

Plasticity in the Morphology of Growing Bamboo: A Bayesian Analysis of Exogenous Treatment Effects on Plant Height, Internode Length, and Internode Numbers

Abstract
:1. Introduction
- (1)
- Does exogenous Suc or GA treatment promote the plant height of P. edulis in the wild?
- (2)
- Is it a large elongation of a few internodes or a small elongation of all the internodes that promotes an increase in the plant height of P. edulis?
- (3)
- Does having more internodes promote the increase in plant height of P. edulis?
- (4)
- Does the promotion of internode length under exogenous Suc and GA treatments diminish with distance from the injection point?
2. Results
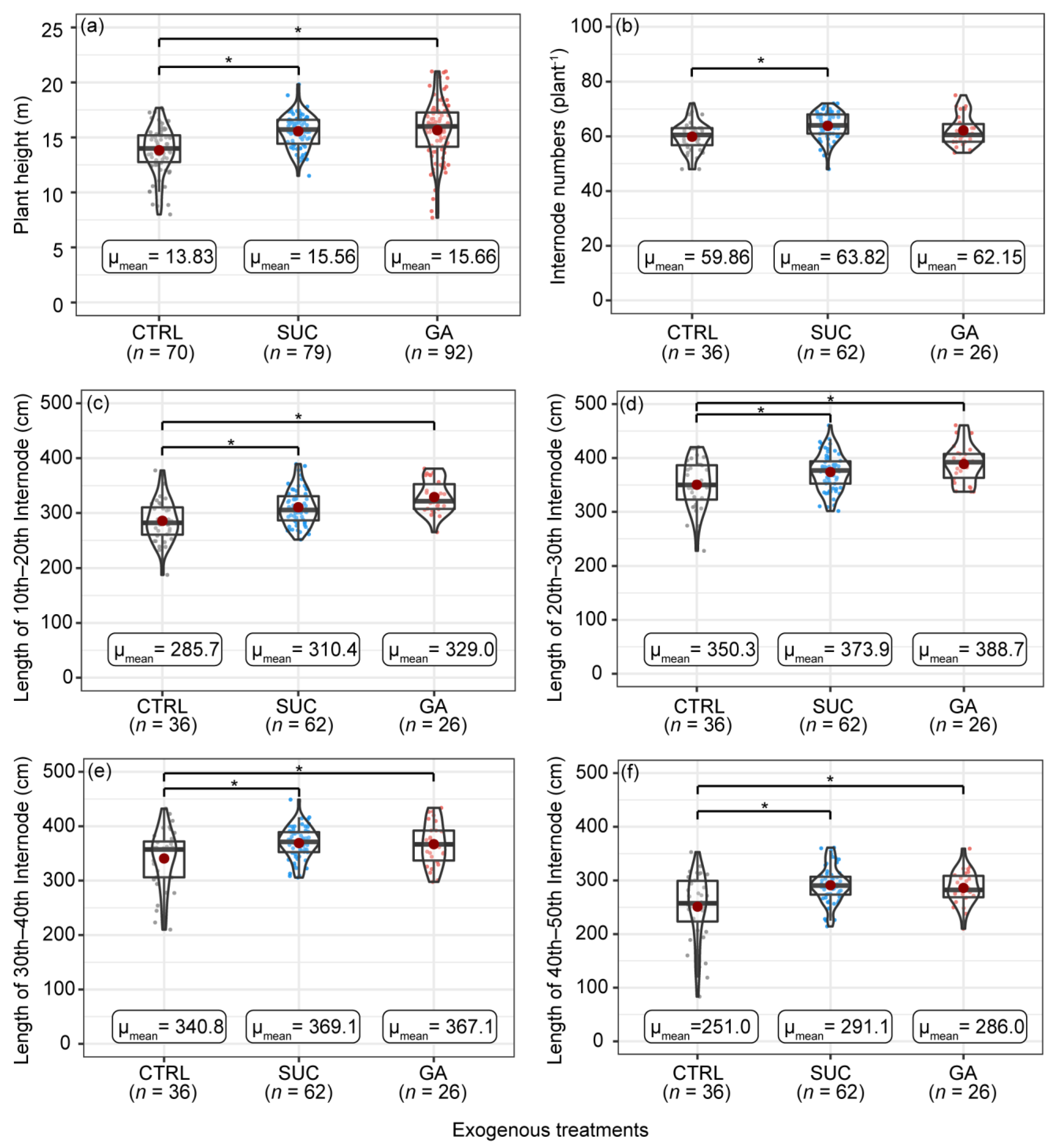
2.1. Effects of Exogenous Treatments on the Plant Height, Internode Numbers, and Internode Lengths
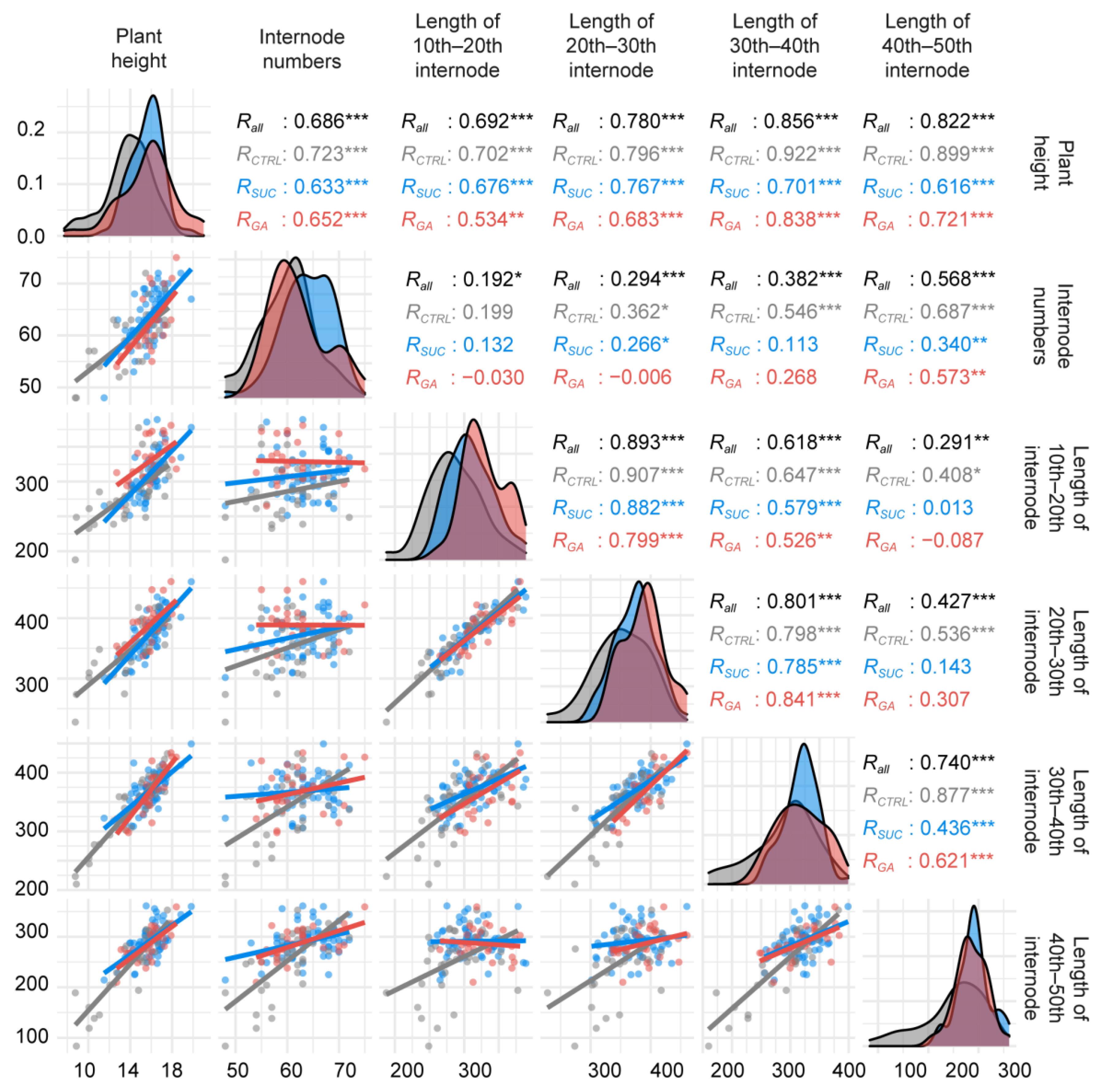
2.2. Data Characteristics and Correlations between Variables
2.3. Effects of Exogenous Treatments on the Internode Lengths
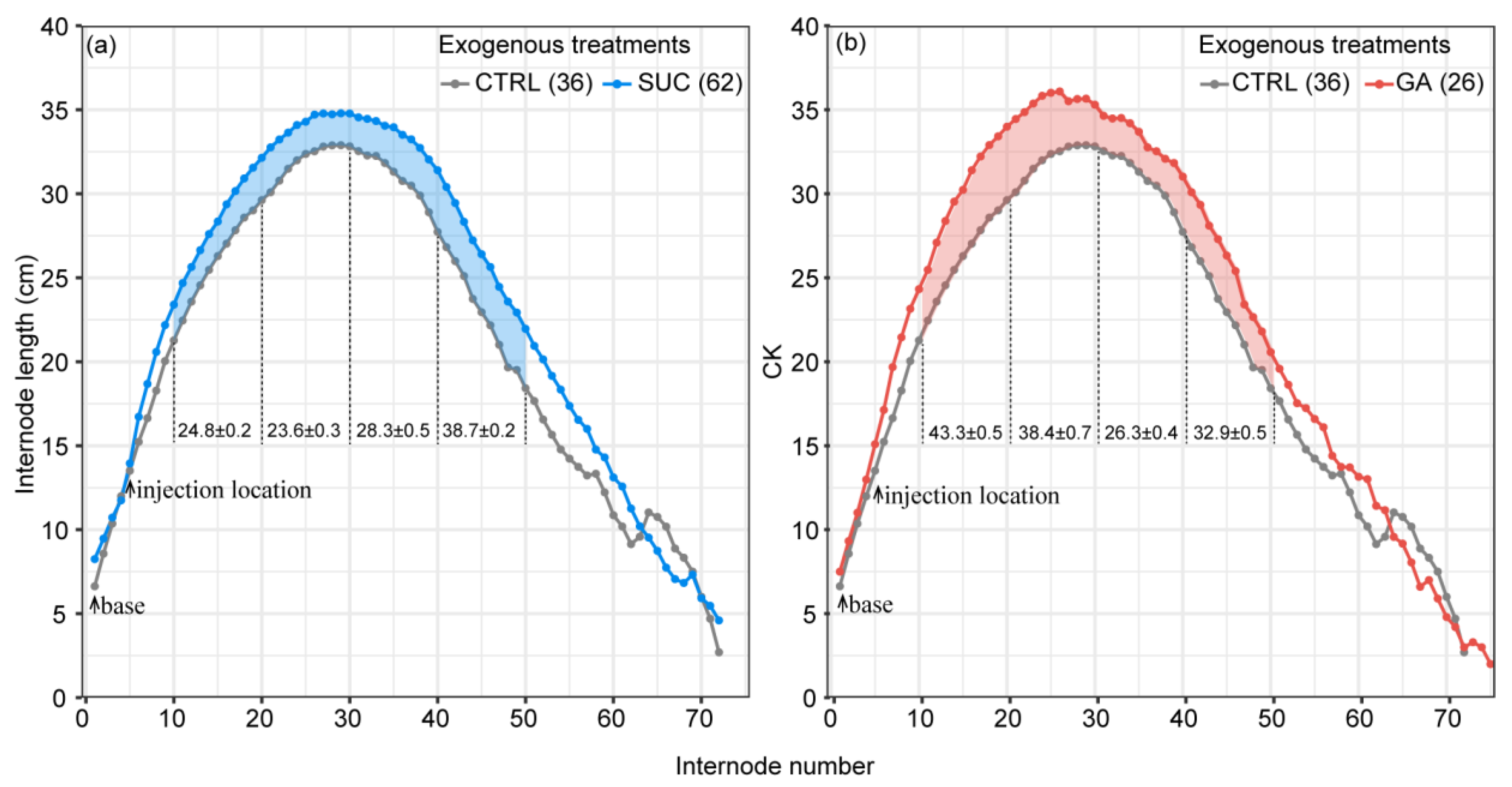
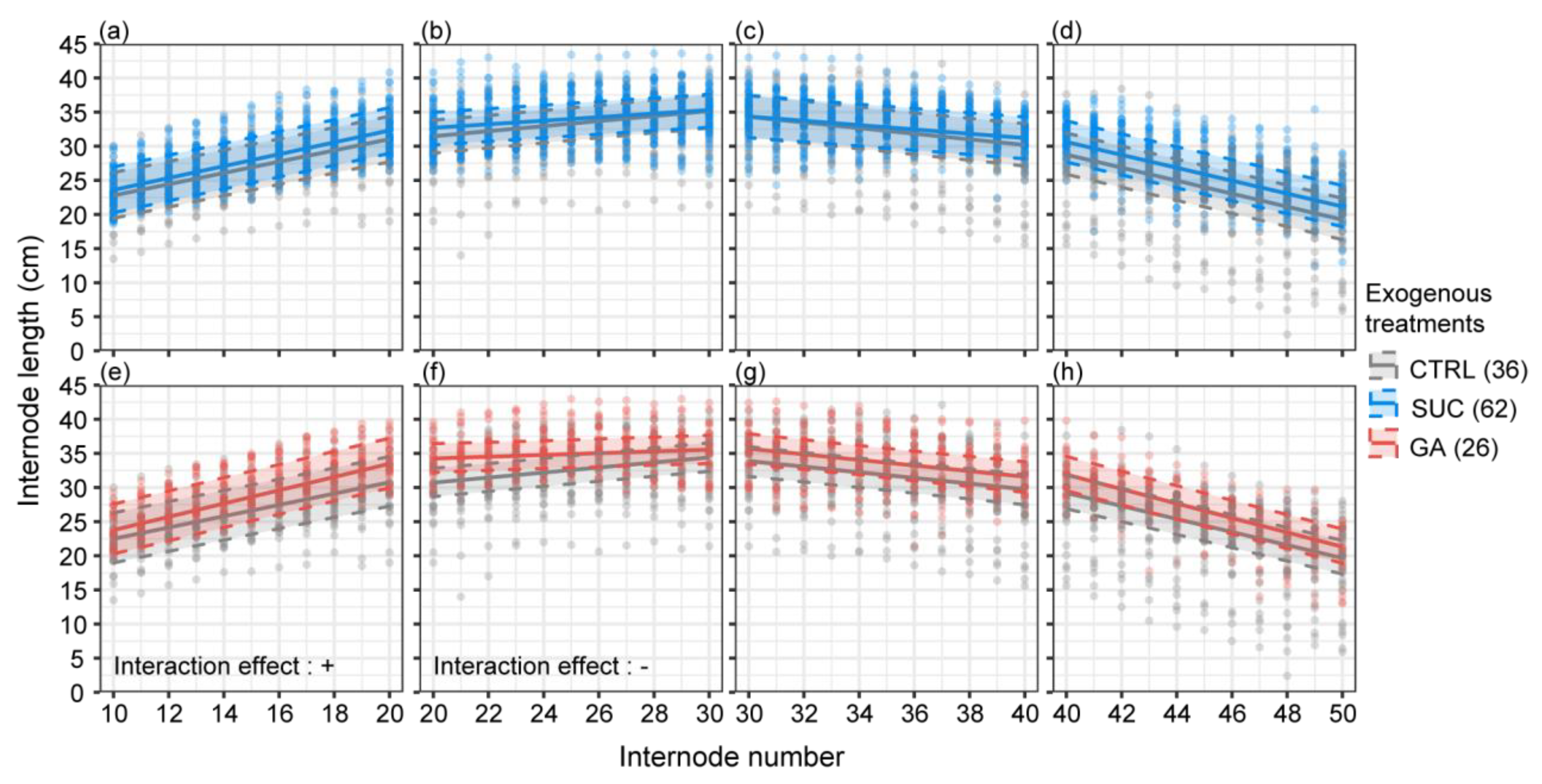
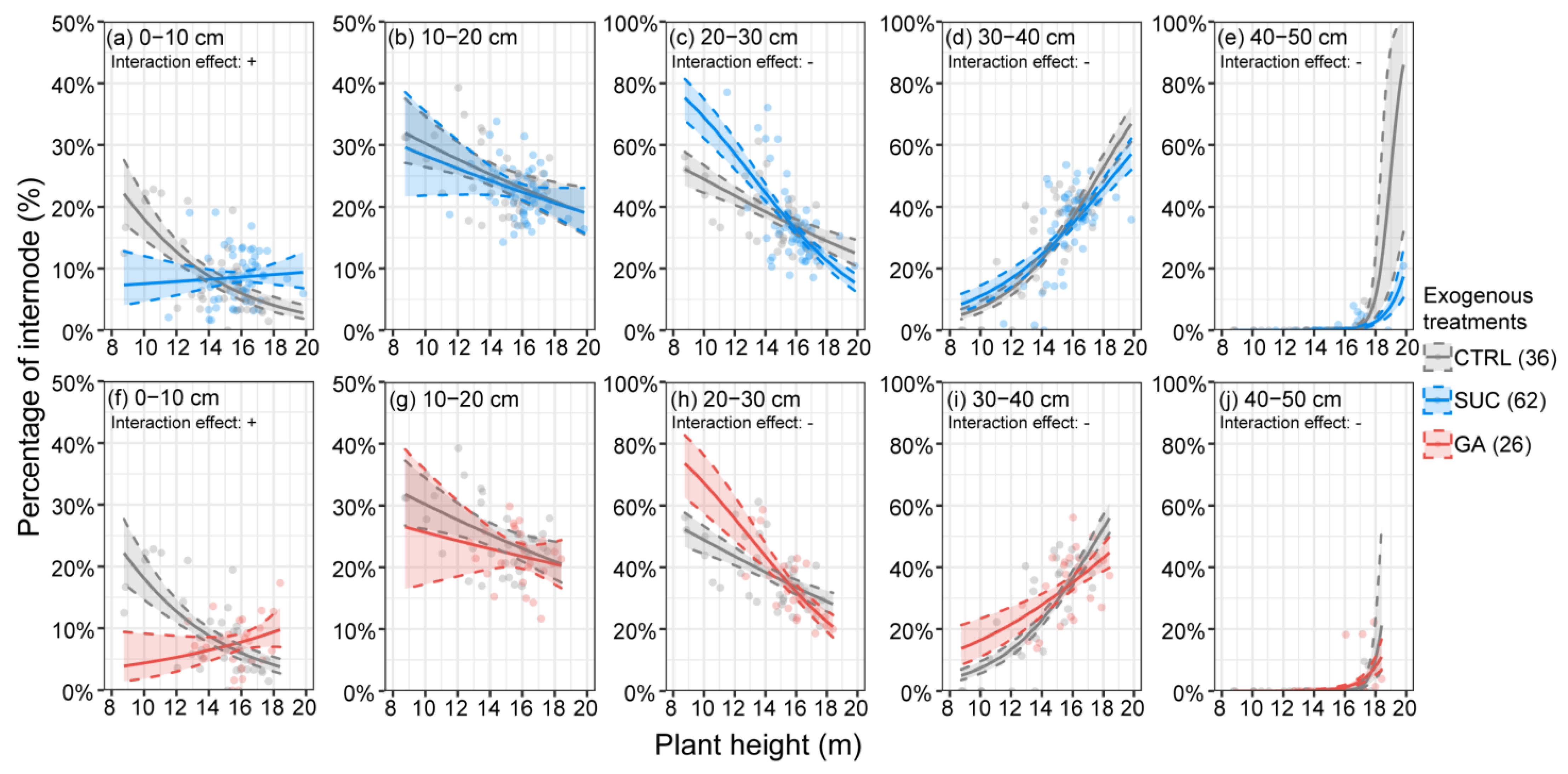
2.4. Variation in Promotion of Internode Elongation under Exogenous Treatments with Distance
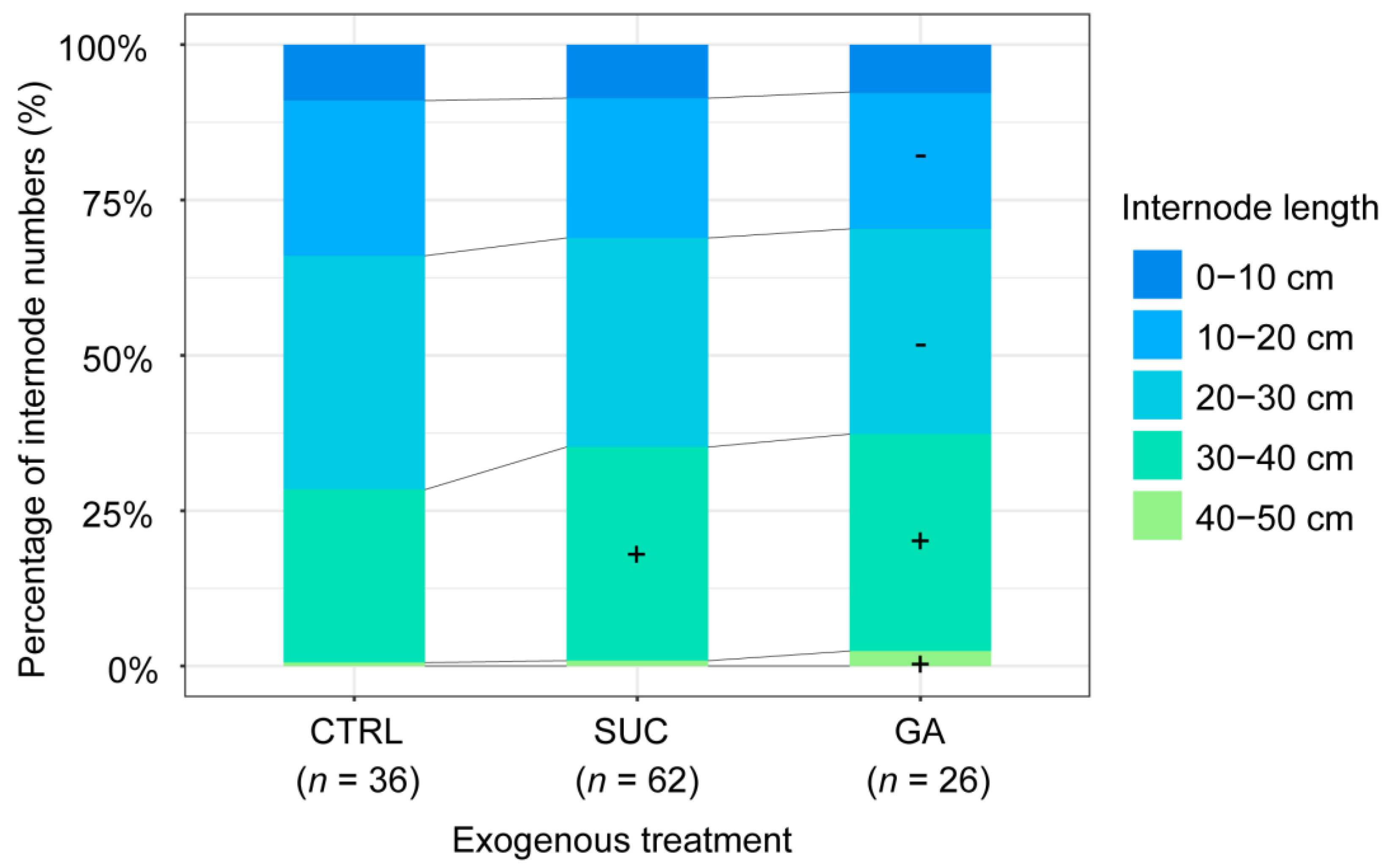
2.5. Effects of Exogenous Treatments on the Percentage of Internodes with Different Lengths
2.6. Effects of Exogenous Treatments on the Percentages of Internodes at Different Plant Heights
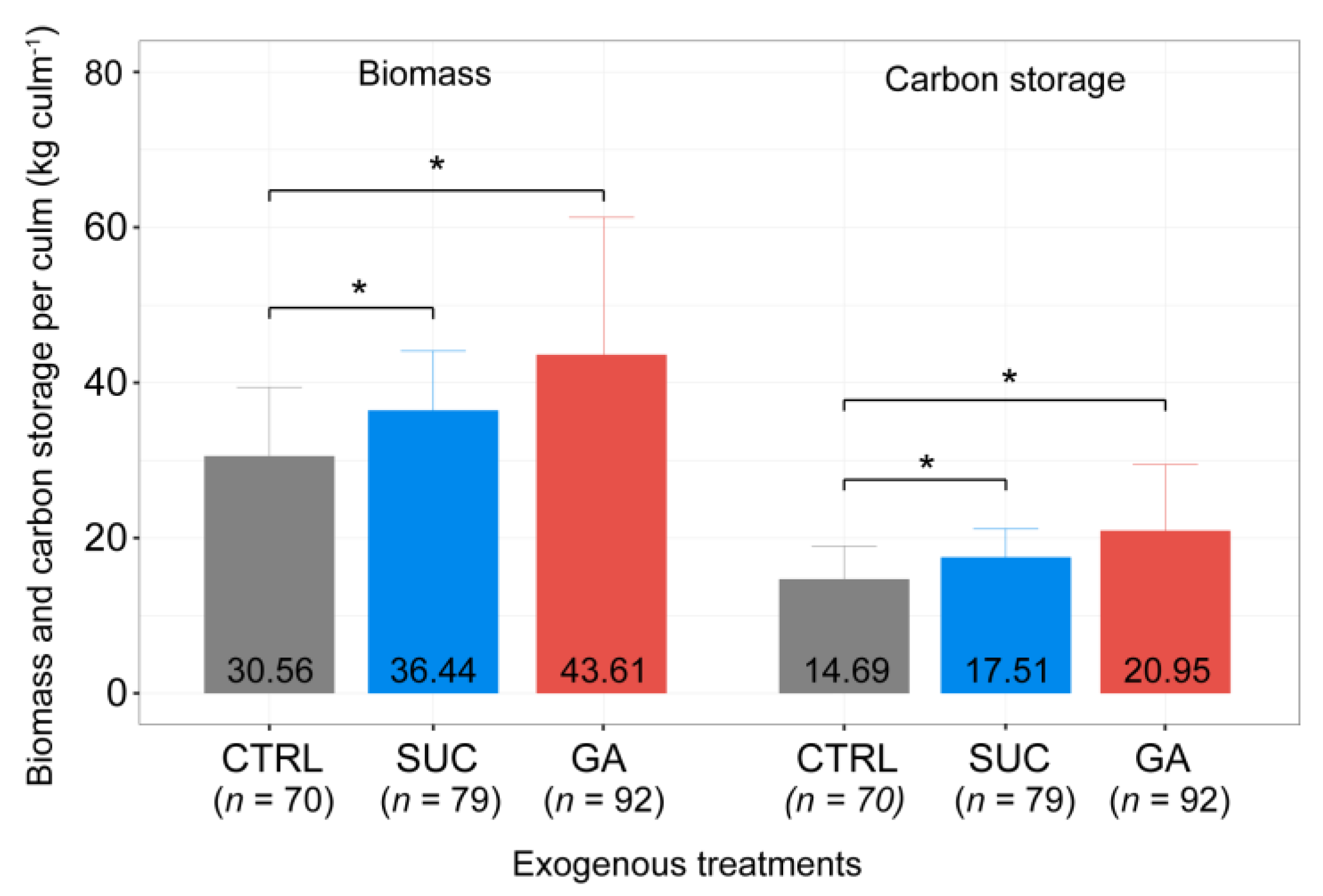
2.7. Effects of Exogenous Treatments on Ecological Benefits
3. Discussion
3.1. Exogenous Treatments Did Not Affect the Characteristics of the Culm Morphological and Internode Length Distributions
3.2. Effects of the Exogenous Treatments on the Internode Lengths
3.3. Effects of the Exogenous Treatments on the Internode Numbers
3.4. Effects of Exogenous Treatments on the Plant Height and Ecological Benefits
4. Materials and Methods
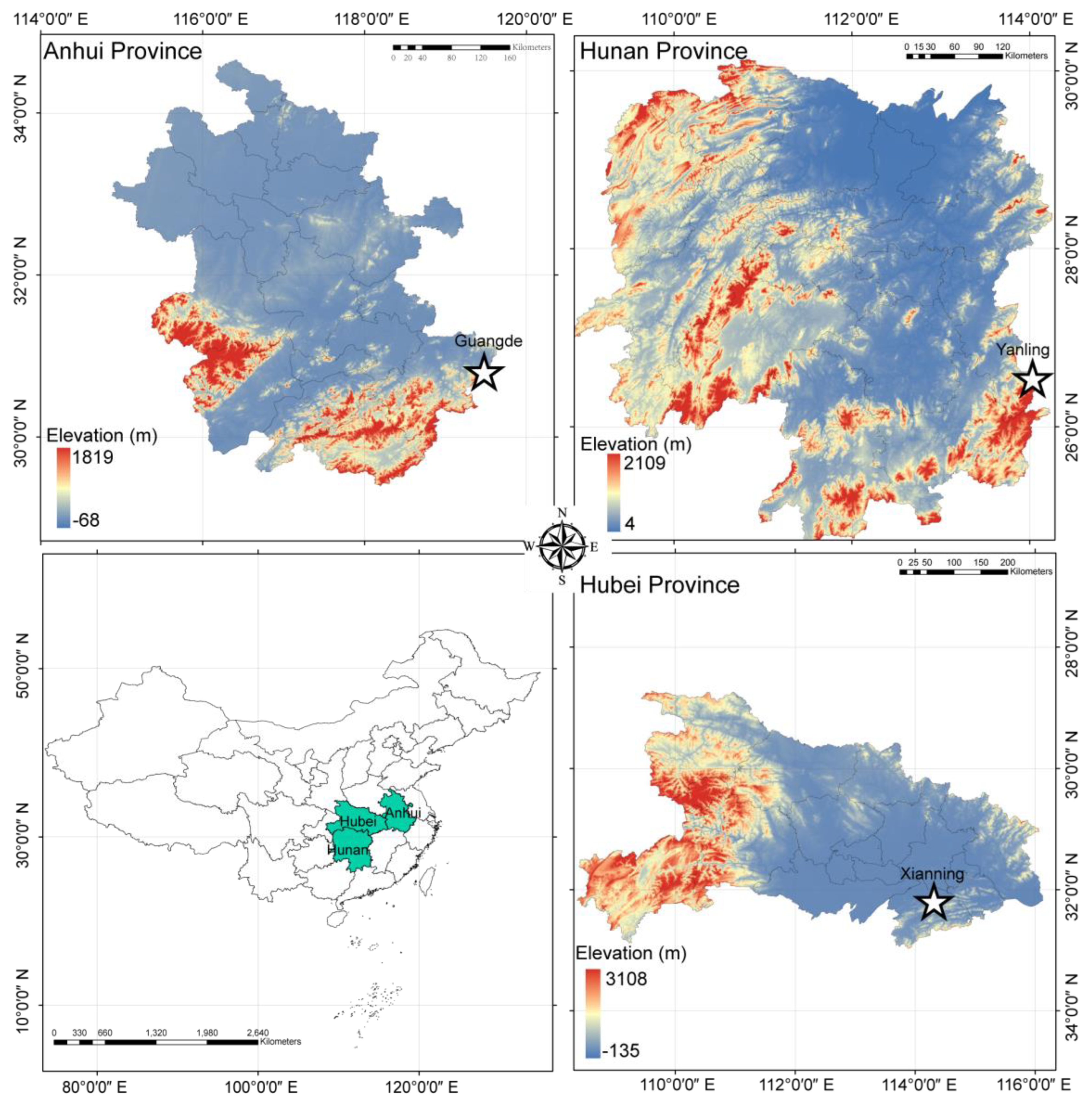
4.1. Study Site and Experiment Design
4.2. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, Y. Bamboos in the Asia Pacific Region; Lessard, G., Chouinard, A., Eds.; International Development Research Centre: Ottawa, ON, Canada, 1980. [Google Scholar]
- Scurlock, J.M.; Dayton, D.C.; Hames, B. Bamboo: An overlooked biomass resource? Biomass Bioenergy 2000, 19, 229–244. [Google Scholar] [CrossRef]
- Zhou, B.Z.; Fu, M.Y.; Xie, J.Z.; Yang, X.S.; Li, Z.C. Ecological functions of bamboo forest: Research and Application. J. For. Res. 2005, 16, 143–147. [Google Scholar] [CrossRef]
- Song, X.Z.; Zhou, G.M.; Jiang, H.; Yu, S.Q.; Fu, J.H.; Li, W.Z.; Wang, W.F.; Ma, Z.H.; Peng, C.H. Carbon sequestration by Chinese bamboo forests and their ecological benefits: Assessment of potential, problems, and future challenges. Environ. Rev. 2011, 19, 418–428. [Google Scholar] [CrossRef]
- Choudhury, D.; Sahu, J.K.; Sharma, G.D. Value addition to bamboo shoots: A review. J. Food Sci. Technol. 2012, 49, 407–414. [Google Scholar] [CrossRef]
- Sawarkar, A.D.; Shrimankar, D.D.; Kumar, A.; Kumar, A.; Singh, E.; Singh, L.; Kumar, S.; Kumar, R. Commercial clustering of sustainable bamboo species in India. Ind. Crops Prod. 2020, 154, 112693. [Google Scholar] [CrossRef]
- Gao, J. The Moso Bamboo Genome; Springer International Publishing: Cham, Switzerland, 2021. [Google Scholar]
- Buziquia, S.T.; Lopes, P.V.F.; Almeida, A.K.; de Almeida, I.K. Impacts of bamboo spreading: A review. Biodivers. Conserv. 2019, 28, 3695–3711. [Google Scholar] [CrossRef]
- Xu, Q.F.; Liang, C.F.; Chen, J.H.; Li, Y.C.; Qin, H.; Fuhrmann, J.J. Rapid bamboo invasion (expansion) and its effects on biodiversity and soil processes +. Glob. Ecol. Conserv. 2020, 21, e00787. [Google Scholar] [CrossRef]
- Ahmad, Z.; Upadhyay, A.; Ding, Y.L.; Emamverdian, A.; Shahzad, A. Bamboo: Origin, habitat, distributions and global prospective. In Biotechnological Advances in Bamboo; Ahmad, Z., Ding, Y., Shahzad, A., Eds.; Springer: Singapore, 2021; pp. 1–31. [Google Scholar]
- Chen, S.L.; Li, D.Z.; Zhu, G.H.; Wu, Z.L.; Lu, S.L.; Liu, L.; Wang, Z.P.; Wang, C.P.; Sun, B.X.; Sun, B.S.; et al. Flora of China; Science Press: Beijing, China, 2018; Volume 22. [Google Scholar]
- State Forestry and Grassland Administration of China. Report of Forest Resources in China (2014–2018); China Forestry Publishing House: Beijing, China, 2019. [Google Scholar]
- INBAR. China Bamboo and Rattan Commodity International Trade Report; Wu, J.Q., Wang, X.D., Eds.; International Bamboo and Rattan Organisation: Beijing, China, 2020. [Google Scholar]
- Dixon, P.G.; Gibson, L.J. The structure and mechanics of moso bamboo material. J. R. Soc. Interface 2014, 11, 20140321. [Google Scholar] [CrossRef]
- Chen, M.L.; Ye, L.; Li, H.; Wang, G.; Chen, Q.; Fang, C.H.; Dai, C.P.; Fei, B.H. Flexural strength and ductility of moso bamboo. Constr. Build. Mater. 2020, 246, 118418. [Google Scholar] [CrossRef]
- Fang, C.H.; Jiang, Z.H.; Sun, Z.J.; Liu, H.R.; Zhang, X.B.; Zhang, R.; Fei, B.H. An overview on bamboo culm flattening. Constr. Build. Mater. 2018, 171, 65–74. [Google Scholar] [CrossRef]
- Li, Y.J.; Lou, Z.C.; Jiang, Y.J.; Wang, X.Z.; Yuan, T.C.; Yang, M.F. Flattening technique without nicked in curved bamboo strips. For. Mach Woodwork Equip. 2020, 48, 28–30+34. [Google Scholar] [CrossRef]
- Li, Z.; Chen, C.; Mi, R.; Gan, W.; Dai, J.; Jiao, M.; Xie, H.; Yao, Y.; Xiao, S.; Hu, L. A strong, tough, and scalable structural material from fast-growing bamboo. Adv. Mater. 2020, 32, e1906308. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.C.; Wang, Q.Y.; Sun, W.; Zhao, Y.H.; Wang, X.Z.; Liu, X.R.; Li, Y.J. Bamboo flattening technique: A literature and patent review. Eur. J. Wood Wood Prod. 2021, 79, 1035–1048. [Google Scholar] [CrossRef]
- Muktar, M.S.; Habte, E.; Teshome, A.; Assefa, Y.; Negawo, A.T.; Lee, K.W.; Zhang, J.; Jones, C.S. Insights into the genetic architecture of complex traits in napier grass (Cenchrus purpureus) and QTL regions governing forage biomass yield, water use efficiency and feed quality traits. Front. Plant Sci. 2021, 12, 678862. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Li, J.; Lu, L.Y.; Gao, L.J.; Lai, D.W.; Yao, N.; Yi, X.F.; Wu, Z.Y.; Lai, Z.Q.; Zhang, J.Y. Integrated analyses of phenotype, phytohormone, and transcriptome to elucidate the mechanism governing internode elongation in two contrasting elephant grass (Cenchrus purpureus) cultivars. Ind. Crops Prod. 2021, 170, 113693. [Google Scholar] [CrossRef]
- Wind, J.; Smeekens, S.; Hanson, J. Sucrose: Metabolite and signaling molecule. Phytochemistry 2010, 71, 1610–1614. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Aluko, O.O.; Li, C.; Wang, Q.; Liu, H. Sucrose utilization for improved crop yields: A review article. Int. J. Mol. Sci. 2021, 22, 4704. [Google Scholar] [CrossRef] [PubMed]
- Doehlert, D.C.; Kuo, T.M.; Felker, F.C. Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize. Plant Physiol. 1988, 86, 1013–1019. [Google Scholar] [CrossRef]
- Patrick, J.W. Does Don Fisher’s high-pressure manifold model account for phloem transport and resource partitioning? Front. Plant Sci. 2013, 4, 184. [Google Scholar] [CrossRef]
- Kebrom, T.H.; Mullet, J.E. Photosynthetic leaf area modulates tiller bud outgrowth in sorghum. Plant Cell Environ. 2015, 38, 1471–1478. [Google Scholar] [CrossRef]
- Xia, X.; Dong, H.; Yin, Y.; Song, X.; Gu, X.; Sang, K.; Zhou, J.; Shi, K.; Zhou, Y.; Foyer, C.H.; et al. Brassinosteroid signaling integrates multiple pathways to release apical dominance in tomato. Proc. Natl. Acad. Sci. USA 2021, 118, e2004384118. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.Y.; Yang, M.T.; Chen, S.Y.; Zhou, Z.Q.; Li, X.; Wang, X.J.; Bai, J.G. Exogenous sucrose influences antioxidant enzyme activities and reduces lipid peroxidation in water-stressed cucumber leaves. Biol. Plant. 2015, 59, 147–153. [Google Scholar] [CrossRef]
- Zhou, Z.P.; Yuan, Y.Z.; Zhou, W.; Zhang, C.F. Effects of exogenously supplied sucrose on OsSUTs and OsSPSs transcript abundances and rice root ammonium assimilation. Acta Physiol. Plant. 2016, 38, 274. [Google Scholar] [CrossRef]
- Medeiros, D.B.; Perez Souza, L.; Antunes, W.C.; Araujo, W.L.; Daloso, D.M.; Fernie, A.R. Sucrose breakdown within guard cells provides substrates for glycolysis and glutamine biosynthesis during light-induced stomatal opening. Plant J. 2018, 94, 583–594. [Google Scholar] [CrossRef]
- Ishioka, N.; Tanimoto, S.; Harada, H. Roles of nitrogen and carbohydrate in floral-bud formation in Pharbitis Apex Cultures. J. Plant Physiol. 1991, 138, 573–576. [Google Scholar] [CrossRef]
- Donnison, I.S.; Francis, D. Experimental control of floral reversion in isolated shoot apices of the long-day plant Silene coeli-rosa. Physiol. Plant. 1994, 92, 329–335. [Google Scholar] [CrossRef]
- Wang, X.; Geng, X.; Yang, L.; Chen, Y.; Zhao, Z.; Shi, W.; Kang, L.; Wu, R.; Lu, C.; Gao, J. Total and mitochondrial transcriptomic and proteomic insights into regulation of bioenergetic processes for shoot fast-growth initiation in moso bamboo. Cells 2022, 11, 1240. [Google Scholar] [CrossRef]
- Zheng, H.F.; Cai, M.M.; Bai, Y.C.; Xu, J.L.; Xie, Y.L.; Song, H.J.; Li, J.; Gao, J. The effect of guttation on the growth of bamboo shoots. Forests 2021, 13, 31. [Google Scholar] [CrossRef]
- Wang, S.; Pei, J.; Li, J.; Tang, G.; Zhao, J.; Peng, X.; Nie, S.; Ding, Y.; Wang, C. Sucrose and starch metabolism during Fargesia yunnanensis shoot growth. Physiol. Plant. 2020, 168, 188–204. [Google Scholar] [CrossRef]
- Wei, Q.; Guo, L.; Jiao, C.; Fei, Z.; Chen, M.; Cao, J.; Ding, Y.; Yuan, Q. Characterization of the developmental dynamics of the elongation of a bamboo internode during the fast growth stage. Tree Physiol. 2019, 39, 1201–1214. [Google Scholar] [CrossRef]
- Hori, S. Some observations on ‘Bakanae’disease of the rice plant. Mem. Agric. Res. Sta. 1898, 12, 110–119. [Google Scholar]
- Randoux, M.; Jeauffre, J.; Thouroude, T.; Vasseur, F.; Hamama, L.; Juchaux, M.; Sakr, S.; Foucher, F. Gibberellins regulate the transcription of the continuous flowering regulator, RoKSN, a rose TFL1 homologue. J. Exp. Bot. 2012, 63, 6543–6554. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Sponsel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, D.; Fan, S.; Du, L.; Shen, Y.; Xing, L.; Li, Y.; Ma, J.; Han, M. Effect of exogenous GA3 and its inhibitor paclobutrazol on floral formation, endogenous hormones, and flowering-associated genes in ‘Fuji’ apple (Malus domestica Borkh.). Plant Physiol. Biochem. 2016, 107, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Stowe, B.B.; Yamaki, T. The history and physiological action of the gibberellins. Annu. Rev. Plant Physiol. 1957, 8, 181–216. [Google Scholar] [CrossRef]
- Yan, Q.; Li, J.; Lu, L.Y.; Yi, X.F.; Yao, N.; Lai, Z.Q.; Zhang, J.Y. Comparative transcriptome study of the elongating internode in elephant grass (Cenchrus purpureus) seedlings in response to exogenous gibberellin applications. Ind. Crops Prod. 2022, 178, 114653. [Google Scholar] [CrossRef]
- Koornneef, M.; van der Veen, J.H. Induction and analysis of gibberellin sensitive mutants in Arabidopsis thaliana (L.) heynh. Theor. Appl. Genet. 1980, 58, 257–263. [Google Scholar] [CrossRef]
- Fu, X.; Harberd, N.P. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature 2003, 421, 740–743. [Google Scholar] [CrossRef]
- Liao, Z.; Yu, H.; Duan, J.; Yuan, K.; Yu, C.; Meng, X.; Kou, L.; Chen, M.; Jing, Y.; Liu, G.; et al. SLR1 inhibits MOC1 degradation to coordinate tiller number and plant height in rice. Nat. Commun. 2019, 10, 2738. [Google Scholar] [CrossRef]
- Chen, Y.; Hou, M.; Liu, L.; Wu, S.; Shen, Y.; Ishiyama, K.; Kobayashi, M.; McCarty, D.R.; Tan, B.C. The maize DWARF1 encodes a gibberellin 3-oxidase and is dual localized to the nucleus and cytosol. Plant Physiol. 2014, 166, 2028–2039. [Google Scholar] [CrossRef]
- Ishii, Y.; Yamano, A.; Idota, S. Effects of short-day and gibberellic acid treatments on summer vegetative propagation of napier grass (Pennisetum purpureum Schumach). Int. J. Agron. 2016, 2016, 9606914. [Google Scholar] [CrossRef]
- Zhang, X.; He, L.; Zhao, B.; Zhou, S.; Li, Y.; He, H.; Bai, Q.; Zhao, W.; Guo, S.; Liu, Y.; et al. Dwarf and Increased Branching 1 controls plant height and axillary bud outgrowth in Medicago truncatula. J. Exp. Bot. 2020, 71, 6355–6365. [Google Scholar] [CrossRef] [PubMed]
- Rebetzke, G.J.; Richards, R.A. Gibberellic acid-sensitive dwarfing genes reduce plant height to increase kernel number and grain yield of wheat. Aust. J. Agric. Res. 2000, 51, 235–246. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet 2007, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Du, Y.Y.; Yang, Z.Y.; Chen, L.; Condon, A.G.; Hu, Y.G. Comparing the effects of GA-responsive dwarfing genes Rht13 and Rht8 on plant height and some agronomic traits in common wheat. Field Crops Res. 2015, 179, 35–43. [Google Scholar] [CrossRef]
- Tian, X.; Xia, X.; Xu, D.; Liu, Y.; Xie, L.; Hassan, M.A.; Song, J.; Li, F.; Wang, D.; Zhang, Y.; et al. Rht24b, an ancient variation of TaGA2ox-A9, reduces plant height without yield penalty in wheat. New Phytol. 2022, 233, 738–750. [Google Scholar] [CrossRef]
- Mitsunaga, S.; Tashiro, T.; Yamaguchi, J. Identification and characterization of gibberellin-insensitive mutants selected from among dwarf mutants of rice. Theor. Appl. Genet. 1994, 87, 705–712. [Google Scholar] [CrossRef]
- Qiao, F.; Yang, Q.; Wang, C.L.; Fan, Y.L.; Wu, X.F.; Zhao, K.J. Modification of plant height via RNAi suppression of OsGA20ox2 gene in rice. Euphytica 2007, 158, 35–45. [Google Scholar] [CrossRef]
- Voorend, W.; Nelissen, H.; Vanholme, R.; De Vliegher, A.; Van Breusegem, F.; Boerjan, W.; Roldan, R.I.; Muylle, H.; Inze, D. Overexpression of GA20-OXIDASE1 impacts plant height, biomass allocation and saccharification efficiency in maize. Plant Biotechnol. J. 2016, 14, 997–1007. [Google Scholar] [CrossRef]
- Hu, S.; Wang, C.; Sanchez, D.L.; Lipka, A.E.; Liu, P.; Yin, Y.; Blanco, M.; Lubberstedt, T. Gibberellins promote brassinosteroids action and both increase heterosis for plant height in maize (Zea mays L.). Front. Plant Sci. 2017, 8, 1039. [Google Scholar] [CrossRef]
- Gebremeskel, H.; Dou, J.; Li, B.; Zhao, S.; Muhammad, U.; Lu, X.; He, N.; Liu, W. Molecular mapping and candidate gene analysis for GA3 responsive short internode in watermelon (Citrullus lanatus). Int. J. Mol. Sci. 2019, 21, 290. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Wu, D.; Wang, C.; Liu, Y.; Wu, D. Characterization of the molecular mechanism underlying the dwarfism of dsh mutant watermelon plants. Plant Sci. Lett. 2021, 313, 111074. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cheng, Z.; Ma, Y.; Bai, Q.; Li, X.; Cao, Z.; Wu, Z.; Gao, J. The association of hormone signalling genes, transcription and changes in shoot anatomy during moso bamboo growth. Plant Biotechnol. J. 2018, 16, 72–85. [Google Scholar] [CrossRef]
- Hou, D.; Bai, Q.S.; Li, J.; Xie, L.H.; Li, X.Y.; Cheng, Z.C.; Gao, J. The gibberellic acid-stimulated transcript gene family in moso bamboo: A genome-wide survey and expression profiling during development and abiotic stresses. J. Plant Growth Regul. 2018, 37, 1135–1147. [Google Scholar] [CrossRef]
- Cai, M.; Cheng, W.; Bai, Y.; Mu, C.; Zheng, H.; Cheng, Z.; Gao, J. PheGRF4e initiated auxin signaling during moso bamboo shoot development. Mol. Biol. Rep. 2022, 49, 8815–8825. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, L.; Wang, X.; Liang, L.; Yue, J.; Li, L. Comparative analyses of anatomical structure, phytohormone levels, and gene expression profiles reveal potential dwarfing mechanisms in shengyin bamboo (Phyllostachys edulis f. tubaeformis). Int. J. Mol. Sci. 2018, 19, 1697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, H.; Zhu, Q.; Gao, Y.; Wang, H.; Zhao, L.; Wang, Y.; Xi, F.; Wang, W.; Yang, Y.; et al. Transcriptome characterization of moso bamboo (Phyllostachys edulis) seedlings in response to exogenous gibberellin applications. BMC Plant Biol. 2018, 18, 125. [Google Scholar] [CrossRef]
- Chen, M.; Guo, L.; Ramakrishnan, M.; Fei, Z.; Vinod, K.K.; Ding, Y.; Jiao, C.; Gao, Z.; Zha, R.; Wang, C.; et al. Rapid growth of moso bamboo (Phyllostachys edulis): Cellular roadmaps, transcriptome dynamics, and environmental factors. Plant Cell 2022, 34, 3577–3610. [Google Scholar] [CrossRef]
- Isagi, Y.; Torii, A. Range expansion and its mechanisms in a naturalized bamboo species, Phyllostachys pubescens, in Japan. J. Sustain. For. 1997, 6, 127–141. [Google Scholar] [CrossRef]
- Inoue, A.; Kuraoka, K.; Kitahara, F. Mathematical expression for the relationship between internode number and internode length for bamboo, Phyllostachys pubescens. J. For. Res. 2012, 23, 435–439. [Google Scholar] [CrossRef]
- Inoue, A. Culm form analysis for bamboo, Phyllostachys pubescens. J. For. Res. 2013, 24, 525–530. [Google Scholar] [CrossRef]
- Gao, X.; Li, Z.D.; Yu, H.M.; Jiang, Z.H.; Wang, C.; Zhang, Y.; Qi, L.H.; Shi, L. Modeling of the height–diameter relationship using an allometric equation model: A case study of stands of Phyllostachys edulis. J. For. Res. 2015, 27, 339–347. [Google Scholar] [CrossRef]
- Yen, T.M. Culm height development, biomass accumulation and carbon storage in an initial growth stage for a fast-growing moso bamboo (Phyllostachy pubescens). Bot. Stud. 2016, 57, 10. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Suga, H. Relationships of culm surface area to other culm dimensions for bamboo, Phyllostachys pubescens. J. For. Res. 2017, 14, 236–239. [Google Scholar] [CrossRef]
- Suga, H.; Inoue, A.; Kitahara, F. Derivation of two-way volume equation for bamboo, Phyllostachys pubescens. J. For. Res. 2017, 16, 261–267. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, P.; Zhang, Z.; Zhu, L.; Hu, Y.; Ouyang, L.; Ni, G.; Ye, Q. Culm age and rhizome affects night-time water recharge in the bamboo Phyllostachys pubescens. Front. Plant Sci. 2017, 8, 1928. [Google Scholar] [CrossRef]
- Inoue, A.; Sato, M.; Shima, H. A new taper index based on form-factor: Application to three bamboo species (Phyllostachys spp.). Eur. J. For. Res. 2021, 140, 1533–1542. [Google Scholar] [CrossRef]
- Banik, R.L. Morphology and growth. In Bamboo; Springer: Berlin/Heidelberg, Germany, 2015; pp. 43–89. [Google Scholar]
- Zhou, Z.S.; Tang, Y.; Xu, H.X.; Wang, J.Z.; Hu, L.L.; Xu, X.J. Dynamic changes in leaf biomass and the modeling of individual moso bamboo (Phyllostachys edulis (Carrière) J. Houz) under intensive management. Forests 2022, 13, 693. [Google Scholar] [CrossRef]
- Galaud, J.-P.; Gaspar, T.; Boyer, N. Inhibition of internode growth due to mechanical stress in Bryonia dioica: Relationship between changes in DNA methylation and ethylene metabolism. Physiol. Plant. 1993, 87, 25–30. [Google Scholar] [CrossRef]
- Valinger, E.; Lundqvist, L.; Sundberg, B. Mechanical stress during dormancy stimulates stem growth of Scots pine seedlings. For. Ecol. Manag. 1994, 67, 299–303. [Google Scholar] [CrossRef]
- Takano, M.; Takahashi, H.; Suge, H. Mechanical stress and gibberellin: Regulation of hollowing induction in the stem of a bean plant, Phaseolus vulgaris L. Plant Cell Physiol. 1995, 36, 101–108. [Google Scholar] [CrossRef]
- Huber, H.; de Brouwer, J.; von Wettberg, E.J.; During, H.J.; Anten, N.P.R. More cells, bigger cells or simply reorganization? Alternative mechanisms leading to changed internode architecture under contrasting stress regimes. New Phytol. 2014, 201, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Werger, M.; During, H.; Zhong, Z.C. Carbon and nutrient dynamics in relation to growth rhythm in the giant bamboo Phyllostachys pubescens. Plant Soil 1998, 201, 113–123. [Google Scholar] [CrossRef]
- Zhou, G.; Jiang, P.; Xu, Q. Carbon Fixing and Transition in the Ecosystem of Bamboo Stands; Science Press: Beijing, China, 2010. [Google Scholar]
- O’Toole, J.C.; Cruz, R.T.; Singh, T.N. Leaf rolling and transpiration. Plant Sci. Lett. 1979, 16, 111–114. [Google Scholar] [CrossRef]
- Wheeler, T.D.; Stroock, A.D. The transpiration of water at negative pressures in a synthetic tree. Nature 2008, 455, 208–212. [Google Scholar] [CrossRef]
- Vadez, V.; Kholova, J.; Medina, S.; Kakkera, A.; Anderberg, H. Transpiration efficiency: New insights into an old story. J. Exp. Bot. 2014, 65, 6141–6153. [Google Scholar] [CrossRef]
- Peel, A.J. Transport of Nutrients in Plants; Elsevier: London, UK, 2013. [Google Scholar]
- Song, X.; Peng, C.; Zhou, G.; Gu, H.; Li, Q.; Zhang, C. Dynamic allocation and transfer of non-structural carbohydrates, a possible mechanism for the explosive growth of moso bamboo (Phyllostachys heterocycla). Sci. Rep. 2016, 6, 25908. [Google Scholar] [CrossRef]
- Kende, H.; van der Knaap, E.; Cho, H.T. Deepwater rice: A model plant to study stem elongation. Plant Physiol. 1998, 118, 1105–1110. [Google Scholar] [CrossRef]
- Kaneko, M.; Itoh, H.; Inukai, Y.; Sakamoto, T.; Ueguchi, M.; Ashikari, M.; Matsuoka, M. Where do gibberellin biosynthesis and gibberellin signaling occur in rice plants? Plant J. 2003, 35, 104–115. [Google Scholar] [CrossRef]
- Nagai, K.; Mori, Y.; Ishikawa, S.; Furuta, T.; Gamuyao, R.; Niimi, Y.; Hobo, T.; Fukuda, M.; Kojima, M.; Takebayashi, Y.; et al. Antagonistic regulation of the gibberellic acid response during stem growth in rice. Nature 2020, 584, 109–114. [Google Scholar] [CrossRef]
- Qazi, H.A.; Paranjpe, S.; Bhargava, S. Stem sugar accumulation in sweet sorghum—Activity and expression of sucrose metabolizing enzymes and sucrose transporters. J. Plant Physiol. 2012, 169, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Zhu, Z.; Wang, W.; Cai, J.; Chen, Y.; Li, L.; Tian, S. A tomato vacuolar invertase inhibitor mediates sucrose metabolism and influences fruit ripening. Plant Physiol. 2016, 172, 1596–1611. [Google Scholar] [CrossRef] [PubMed]
- Robaglia, C.; Thomas, M.; Meyer, C. Sensing nutrient and energy status by SnRK1 and TOR kinases. Curr. Opin. Plant Biol. 2012, 15, 301–307. [Google Scholar] [CrossRef]
- Fang, D.; Mei, T.; Roll, A.; Holscher, D. Water transfer between bamboo culms in the period of sprouting. Front. Plant Sci. 2019, 10, 786. [Google Scholar] [CrossRef] [PubMed]
- Shan, F.; Zhang, R.; Zhang, J.; Wang, C.; Lyu, X.; Xin, T.; Yan, C.; Dong, S.; Ma, C.; Gong, Z. Study on the regulatory effects of GA3 on soybean internode elongation. Plants 2021, 10, 1737. [Google Scholar] [CrossRef]
- Mendu, N.; Silflow, C.D. Elevated levels of tubulin transcripts accompany the GA3-induced elongation of Oat internode segments. Plant Cell Physiol. 1993, 34, 973–983. [Google Scholar] [CrossRef]
- Xu, N.F.; York, K.; Miller, P.; Cheikh, N. Co-regulation of ear growth and internode elongation in corn. Plant Growth Regul. 2004, 44, 231–241. [Google Scholar] [CrossRef]
- Honi, U.; Amin, M.R.; Kabir, S.M.T.; Bashar, K.K.; Moniruzzaman, M.; Jahan, R.; Jahan, S.; Haque, M.S.; Islam, S. Genome-wide identification, characterization and expression profiling of gibberellin metabolism genes in jute. BMC Plant Biol. 2020, 20, 306. [Google Scholar] [CrossRef]
- Schaik, P.H.; Probst, A.H. Effects of some environmental factors on flower production and reproductive efficiency in soybeans. Agron. J. 1958, 50, 192–197. [Google Scholar] [CrossRef]
- Thomas, J.F.; Raper, C.D. Morphological response of soybeans as governed by photoperiod, temperature, and age at treatment. Bot. Gaz. 1977, 138, 321–328. [Google Scholar] [CrossRef]
- Allen, L.H.; Zhang, L.X.; Boote, K.J.; Hauser, B.A. Elevated temperature intensity, timing, and duration of exposure affect soybean internode elongation, mainstem node number, and pod number per plant. Crop J. 2018, 6, 148–161. [Google Scholar] [CrossRef]
- Misra, V.; Solomon, S.; Mall, A.K.; Prajapati, C.P.; Hashem, A.; Abd Allah, E.F.; Ansari, M.I. Morphological assessment of water stressed sugarcane: A comparison of waterlogged and drought affected crop. Saudi J. Biol. Sci. 2020, 27, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.L.; Shao, Z.P.; Wu, Y.J.; Wu, D. The toughness contribution of bamboo node to the Mode I interlaminar fracture toughness of bamboo. Wood Sci. Technol. 2013, 48, 1257–1268. [Google Scholar] [CrossRef]
- Shima, H.; Sato, M.; Inoue, A. Self-adaptive formation of uneven node spacings in wild bamboo. Phys. Rev. E 2016, 93, 022406. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiao, C.; Guo, L.; Ding, Y.; Cao, J.; Feng, J.; Dong, X.; Mao, L.; Sun, H.; Yu, F.; et al. Exploring key cellular processes and candidate genes regulating the primary thickening growth of moso underground shoots. New Phytol. 2017, 214, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Zhang, Y.H.; Zhang, L.; Li, Y.; Sun, Y.D.; Li, Z.Y. Changes in the distribution of endogenous hormones in Phyllostachys edulis ‘Pachyloen’ during bamboo shooting. PLoS ONE 2020, 15, e0241806. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, R.; Ding, X.; Huang, M.; Shen, K.; Zhao, S.; Xiao, Z.; Xiu, C. Association among starch storage, metabolism, related genes and growth of moso bamboo (Phyllostachys heterocycla) shoots. BMC Plant Biol. 2021, 21, 477. [Google Scholar] [CrossRef]
- Falster, D.S.; Westoby, M. Plant height and evolutionary games. Trends Ecol. Evol. 2003, 18, 337–343. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Bond, B.J.; Meinzer, F.C. Does turgor limit growth in tall trees? Plant Cell Environ. 2004, 27, 229–236. [Google Scholar] [CrossRef]
- Ryan, M.G.; Phillips, N.; Bond, B.J. The hydraulic limitation hypothesis revisited. Plant Cell Environ. 2006, 29, 367–381. [Google Scholar] [CrossRef]
- Niklas, K.J. Maximum plant height and the biophysical factors that limit it. Tree Physiol. 2007, 27, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Li, R.; Werger, M.J.A.; Kroon, H.; During, H.J.; Zhong, Z.C. Interactions between shoot age structure, nutrient availability and physiological integration in the giant bamboo Phyllostachys pubescens. Plant Biol. 2000, 2, 437–446. [Google Scholar] [CrossRef]
- Saitoh, T.; Seiwa, K.; Nishiwaki, A. Importance of physiological integration of dwarf bamboo to persistence in forest understorey: A field experiment. J. Ecol. 2002, 90, 78–85. [Google Scholar] [CrossRef]
- Zhai, W.L.; Wang, Y.; Luan, J.W.; Liu, S.R. Effects of nitrogen addition on clonal integration between mother and daughter ramets of moso bamboo: A 13C-CO2 pulse labeling study. J. Plant Ecol. 2021, 15, 756–770. [Google Scholar] [CrossRef]
- Lu, Z.; Franklin, S.B. Clonal integration and regeneration in bamboo Bashania fargesii. For. Ecol. Manag. 2022, 523, 120504. [Google Scholar] [CrossRef]
- Yen, T.M.; Lee, J.S. Comparing aboveground carbon sequestration between moso bamboo (Phyllostachys heterocycla) and China fir (Cunninghamia lanceolata) forests based on the allometric model. For. Ecol. Manag. 2011, 261, 995–1002. [Google Scholar] [CrossRef]
- Chen, X.G.; Zhang, X.Q.; Zhang, Y.P.; Booth, T.; He, X.H. Changes of carbon stocks in bamboo stands in China during 100 years. For. Ecol. Manag. 2009, 258, 1489–1496. [Google Scholar] [CrossRef]
- Jyoti Nath, A.; Das, G.; Das, A.K. Above ground standing biomass and carbon storage in village bamboos in North East India. Biomass Bioenergy 2009, 33, 1188–1196. [Google Scholar] [CrossRef]
- Wu, W.G.; Liu, Q.; Zhu, Z.; Shen, Y.Q. Managing bamboo for carbon sequestration, bamboo stem and bamboo shoots. Small-Scale For. 2014, 14, 233–243. [Google Scholar] [CrossRef]
- Gupta, A.; Kumar, A. Potential of bamboo in sustainable development. Asia Pac. Bus. Rev. 2008, 4, 100–107. [Google Scholar] [CrossRef]
- Chele, E.S.; Ricardo, M.C.; Ana, P.M.; Teresa, M.R. Bamboo, from traditional crafts to contemporary design and architecture. Procedia Soc. Behav. Sci. 2012, 51, 777–781. [Google Scholar] [CrossRef]
- Manandhar, R.; Kim, J.H.; Kim, J.T. Environmental, social and economic sustainability of bamboo and bamboo-based construction materials in buildings. J. Asian Archit. Build. Eng. 2019, 18, 49–59. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2010: Main Report. FAO Forestry Paper 163; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Liese, W. Research on bamboo. Wood Sci. Technol. 1987, 21, 189–209. [Google Scholar] [CrossRef]
- Liese, W.; Köhl, M. Bamboo: The Plant and Its Uses; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- McNally, A. GES DISC Dataset: FLDAS Noah Land Surface Model L4 Global Monthly 0.1 × 0.1 Degree (MERRA-2 and CHIRPS) (FLDAS_NOAH01_C_GL_M 001); NASA [Data Set]; NASA: Washington, DC, USA, 2018. [Google Scholar] [CrossRef]
- Elmariah, S.; Mauri, L.; Doros, G.; Galper, B.Z.; O’Neill, K.E.; Steg, P.G.; Kereiakes, D.J.; Yeh, R.W. Extended duration dual antiplatelet therapy and mortality: A systematic review and meta-analysis. Lancet 2015, 385, 792–798. [Google Scholar] [CrossRef]
- Feehan, D.M.; Mahmud, A.S. Quantifying population contact patterns in the United States during the COVID-19 pandemic. Nat. Commun. 2021, 12, 893. [Google Scholar] [CrossRef]
- Harrington, S.M.; Wishingrad, V.; Thomson, R.C. Properties of markov chain monte carlo performance across many empirical alignments. Mol. Biol. Evol. 2021, 38, 1627–1640. [Google Scholar] [CrossRef]
- Bürkner, P.C. Brms: An R package for Bayesian multilevel models using stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Hierarchical/Multilevel Models; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Zhou, F.C. Bamboo Forest Cultivation; China Forestry Publishing House: Beijing, China, 1998. [Google Scholar]
- Zhang, H.X.; Zhuang, S.Y.; Sun, B.; Ji, H.B.; Li, C.M.; Zhou, S. Estimation of biomass and carbon storage of moso bamboo (Phyllostachys pubescens Mazel ex Houz.) in southern China using a diameter–age bivariate distribution model. Forestry 2014, 87, 674–682. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Structure | i | j | k | l |
|---|---|---|---|---|
| Phi or Ebi or Eci ~ Normal (μi, σ2i) | 1, 2, …, 241 | 0 or 1 | 1, 2, 3 | |
| μi = β0 + β1Sj + β2Gj + r1i + r2l | ||||
| Ini ~ Poisson (λi) | 1, 2, …, 124 | 0 or 1 | 1, 2, 3 | |
| log(λi) = β0+ β1Sj + β2Gj + r1i+ r2l | ||||
| Il10–20th or 20–30th or 30–40th or 40–50thi ~ Normal (μi, σ2i) | 1, 2, …, 124 | 0 or 1 | 1, 2, 3 | |
| μi = β0 + β1Sj + β2Gj + r1i + r2l | ||||
| Il10–20th or 20–30th or 30–40th or 40–50thi ~ Normal (μi, σ2i) | 1, 2, …, 124 | 0 or 1 | 1, 2, …, 75 | 1, 2, 3 |
| μi = β0 + β1Sj + β2Gj + β3Dk + β4SjDk + β5GjDk + r1i + r2l | ||||
| In0–10 cm or 10–20 cm or 20–30 cm or 30–40 cm or 40–50 cmi ~ Binomial (Ini, logistic (pi)) | 1, 2, …, 124 | 0 or 1 | 1, 2, 3 | |
| pi = β0 + β1Sj + β2Gj + r1i + r2l | ||||
| In0–10 cm or 10–20 cm or 20–30 cm or 30–40 cm or 40–50 cmi ~ Binomial (Ini, logistic (pi)) | 1, 2, …, 124 | 0 or 1 | 1, 2, 3 | |
| μi = β0 + β1Sj + β2Gj + β3Phi + β4SjPhi + β5GjPhi + r1i + r2l |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Bai, Y.; Cao, Z.; Xu, J.; Xie, Y.; Zheng, H.; Jiang, J.; Mu, C.; Cheng, W.; Fang, H.; et al. Plasticity in the Morphology of Growing Bamboo: A Bayesian Analysis of Exogenous Treatment Effects on Plant Height, Internode Length, and Internode Numbers. Plants 2023, 12, 1713. https://doi.org/10.3390/plants12081713
Wu C, Bai Y, Cao Z, Xu J, Xie Y, Zheng H, Jiang J, Mu C, Cheng W, Fang H, et al. Plasticity in the Morphology of Growing Bamboo: A Bayesian Analysis of Exogenous Treatment Effects on Plant Height, Internode Length, and Internode Numbers. Plants. 2023; 12(8):1713. https://doi.org/10.3390/plants12081713
Chicago/Turabian StyleWu, Chongyang, Yucong Bai, Zhihua Cao, Junlei Xu, Yali Xie, Huifang Zheng, Jutang Jiang, Changhong Mu, Wenlong Cheng, Hui Fang, and et al. 2023. "Plasticity in the Morphology of Growing Bamboo: A Bayesian Analysis of Exogenous Treatment Effects on Plant Height, Internode Length, and Internode Numbers" Plants 12, no. 8: 1713. https://doi.org/10.3390/plants12081713