
Two Intra-Individual ITS1 rDNA Sequence Variants Identified in the Female and Male Rotylenchulus reniformis Populations of Alabama

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Nematode DNA Extractions from Soil
4.2. Quality Check, PCR Amplification, and Determination of the Product
4.3. Cloning and Colony PCR
4.4. Sequencing
4.5. Alignment and Phylogenetic Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goraya, M.; Yan, G.; Plaisance, A.; Handoo, Z.A. Identification and reproduction of dagger nematode, Xiphinema americanum, in potato. Nematology 2023, 25, 1127–1139. [Google Scholar] [CrossRef]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture–Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Linford, M.B.; Oliveira, J.M. Rotylenechulus reniformis, nov. gen., n. sp., a nematode parasite of roots. Proc. Helminthol. Soc. Wash. 1940, 7, 35–42. Available online: https://www.cabdirect.org/abstracts/19400800641.html (accessed on 29 May 2022).
- Jones, L.M.; Koehler, A.; Trnka, M.; Bálek, J.; Challinor, A.J.; Atkinson, H.J.; Urwin, P.E. Climate change is predicted to alter the current pest status of Globodera pallida and G. rostochiensis in the United Kingdom. Glob. Change Biol. 2017, 23, 4497–4507. [Google Scholar] [CrossRef] [PubMed]
- Eapen, S.J.; Pandey, R. Nematode parasites of spices and medicinal plants. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; CAB International eBooks; CAB International: Wallingford, UK, 2018; pp. 755–794. [Google Scholar] [CrossRef]
- Robinson, A.F.; Inserra, R.N.; Caswell-Chen, E.P.; Vovlas, N.; Troccoli, A. Review: Rotylenchulus species: Identification, distribution, host ranges, and crop plant resistance. Nematropica 1997, 27, 127–180. Available online: http://journals.fcla.edu/nematropica/article/view/64190 (accessed on 29 January 2022).
- Pulavarty, A.; Egan, A.; Karpinska, A.; Horgan, K.; Kakouli-Duarte, T. Plant parasitic nematodes: A review on their behaviour, host interaction, management approaches and their occurrence in two sites in the republic of Ireland. Plants 2021, 10, 2352. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Cantalapiedra-Navarrete, C.; Archidona-Yuste, A.; Tzortzakakis, E.A.; Birmpilis, I.G.; Vovlas, N.; Subbotin, S.A.; Castillo, P. Prevalence and molecular diversity of reniform nematodes of the genus Rotylenchulus (Nematoda: Rotylenchulinae) in the mediterranean basin. Eur. J. Plant Pathol. 2018, 150, 439–455. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Clavero-Camacho, I.; Archidona-Yuste, A.; Cantalapiedra-Navarrete, C.; León-Ropero, G.; Miyara, S.B.; Karssen, G.; Castillo, P. Global distribution of the reniform nematode genus Rotylenchulus with the synonymy of Rotylenchulus macrosoma with Rotylenchulus borealis. Plants 2020, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Archidona-Yuste, A.; Cantalapiedra-Navarrete, C.; Azpilicueta, A.S.; Saborido, A.; Tzortzakakis, E.A.; Cai, R.; Castillo, P. New distribution and molecular diversity of the reniform nematode Rotylenchulus macrosoma (Nematoda: Rotylenchulinae) in Europe. Phytopathology 2021, 111, 720–730. [Google Scholar] [CrossRef]
- Williams, J.R. Studies on the Nematode Soil Fauna of Sugar Cane Fields in Mauritius: 4: Tylenchoidea (Partim); Mauritius Sugar Industry Research Institute: Moka, Mauritius, 1960; p. 4. [Google Scholar]
- Loof, P.; Oostenbrink, M. Rotylenchulus borealis n.sp. with a key to the species of Rotylenchulus. Nematologica 1962, 7, 83–90. [Google Scholar] [CrossRef]
- Dasgupta, D.R.; Raski, D.J.; Sher, S.A. A revision of the genus Rotylenchulus Linford and Oliveira, 1940 (Nematoda: Tylenchiclae). Proc. Helminthol. Soc. Wash. 1968, 35, 169–192. [Google Scholar]
- IPCC. Intergovernmental Panel on Climate Change. Special Report on the Ocean and Cryosphere in a Changing Climate (SROCC). 2019. Available online: https://archive.ipcc.ch/index.html (accessed on 17 September 2020).
- Colagiero, M.; Ciancio, A. Climate changes and nematodes: Expected effects and perspectives for plant protection. Redia G. Zool. 2012, 94, 113–118. Available online: https://journals-crea.4science.it/index.php/redia/article/download/656/592 (accessed on 24 January 2022).
- Khanal, C.; McGawley, E.C.; Overstreet, C.M.; Stetina, S.R. The elusive search for reniform nematode resistance in cotton. Phytopathology 2018, 108, 532–541. [Google Scholar] [CrossRef]
- Wilkes, J.E.F. Mapping Genetic Resistance to Reniform Nematode (Rotylenchulus reniformis) in Soybean. Ph.D. Thesis, Clemson University, Clemson, SC, USA, 2020. [Google Scholar]
- Calegari, A.; Tiecher, T.; Wutke, E.B.; Canalli, L.B.D.S.; Bunch, R.; Rheinheimer, D.D.S.; Consultant, B. The role and management of soil mulch and cover crops in conservation agriculture systems. In Advances in Conservation Agriculture; Burleigh Dodds Series in Agricultural Science; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 179–248. [Google Scholar] [CrossRef]
- Kularathna, M.; Overstreet, C.M.; McGawley, E.C.; Stetina, S.R.; Khanal, C.; Godoy, F.M.C.; McInnes, B. Pathogenicity and reproduction of isolates of reniform nematode, Rotylenchulus reniforms, from Louisiana on soybean. Nematropica 2019, 49, 31–41. Available online: https://journals.flvc.org/nematropica/article/view/115613 (accessed on 17 August 2022).
- Robinson, A.F. Reniform in U.S. Cotton: When, where, why, and some remedies. Annu. Rev. Phytopathol. 2007, 45, 263–288. [Google Scholar] [CrossRef]
- Lim, J.; Kim, W.; Kim, J.; Lee, J. Telomeric repeat evolution in the phylum nematoda revealed by high-quality genome assemblies and subtelomere structures. Genome Res. 2023, 33, 1947–1957. [Google Scholar] [CrossRef]
- Meng, L.; Gao, Y.; Gong, L. Recombination and incomplete concerted evolution of the ribosomal 18S (partial) -ITS1-5.8S-ITS2-28S (partial) rDNA in Cynoglossus trigrammus genome. Biochem. Syst. Ecol. 2022, 105, 104513. [Google Scholar] [CrossRef]
- Delva, E.; Arisuryanti, T.; Ilmi, M. Genetic diversity of Amylomyces rouxii from ragi tapai in Java Island based on ribosomal regions ITS1/ITS2 and D1/D2. Mycobiology 2022, 50, 132–141. [Google Scholar] [CrossRef]
- Terrana, L.; Flot, J.; Eeckhaut, I. ITS1 variation among Stichopathes cf. maldivensis (Hexacorallia: Antipatharia) whip black corals unveils conspecificity and population connectivity at local and global scales across the Indo-Pacific. Coral Reefs 2021, 40, 521–533. [Google Scholar] [CrossRef]
- Mlinarec, J.; Boštjančić, L.L.; Malenica, N.; Jurkovic, A.; Boland, T.; Yakovlev, S.S.; Besendorfer, V. Structure and methylation of 35S rDNA in Allopolyploids Anemone multifida (2n = 4x = 32, BBDD) and Anemone baldensis (2n = 6x = 48, AABBDD) and their parental species show evidence of nucleolar dominance. Front. Plant Sci. 2022, 13, 908218. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Zhu, L.; Li, Z.; Jiang, R.; Liu, S.; Huang, W.; Li, C.; Kong, L.; Peng, D.; Peng, H. Population genetics of the cereal cyst nematode Heterodera avenae reveal geographical segregation and host adaptation. Phytopathol. Res. 2023, 5, 30. [Google Scholar] [CrossRef]
- Nyaku, S.T.; Kantety, R.V.; Tilahun, Y.; Lawrence, K.; Soliman, K.M.; Cebert, E.; Sharma, G.C. 18S and ITS1 genomic sequence variations in Rotylenchulus reniformis isolates from Alabama. J. Cotton Sci. 2013, 17, 184–194. Available online: https://www.cabdirect.org/cabdirect/abstract/20133321761?q=(au%3a%22Kantety%2c+R.+V.%22) (accessed on 24 January 2022).
- Djiwanti, S.R.; Miftakhurohmah. Molecular detection and identification of the foliar nematode Aphelenchoides fragariae on Andrographis paniculata in Indonesia. Australas. Plant Pathol. 2022, 51, 301–304. [Google Scholar] [CrossRef]
- Ponomareva, E.; Badiss, A.; Sultana, T.; Yu, Q.; Nguyen, H.D. Genome characterization and development of real-time PCR assays for Ditylenchus dipsaci and D. weischeri. J. Nematol. 2022, 54, 20220058. [Google Scholar] [CrossRef] [PubMed]
- Kantor, M.R.; Subbotin, S.A.; Huse, G.; Handoo, Z.A. Morphological and molecular characterization of Punctodera stonei brzeski, 1998 (Nematoda: Heteroderidae) from Virginia, USA. J. Nematol. 2022, 54, 20220008. [Google Scholar] [CrossRef]
- Gong, L.; Luo, H.; Shi, W.; Yang, M. Intra-individual variation and transcribed pseudogenes in the ribosomal ITS1-5.8S-ITS2 rDNA of Paraplagusia japonica (Pleuronectiformes: Cynoglossidae). Biochem. Biophys. Res. Commun. 2019, 513, 726–731. [Google Scholar] [CrossRef]
- Van Den Berg, E.; Palomares-Rius, J.E.; Vovlas, N.; Tiedt, L.R.; Castillo, P.; Subbotin, S.A. Morphological and molecular characterisation of one new and several known species of the reniform nematode, Rotylenchulus Linford & Oliveira, 1940 (Hoplolaimidae: Rotylenchulinae), and a phylogeny of the genus. Nematology 2016, 18, 67–107. [Google Scholar] [CrossRef]
- Agudelo, P.; Robbins, R.T.; Stewart, J.M.; Szalanski, A.L. Intraspecific variability of Rotylenchulus reniformis from cotton-growing regions in the United States. J. Nematol. 2005, 37, 105. Available online: https://pubmed.ncbi.nlm.nih.gov/19262849 (accessed on 24 January 2022).
- Nakae, K.; Yoshiga, T. The variations of ribosomal ITS genes in Rotylenchulus reniformis in Japan, and phylogenetic analysis of its reproductive types based on the mitochondrial COI gene. Nihon Senchu Gakkaishi 2021, 51, 21–28. [Google Scholar] [CrossRef]
- Chow, S.; Hamasaki, K.; Konishi, K.; Yanagimoto, T.; Wagatsuma, R.; Takeyama, H. Variation of length and sequence of the nuclear ribosomal DNA internal transcribed spacer 1 supports “hermit-to-king” crab hypothesis. Crustac. Res. 2022, 52, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.E.; Cooper, R.D.; Beebe, N.W. Internal repetition and intraindividual variation in the rDNA ITS1 of the anopheles punctulatus group (Diptera: Culicidae): Multiple units and rates of turnover. J. Mol. Evol. 2009, 68, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Jenifer, A.; Sharma, S.; Singh, R.; Chakrabarti, S. Morphological and molecular characterization of potato cyst nematode populations from the Nilgiris. Indian J. Agric. Sci. 2020, 90, 273–278. [Google Scholar]
- Bhat, K.A.; Mir, R.A.; Farooq, A.; Manzoor, M.; Hami, A.; Allie, K.A.; Wani, S.M.; Khan, M.N.; Sayyed, R.Z.; Poczai, P.; et al. Advances in nematode identification: A journey from fundamentals to evolutionary aspects. Diversity 2022, 14, 536. [Google Scholar] [CrossRef]
- Ni, C.; Han, B.; Liu, Y.; Maria, M.; Liu, S.; Li, W.; Shi, M.; Li, H.; Peng, D. Diagnosis and characterization of the ribosomal DNA-ITS of potato rot nematode (Ditylenchus destructor) populations from Chinese medicinal herbs. J. Integr. Agric. 2023, 22, 1763–1781. [Google Scholar] [CrossRef]
- Takabayashi, M.; Carter, D.A.; Ward, S.; Hoegh-Guldberg, O. Inter-and intra-specific variability in ribosomal DNA sequence in the internal transcribed spacer region of corals. In Proceedings of the 75th Conference of the Australian Coral Reef Society, Brisbane, Australia, 2–6 October 1997; University of Queensland: Brisbane, Australia, 1998; pp. 241–248. [Google Scholar]
- Spakulova, M.; Bombarova, M.; Miklisova, D.; Nechybova, S.; Langrova, I. How to become a successful invasive tapeworm: A case study of abandoned sexuality and exceptional chromosome diversification in the triploid carp parasite Atractolytocestus huronensis Anthony, 1958 (Caryophyllidea: Lytocestidae). Parasites Vectors 2019, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Tatonova, Y.V.; Chelomina, G.N.; Besprosvannykh, V.V. Genetic diversity of nuclear ITS1–5.8S–ITS2 rDNA sequence in clonorchis sinensis cobbold, 1875 (Trematoda: Opisthorchidae) from the Russian far East. Parasitol. Int. 2012, 61, 664–674. [Google Scholar] [CrossRef]
- Qureshi, A.W.; Javed, B.; Khan, L. Analysis of internal transcribed spacer1 (ITS1) region of rDNA for genetic characterization of Paramphistomum sp. Saudi J. Biol. Sci. 2021, 28, 5617–5620. [Google Scholar] [CrossRef]
- Xue, Q.; Bik, H.M.; Yergaliyev, T.; Gu, J.; Fonderie, P.; Brown-Miyara, S.; Szitenberg, A.; Bert, W. Widespread prevalence but contrasting patterns of intragenomic rRNA polymorphisms in nematodes: Implications for phylogeny, species delimitation and life history inference. Mol. Ecol. Resour. 2019, 20, 318–332. [Google Scholar] [CrossRef]
- Tilahun, Y.; Soliman, K.M.; Lawrence, K.; Cseke, L.J.; Joel, W. Nuclear ribosomal DNA diversity of a cotton pest (Rotylenchulus reniformis) in the United States. Afr. J. Biotechnol. 2008, 7, 3217–3224. [Google Scholar] [CrossRef]
- Deng, Y.; Xu, H.; Tian, Z.; Zheng, J. Interindividual variability of internal transcribed spacer region of ribosomal DNA in Rotylenchulus reniformis. J. Zhejiang Univ. Agric. Life Sci. 2015, 41, 252–260. Available online: https://www.cabdirect.org/cabdirect/abstract/20153302316 (accessed on 24 January 2022).
- Nyaku, S.T.; Kantety, R.V.; Cebert, E.; Lawrence, K.; Honger, J.O.; Sharma, G.C. Principal component analysis and molecular characterization of reniform nematode populations in Alabama. Plant Pathol. J. 2016, 32, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Overstreet, R.M.; Subbotin, S.A. ITS2 secondary structure and phylogeny of cyst-forming nematodes of the genus Heterodera (Tylenchida: Heteroderidae). Org. Divers. Evol. 2008, 8, 182–193. [Google Scholar] [CrossRef]
- Subbotin, S.A.; Sturhan, D.; Vovlas, N.; Castillo, P.; Tambe, J.T.; Moens, M.; Baldwin, J.G. Application of the secondary structure model of rRNA for phylogeny: D2–D3 expansion segments of the LSU gene of plant-parasitic nematodes from the family Hoplolaimidae Filipjev, 1934. Mol. Phylogenet. Evol. 2007, 43, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A.; Deimi, A.M.; Zheng, J.; Chizhov, V.N. Length variation and repetitive sequences of Internal Transcribed Spacer of ribosomal RNA gene, diagnostics, and relationships of populations of potato rot nematode, Ditylenchus destructor Thorne, 1945 (Tylenchida: Anguinidae). Nematology 2011, 13, 773–785. [Google Scholar] [CrossRef]
- Marek, M.; Zouhar, M.; Douda, O.; Mazáková, J.; Ryšánek, P. Bioinformatics-assisted characterization of the ITS1-5·8S-ITS2 segments of nuclear rRNA gene clusters, and its exploitation in molecular diagnostics of European crop-parasitic nematodes of the genus Ditylenchus. Plant Pathol. 2010, 59, 931–943. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M.; Stiegler, P. Optimal computer folding of large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Res. 1981, 9, 133–148. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.H.; Siederdissen, C.H.Z.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- McCaskill, J.S. The equilibrium partition function and base pair binding probabilities for RNA secondary structure. Biopolymers 1990, 29, 1105–1119. [Google Scholar] [CrossRef]
- Ding, Y.; Chan, C.Y.; Lawrence, C.E. RNA secondary structure prediction by centroids in a boltzmann weighted ensemble. RNA 2005, 11, 1157–1166. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
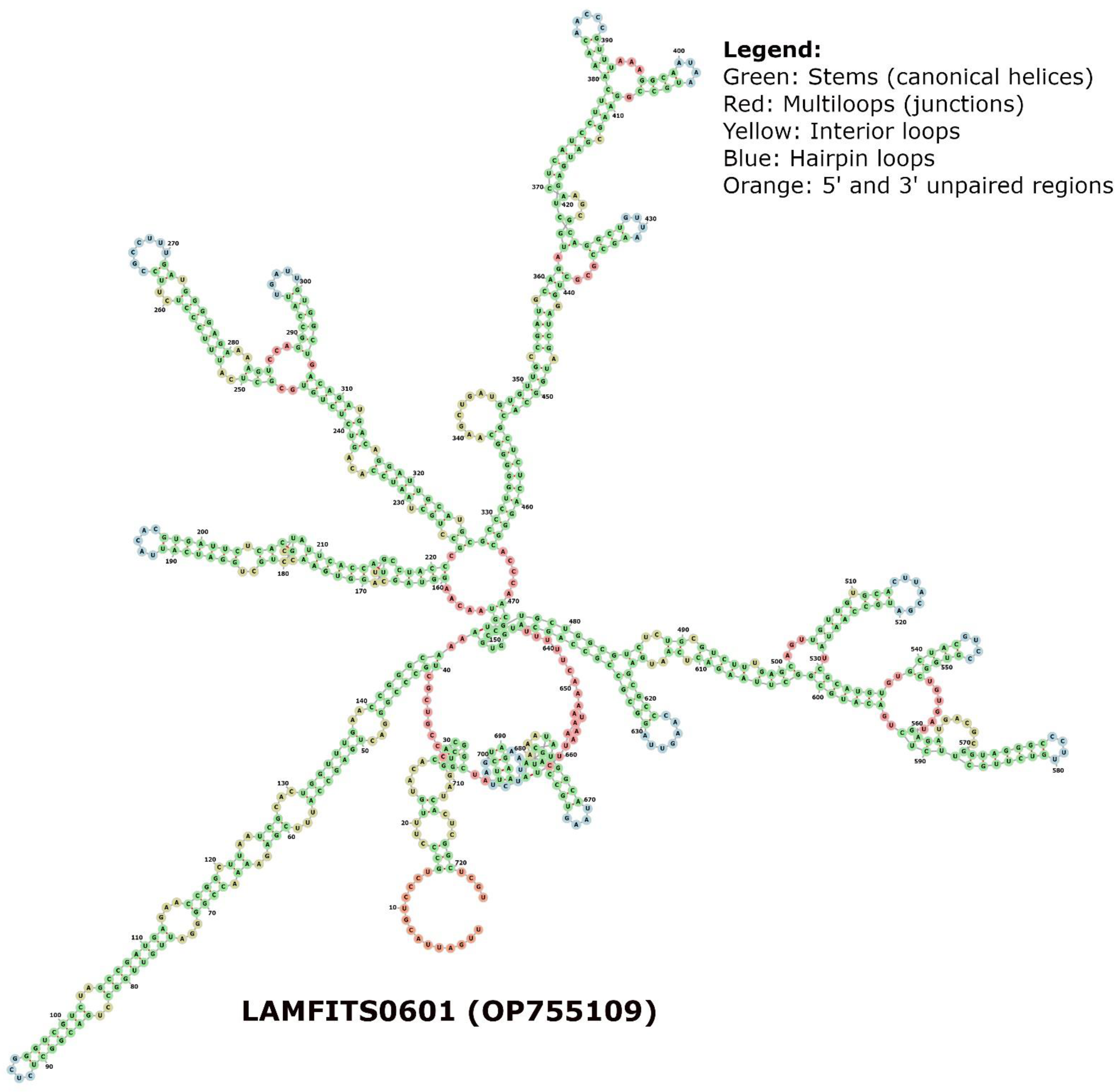{kind=link}
| Hamilton Population | BelleMina Population | Lamons Population | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |||||||
| ITS1-557 | ITS1-732 | ITS1-567 | ITS1-736 | ITS1-588 | ITS1-735 | ITS1-570 | ITS1-729 | ITS1-559 | ITS1-726 | ITS1-559 | ITS1-726 | |
| T% | 27.20 | 25.70 | 27.20 | 25.70 | 27.10 | 25.70 | 27.10 | 25.70 | 27.40 | 25.70 | 27.10 | NA |
| C% | 24.60 | 25.90 | 24.60 | 25.60 | 24.50 | 26.10 | 24.60 | 25.70 | 24.40 | 25.80 | 24.80 | NA |
| A% | 24.30 | 22.10 | 24.40 | 22.30 | 24.40 | 22.30 | 24.50 | 22.10 | 24.30 | 22.20 | 24.40 | NA |
| G% | 23.80 | 26.40 | 23.80 | 26.30 | 24.10 | 25.90 | 23.80 | 26.50 | 23.90 | 26.40 | 23.80 | NA |
| Hamilton Population | BelleMina Population | Lamons Population | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |||||||
| ITS1-557 | ITS1-732 | ITS1-567 | ITS1-736 | ITS1-588 | ITS1-735 | ITS1-570 | ITS1-729 | ITS1-559 | ITS1-726 | ITS1-559 | ITS1-726 | |
| Singletons | 44/557 | 57/732 | 57/567 | 72/736 | 116/588 | 74/735 | 93/570 | 52/729 | 14/559 | 63/726 | 22/559 | NA |
| Conserved sites | 462/557 | 631/732 | 418/567 | 563/736 | 275/588 | 580/735 | 385/570 | 626/729 | 534/559 | 621/726 | 503/559 | NA |
| Parsimony-informative sites | 51/557 | 41/732 | 90/567 | 98/736 | 172/588 | 74/735 | 85/570 | 49/729 | 2/559 | 40/726 | 29/559 | NA |
| Variable sites | 95/557 | 98/732 | 147/567 | 170/736 | 290/588 | 148/735 | 178/570 | 101/729 | 16/559 | 103/726 | 51/559 | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyaku, S.T.; Karapareddy, S.; Cebert, E.; Lawrence, K.; Eleblu, J.S.Y.; Sharma, G.C.; Sripathi, V.R. Two Intra-Individual ITS1 rDNA Sequence Variants Identified in the Female and Male Rotylenchulus reniformis Populations of Alabama. Plants 2024, 13, 5. https://doi.org/10.3390/plants13010005
Nyaku ST, Karapareddy S, Cebert E, Lawrence K, Eleblu JSY, Sharma GC, Sripathi VR. Two Intra-Individual ITS1 rDNA Sequence Variants Identified in the Female and Male Rotylenchulus reniformis Populations of Alabama. Plants. 2024; 13(1):5. https://doi.org/10.3390/plants13010005
Chicago/Turabian StyleNyaku, Seloame T., Sowndarya Karapareddy, Ernst Cebert, Kathy Lawrence, John S. Y. Eleblu, Govind C. Sharma, and Venkateswara R. Sripathi. 2024. "Two Intra-Individual ITS1 rDNA Sequence Variants Identified in the Female and Male Rotylenchulus reniformis Populations of Alabama" Plants 13, no. 1: 5. https://doi.org/10.3390/plants13010005