [Google Scholar] [CrossRef]
- Czarnes, S.; Hallett, P.D.; Bengough, A.G.; Young, I.M. Root- and microbial-derived mucilages affect soil structure and water transport. Eur. J. Soil Sci. 2000, 51, 435–443. [Google Scholar] [CrossRef]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Yancey, P.H. Compatible and counteracting solutes. In Cellular and Molecular Physiology of Cell Volume Regulation; Strange, K., Ed.; CRC Press: Boca Raton, FL, USA, 1994; pp. 81–109. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Rhodes, D.; Hanson, A.D. Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2003, 44, 357–384. [Google Scholar] [CrossRef]
- Bremer, E.; Kramer, R. Coping with osmotic challenges: Osmoregulation through accumulation and release of compatible solutes. In Bacterial Stress Responses; Storz, G., Hengge-Aronis, R., Eds.; American Society for Microbiology: Washington, DC, USA, 2000; pp. 79–97. [Google Scholar]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic stress. Microbiol Rev. 1989, 53, 121–147. [Google Scholar] [CrossRef] [PubMed]
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Berg, G. Impact of formulation procedures on the effect of the biocontrol agent Serratia plymuthica HRO-C48 on Verticillium wilt in oilseed rape. BioControl 2008, 53, 905–913. [Google Scholar] [CrossRef]
- Cardinale, M.; de Castro, J.V.; Müller, H.; Berg, G.; Grube, M. In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria. FEMS Microbiol. Ecol. 2008, 66, 63–71. [Google Scholar] [CrossRef]
- Hoffland, E.; Findenberg, G.; Nielemans, J. Solubilization of rock phosphate by rape. Plant Soil 1989, 113, 161–165. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 June 2023).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteases (Halo Zone) (mm) | Lipases (Halo Zone) (mm) | Beta-Glucanases (Halo Zone) (mm) | Cellulases (Halo Zone) (mm) |
|---|---|---|---|
| 12.7 ± 1.1 | 7.1 ± 0.9 | 2.5 ± 0.03 | 17.3 ± 1.1 |
| IAA (µg/L) | ABA (µg/L) | tZ (µg/L) |
|---|---|---|
| 366.1 ± 12.5 | 0.646 ± 0.02 | 0.822 ± 0.04 |
| 1% NaCl | 5% NaCl | 10% NaCl | 15% NaCl | 20% NaCl |
|---|---|---|---|---|
| 109.2 ± 8.7 | 33.5 ± 2.7 | 12.6 ± 1.3 | 0.7 ± 0.03 | 0 |
| NaCl Concentration | Average Length of Seedling (mm) | Average Length of Root (mm) |
|---|---|---|
| Radish | ||
| 0 | 30.1 ± 2.2 a | 61.0 ± 4.1 a |
| 0.5 | 23.3 ± 1.3 b | 51.5 ± 3.5 b |
| 1.0 | 18.7 ± 1.0 c | 38.7 ± 2.5 c |
| 1.5 | 12.5 ± 0.9 d | 11.3 ± 1.5 d |
| 2.0 | 4.5 ± 0.5 e | 6.1 ± 0.5 e |
| 2.5 | - | - |
| 5.0 | - | - |
| Oats | ||
| 0 | 83.5 ± 5.5 a | 106.1 ± 8.3 a |
| 0.5 | 52.5 ± 3.7 b | 87.3 ± 6.1 b |
| 1.0 | 29.1 ± 2.3 c | 63.5 ± 4.7 c |
| 1.5 | 16.7 ± 0.9 d | 35.1 ± 1.7 d |
| 2.0 | 9.3 ± 0.5 e | 23.7 ± 1.1 e |
| 2.5 | 3.3 ± 0.3 f | 5.5 ± 0.3 f |
| 5.0 | - | - |
| Winter Wheat | ||
| 0 | 43.3 ± 2.9 a | 89.1 ± 6.7 a |
| 0.5 | 26.7 ± 1.5 b | 48.3 ± 3.1 b |
| 1.0 | 19.5 ± 1.1 c | 37.1 ± 2.0 c |
| 1.5 | 14.7 ± 0.7 d | 23.1 ± 1.3 d |
| 2.0 | 5.6 ± 0.3 e | 10.1 ± 0.5 e |
| 2.5 | - | |
| 5.0 | - | |
| Variant | Shoot Length (mm) | Root Length (mm) |
|---|---|---|
| Negative control without NaCl | 166 ± 10.1 a | 79 ± 5.3 a |
| Positive control with NaCl | 26 ± 1.9 d | 29 ± 1.5 b |
| NaCl + TS3, 105 CFU/mL | 85 ± 7.6 c | 51 ± 4.7 c |
| NaCl + TS3, 106 CFU/mL | 90 ± 7.9 c | 52 ± 4.5 c |
| NaCl + TS3, 107 CFU/mL | 100 ± 8.3 b | 52 ± 4.3 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebotar, V.K.; Zaplatkin, A.N.; Chizhevskaya, E.P.; Gancheva, M.S.; Voshol, G.P.; Malfanova, N.V.; Baganova, M.E.; Khomyakov, Y.V.; Pishchik, V.N. Phytohormone Production by the Endophyte Bacillus safensis TS3 Increases Plant Yield and Alleviates Salt Stress. Plants 2024, 13, 75. https://doi.org/10.3390/plants13010075
Chebotar VK, Zaplatkin AN, Chizhevskaya EP, Gancheva MS, Voshol GP, Malfanova NV, Baganova ME, Khomyakov YV, Pishchik VN. Phytohormone Production by the Endophyte Bacillus safensis TS3 Increases Plant Yield and Alleviates Salt Stress. Plants. 2024; 13(1):75. https://doi.org/10.3390/plants13010075
Chicago/Turabian StyleChebotar, Vladimir K., Alexander N. Zaplatkin, Elena P. Chizhevskaya, Maria S. Gancheva, Gerben P. Voshol, Natalia V. Malfanova, Maria E. Baganova, Yuriy V. Khomyakov, and Veronika N. Pishchik. 2024. "Phytohormone Production by the Endophyte Bacillus safensis TS3 Increases Plant Yield and Alleviates Salt Stress" Plants 13, no. 1: 75. https://doi.org/10.3390/plants13010075