
Colletotrichum Species Associated with Anthracnose in Salix babylonica in China

Abstract
:1. Introduction
2. Results
2.1. Field Symptoms and Fungal Isolation
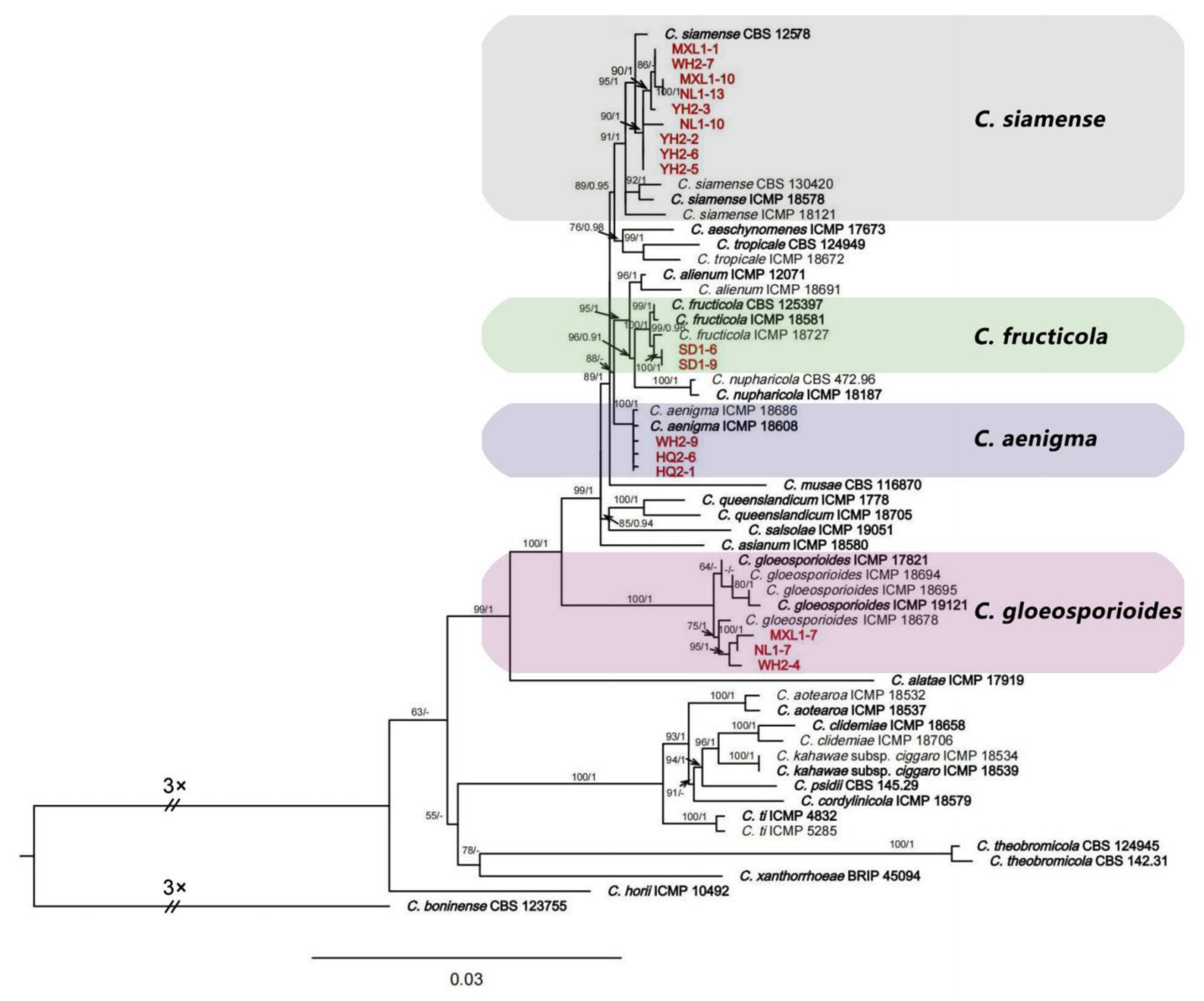
2.2. Multi-Locus Phylogenetic Analyses
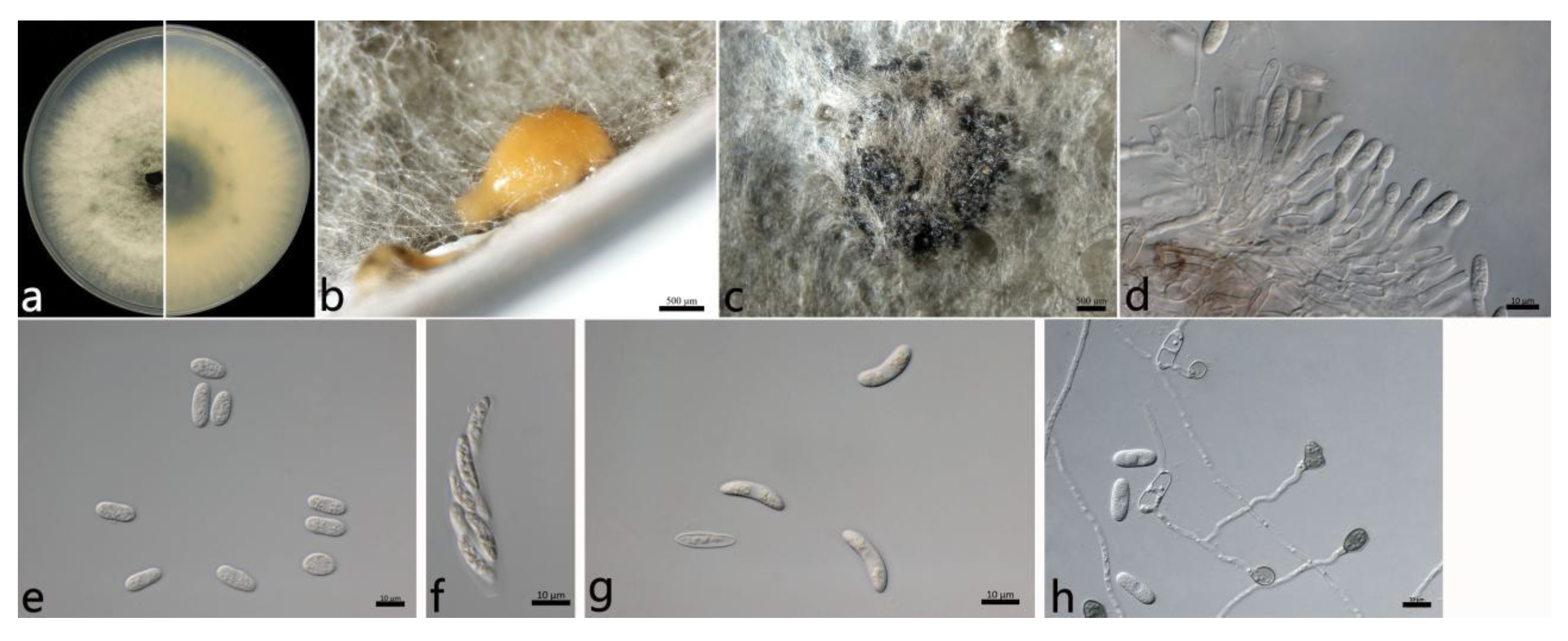
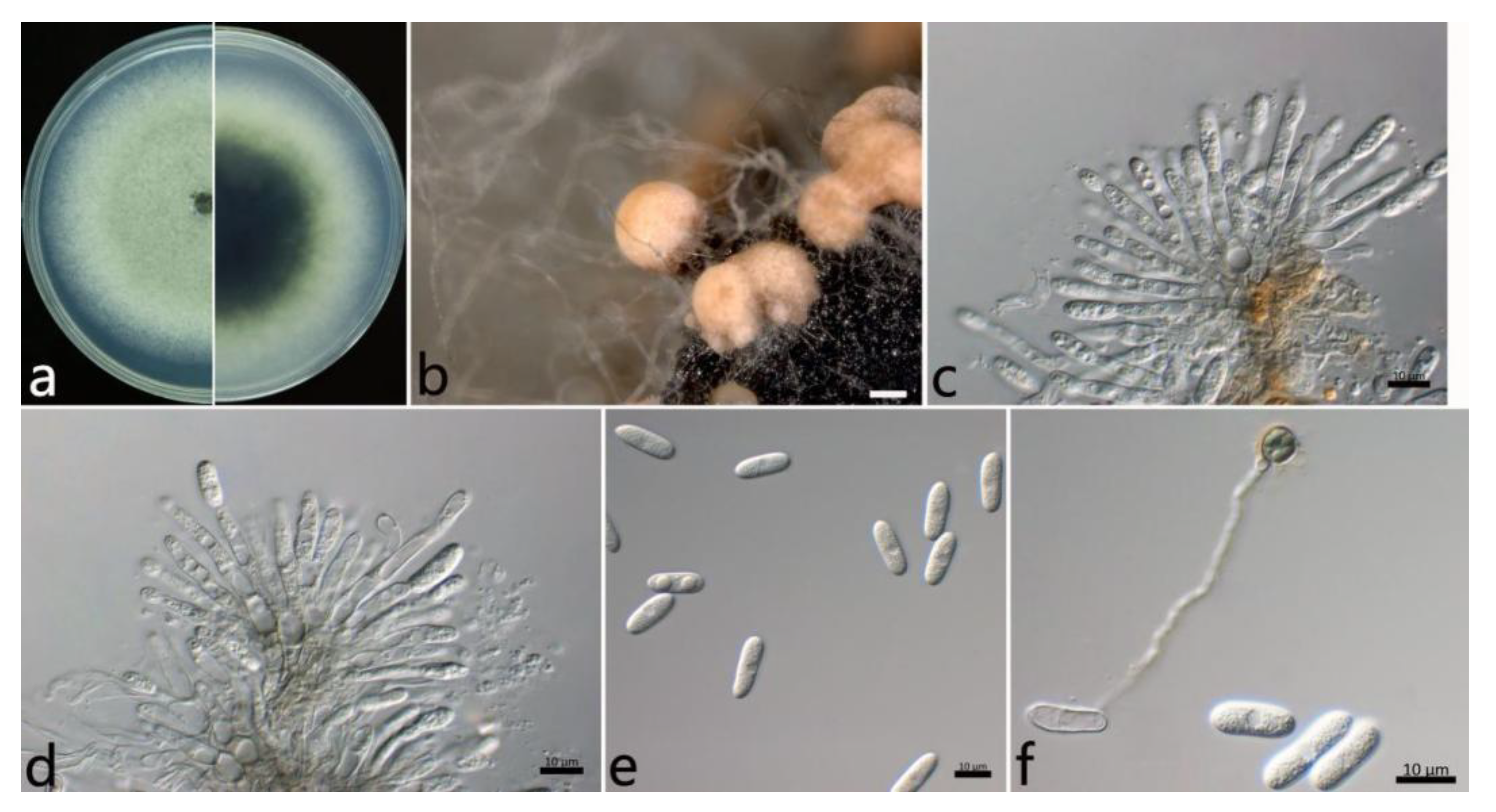
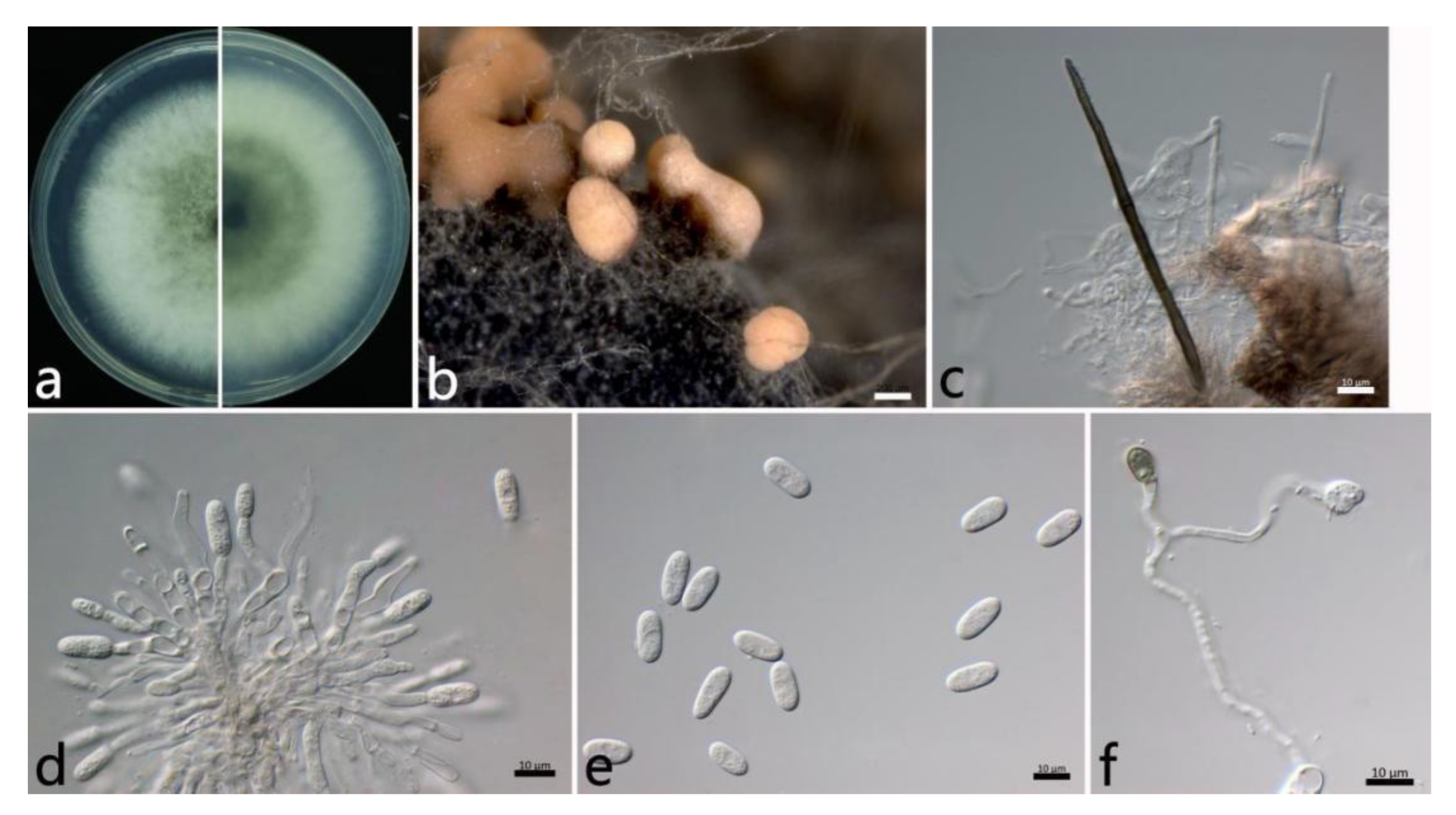
2.3. Morphological Study
2.3.1. Colletotrichum aenigma B. Weir and P.R. Johnst (Figure 3)

2.3.2. Colletotrichum fructicola Prihastuti, L. Cai and K.D. Hyde (Figure 4)

2.3.3. Colletotrichum gloeosporioides s.s. (Penz.) Penz. and Sacc (Figure 5)

2.3.4. Colletotrichum siamense Prihastuti, L. Cai and K. D. Hyde (Figure 6)

2.4. Pathogenicity Tests
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Fungi Isolation
4.2. DNA Extraction, PCR Amplification, and Sequencing
4.3. Phylogenetic Analyses
4.4. Morphological Study
4.5. Pathogenicity Tests
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum-current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Diao, Y.Z.; Zhang, C.; Liu, F.; Wang, W.Z.; Liu, L.; Cai, L.; Liu, X.L. Colletotrichum species causing anthracnose disease of chili in China. Persoonia 2017, 38, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Cannon, P.F.; Liu, F.; Barreto, R.W.; Guatimosim, E.; Crous, P.W. The Colletotrichum orbiculare species complex: Important pathogens of field crops and weeds. Fungal Divers. 2013, 61, 29–59. [Google Scholar] [CrossRef]
- Damm, U.; O’Connell, R.J.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum destructivum species complex—Hemibiotrophic pathogens of forage and field crops. Stud. Mycol. 2014, 79, 49–84. [Google Scholar] [CrossRef]
- Crouch, J.A. Colletotrichum caudatum s.l. is a species complex. IMA Fungus 2014, 5, 17–30. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.R.; Jones, E.B.G.; et al. Colletotrichum-names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Cai, L.; Hyde, K.; Taylor, P.; Weir, B.; Waller, J.; Abang, M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 2009, 39, 183–204. [Google Scholar]
- Liu, L.P.; Gao, J.; Li, Y. Research progress of plant Colletotrichum spp. J. Fungal Res. 2020, 18, 266–281. [Google Scholar]
- Sharma, G.; Shenoy, B.D. Colletotrichum systematics: Past, present and prospects. Mycosphere 2016, 7, 1093–1102. [Google Scholar] [CrossRef]
- Werbel, W.A.; Baroncelli, R.; Shoham, S.; Zhang, S.X. Angioinvasive, cutaneous infection due to Colletotrichum siamense in a stem cell transplant recipient: Report and review of prior cases. Transpl. Infect. Dis. 2019, 21, e13153. [Google Scholar] [CrossRef]
- Marcelino, J.A.; Gouli, S.; Parker, B.L.; Skinner, M.; Giordano, R. Entomopathogenic activity of a variety of the fungus, Colletotrichum acutatum, recovered from the elongate hemlock scale, Fiorinia externa. J. Insect Sci. 2009, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.J.; Edwards, S.; Inocencio, H.A.; Machado, F.J.; Nuckles, E.M.; Farman, M.; Gauthier, N.A.; Vaillancourt, L.J. Diversity and cross-infection potential of Colletotrichum causing fruit rots in mixed-fruit orchards in Kentucky. Plant Dis. 2021, 105, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Manova, V.; Stoyanova, Z.; Rodeva, R.; Boycheva, I.; Korpelainen, H.; Vesterinen, E.; Wirta, H.; Bonchev, G. Morphological, pathological, and genetic diversity of Colletotrichum species pathogens on solanaceous vegetable crops in Bulgaria. J. Fungi 2022, 8, 1123. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Cai, L.; McKenzie, E.H.C.; Yang, Y.L.; Zhang, J.Z.; Prihastuti, H. Colletotrichum: A catalogue of confusion. Fungal Divers. 2009, 39, 1–17. [Google Scholar]
- Seifert, K.A. Progress towards DNA barcoding of fungi. Mol. Ecol. Resour. 2010, 9 (Suppl. 1), 83–89. [Google Scholar] [CrossRef]
- Mills, P.R.; Sreenivasaprasad, S.; Brown, A.E. Detection and differentiation of Colletotrichum gloeosporioides isolates using PCR. FEMS Microbiol. Lett. 1992, 98, 137–143. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Crouch, J.A.; Clarke, B.B.; Hillman, B.I. What is the value of ITS sequence data in Colletotrichum systematics and species diagnosis? A case study using the falcate-spored graminicolous Colletotrichum group. Mycologia 2009, 101, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Stielow, J.B.; Levesque, C.A.; Seifert, K.A.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Damm, U.; Cai, L.; Crous, P.W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Divers. 2013, 61, 89–105. [Google Scholar] [CrossRef]
- Liu, F.; Cai, L.; Crous, P.W.; Damm, U. The Colletotrichum gigasporum species complex. Persoonia 2014, 33, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Groenewald, J.Z.; Polizzi, G.; Crous, P.W. High species diversity in Colletotrichum associated with citrus diseases in Europe. Persoonia 2017, 39, 32–50. [Google Scholar] [CrossRef]
- Huang, R.; Sun, W.X.; Wang, L.R.; Li, Q.L.; Huang, S.P.; Tang, L.H.; Guo, T.X.; Mo, J.Y.; Hsiang, T. Identification and characterization of Colletotrichum species associated with anthracnose disease of banana. Plant Pathol. 2021, 70, 1827–1837. [Google Scholar] [CrossRef]
- Argus, G.W. Infrageneric classification of Salix (Salicaceae) in the New World. Syst. Bot. Monogr. 1997, 52, 1–121. [Google Scholar] [CrossRef]
- Li, H.; Zhang, G.C.; Xie, H.C.; Zhang, S.Y. The effects of the phenol concentrations on photosynthetic parameters of Salix babylonica L. Photosynthetica 2015, 53, 430–435. [Google Scholar] [CrossRef]
- Chen, C.H.; Liu, Y.K.; Chen, G.C.; Shan, Q.H.; Zhang, J.F. Uptake kinetic characteristics of Cu2+ by Salix jiangsuensis CL J-172 and Salix babylonica Linn and the influence of organic acids. Acta Ecol. Sin. 2011, 31, 5255–5263. [Google Scholar]
- Zhang, M.Y.; Li, D.W.; Zhu, L.H. Leaf spots of Salix babylonica caused by Colletotrichum gloeosporioides s.s. and C. siamense newly reported in China. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Mirck, J.; Isebrands, J.G.; Verwijst, T.; Ledin, S. Development of short-rotation willow coppice system for environmental purposes in Sweden. Biomass Bioenergy 2005, 28, 219–228. [Google Scholar] [CrossRef]
- Yu, X.Z.; Trapp, S.; Zhou, P.H. Phytotoxicity of cyanide to weeping willow trees. Environ. Sci. Pollut. Res. 2005, 12, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Trapp, S.; Pirandello, A. Removal of cyanide by woody plants. Chemosphere 2004, 54, 325–333. [Google Scholar] [CrossRef]
- Yulla, A.K.; Martin, F. Willows beyond wetland: Use of Salix L. species for environmental projects. Water Air Soil Pollut. 2005, 162, 183–204. [Google Scholar]
- Perttu, K.L.; Kowalik, P.J. Salix vegetation filters for purification of water and soils. Biomass Bioenergy 1997, 12, 9–19. [Google Scholar] [CrossRef]
- Pulford, I.D.; Riddell-Black, D.; Stewart, C. Heavy metal uptake by willow clones from sewage sludge-treated soil: The potential for phytoremediation. Int. J. Phytoremed. 2002, 4, 59–72. [Google Scholar] [CrossRef]
- Singh, H.; Raturi, R.; Badoni, P.P. Isolation of secondary metabolites from the roots of Salix babylonica. In Proceedings of the International Conference on Materials, Alloys and Experimental Mechanics (ICMAEM), Secunderabad, India, 3–4 July 2017. [Google Scholar]
- Ganai, A.M.; Ahmad, H.A.; Matto, F.A. Nutritive value of Salix leaves for sheep. Indian Vet. J. 2006, 83, 895–896. [Google Scholar]
- Mu, D.; Liu, Z.K.; Tao, Y.; Zhou, L.Z.; Wang, J.Q.; Xu, W.W.; Luo, G.Q. Analysis of volatile compounds of Salix babylonica by GC-MS. J. Chin. Med. Mater. 2014, 37, 1001–1005. [Google Scholar]
- Tzavella-Klonari, K.; Aggelaki, M.D.; Karadimos, D.A. First report of anthracnose on weeping willow in Greece. Plant Dis. 1997, 81, 960. [Google Scholar] [CrossRef] [PubMed]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Yang, Y.L.; Liu, Z.Y.; Cai, L.; Hyde, K.D.; Yu, Z.N.; McKenzie, E.H.C. Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers. 2009, 39, 123–146. [Google Scholar]
- Wikee, S.; Cai, L.; Pairin, N.; McKenzie, E.H.C.; Su, Y.Y.; Chukeatirote, E.; Thi, H.N.; Bahkali, A.H.; Moslem, M.A.; Abdelsalam, K.; et al. Colletotrichum species from jasmine (Jasminum sambac). Fungal Divers. 2011, 46, 171–182. [Google Scholar] [CrossRef]
- Thaochan, N.; Pornsuriya, C.; Chairin, T.; Chomnunti, P.; Sunpapao, A. Morphological and molecular characterization of Calonectria foliicola associated with leaf blight on rubber Tree (Hevea brasiliensis) in Thailand. J. Fungi 2022, 8, 986. [Google Scholar] [CrossRef]
- Silva, D.N.; Talhinhas, P.; Várzea, V.; Cai, L.; Paulo, O.S.; Batista, D. Application of the Apn2/MAT locus to improve the systematics of the Colletotrichum gloeosporioides complex: An example from coffee (Coffea spp.) hosts. Mycologia 2012, 104, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Sunpapao, A.; Suwannarach, N.; Kumla, J.; Dumhai, R.; Riangwong, K.; Sanguansub, S.; Wanchana, S.; Arikit, S. Morphological and molecular identification of plant pathogenic fungi associated with dirty panicle disease in coconuts (Cocos nucifera) in Thailand. J. Fungi 2022, 8, 335. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef]
- Wang, Y.C.; Xiong, F.; Lu, Q.H.; Hao, X.Y.; Zheng, M.X.; Wang, L.; Li, N.N.; Ding, C.Q.; Wang, X.C.; Yang, Y.J. Diversity of Pestalotiopsis-like species causing gray blight disease of tea plants (Camellia sinensis) in China, including two novel Pestalotiopsis species, and analysis of their pathogenicity. Plant Dis. 2019, 103, 2548–2558. [Google Scholar] [CrossRef]
- Poudel, B.; Zhang, S. First report of Erysiphe fallax causing powdery mildew on phasey bean (Macroptilium lathyroides) in the United States. Plant Dis. 2019, 20, 35–37. [Google Scholar] [CrossRef]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Li, D.W.; Zhu, Y.N.; Si, Y.Z.; Huang, J.H.; Zhu, L.H.; Ye, J.R.; Huang, L. Diversity and pathogenicity of Colletotrichum species causing anthracnose on Cunninghamia lanceolata. Plant Pathol. 2022, 71, 1757–1773. [Google Scholar] [CrossRef]
- Zhang, P.F.; Zhai, L.F.; Zhang, X.K.; Huang, X.Z.; Hong, N.; Xu, W.X.; Wang, G.P. Characterization of Colletotrichum fructicola, a new causal agent of leaf black spot disease of sandy pear (Pyrus pyrifolia). Eur. J. Plant Pathol. 2015, 143, 651–662. [Google Scholar] [CrossRef]
- Li, S.N.; Zhang, W.M. Post-harvest anthracnose of carambola (Averrhoa carambola) caused by Colletotrichum fructicola in China. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Han, S.; Xu, X.; Jiang, Y.R.; Yuan, H.; Li, S.J.; Liu, Y.; Lin, T.; Qiao, T.; Yang, C.; Li, S.; et al. Colletotrichum fructicola causal agent of shot-hole symptoms on leaves of Prunus sibirica in China. Plant Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Usman, H.M.; Tan, Q.; Karim, M.M.; Adnan, M.; Yin, W.X.; Zhu, F.X.; Luo, C.X. Sensitivity of Colletotrichum fructicola and Colletotrichum siamense of peach in China to multiple classes of fungicides and characterization of pyraclostrobin-resistant isolates. Plant Dis. 2022, 105, 3459–3465. [Google Scholar] [CrossRef]
- Lin, W.L.; Duan, C.H.; Wang, C.L. Identification and virulence of Colletotrichum species causing anthracnose on mango. Plant Pathol. 2023, 72, 623–635. [Google Scholar] [CrossRef]
- Swain, S.V.; Koike, S.T.; Michailides, T.J.; Feng, C.; Correll, J.C. First report of twig canker on willow caused by Colletotrichum acutatum in California. Plant Dis. 2012, 96, 1822–1823. [Google Scholar] [CrossRef]
- Si, Y.Z.; Sun, J.W.; Li, D.W.; Huang, L.; Ju, Y.; Zhu, L.H. Leaf spot of Sapindus mukorossi caused by Diaporthe biconispora in China. Australas. Plant Pathol. 2021, 50, 193–202. [Google Scholar] [CrossRef]
- Sun, J.W.; Si, Y.Z.; Li, D.W.; Jin, G.Q.; Zhu, L.H. First report of leaf blotch of Aesculus chinensis caused by Colletotrichum gloeosporioides and Colletotrichum fructicola in China. Plant Dis. 2020, 104, 3065–3066. [Google Scholar] [CrossRef]
- Huang, L.; Li, Q.C.; Zhang, Y.; Li, D.W.; Ye, J.R. Colletotrichum gloeosporioides sensu stricto is a pathogen of leaf anthracnose on Evergreen Spindle tree (Euonymus japonicus). Plant Dis. 2016, 100, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Li, M.F.; He, J.; Ding, L.; Kang, J.; Zhang, Q.; Zheng, Q. Single spore strains without producing fruit body isolated from Cordyceps militeris and their RAPD analysis. Southwest China J. Agric. Sci. 2007, 20, 547–550. [Google Scholar]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum gloeosporioides isolates from avocado and almond fruits with molecular and pathogenicity tests. Appl. Environ. Microbiol. 1996, 62, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Luo, C.X.; Wu, H.J.; Peng, B.; Kang, B.S.; Liu, L.M.; Zhang, M.; Gu, Q.S. Colletotrichum species associated with anthracnose disease of watermelon (Citrullus lanatus) in China. J. Fungi 2022, 8, 790. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Host Tissue | Time | Latitude and Longitude | Number of Isolates |
|---|---|---|---|---|
| Suzhou, Jiangsu | leaf | 2021.6.23 | 31°20′34″ N, 120°35′18″ E | 6 |
| Suzhou, Jiangsu | leaf | 2021.6.23 | 31°18′30″ N, 120°34′41″ E | 6 |
| Zibo, Shandong | leaf | 2021.9.11 | 36°37′58″ N, 117°53′43″ E | 10 |
| Wuhan, Hubei | leaf | 2021.10.13 | 30°43′10″ N, 114°31′59″ E | 10 |
| Nanjing, Jiangsu | leaf | 2021.10.20 | 32°5′10″ N, 118°49′13″ E | 13 |
| Nanjing, Jiangsu | leaf | 2021.10.28 | 32°3′2″ N, 118°50′26″ E | 10 |
| No. | Species | Isolate | Lesion Length (mm) | No. | Species | Isolate | Lesion Length (mm) |
|---|---|---|---|---|---|---|---|
| 1 | C. aenigma | HQ2-1 | 1.4 ± 0.1 f | 10 | C. siamense | YH2-3 | 8.8 ± 0.1 a |
| 2 | C. aenigma | HQ2-6 | 1.6 ± 0.1 f | 11 | C. siamense | YH2-5 | 5.7 ± 0.1 c |
| 3 | C. aenigma | WH2-9 | 3.3 ± 0.2 e | 12 | C. siamense | YH2-6 | 5.6 ± 0.2 c |
| 4 | C. fructicola | SD1-6 | 3.2 ± 0.2 e | 13 | C. siamense | WH2-7 | 4.1 ± 0.3 d |
| 5 | C. fructicola | SD1-9 | 3.3 ± 0.1 e | 14 | C. siamense | NL1-10 | 1.4 ± 0.2 f |
| 6 | C. gloeosporioides | WH2-4 | 3.4 ± 0.2 e | 15 | C. siamense | NL1-13 | 8.3 ± 0.1 b |
| 7 | C. gloeosporioides | NL1-7 | 1.5 ± 0.3 f | 16 | C. siamense | MXL1-1 | 8.3 ± 0.2 b |
| 8 | C. gloeosporioides | MXL1-7 | 5.5 ± 0.2 c | 17 | C. siamense | MXL1-10 | 8.2 ± 0.2 b |
| 9 | C. siamense | YH2-2 | 5.6 ± 0.2 c |
| Region | Primer | Direction | Sequence (5′–3′) | Tm (°C) |
|---|---|---|---|---|
| ITS | ITS1 | Forward | TCCGTAGGTGAACCTGCGG | 55 |
| ITS4 | Reverse | TCCTCCGCTTATTGATATGC | ||
| ACT | ACT-512F | Forward | ATGTGCAAGGCCGGTTTCGC | 58 |
| ACT-783R | Reverse | TACGAGTCCTTCTGGCCCAT | ||
| CHS-1 | CHS-79F | Forward | TGGGGCAAGGATGCCTGGAAGAAG | 58 |
| CHS-354R | Reverse | TGGAAGAACCATCTGTGAGAGTTG | ||
| TUB2 | T1 | Forward | AACATGCGTGAGATTGTAAGT | 55 |
| Bt2b | Reverse | ACCCTCAGTGTAGTGACCCTTGGC | ||
| CAL | CL1C | Forward | GAATTCAAGGAGGCCTTCTC | 55 |
| CL2C | Reverse | CTTCTGCATCATGAGCTGGAC | ||
| GAPDH | GDF1 | Forward | GCCGTCAACGACCCCTTCATTGA | 58 |
| Species | Culture * | Host | Country | GenBank Accession Number | |||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | GAPDH | CAL | ACT | CHS-1 | TUB2 | ||||
| C. aenigma | ICMP 18608 * | Persea americana | Israel | JX010244 | JX010044 | JX009683 | JX009443 | JX009774 | JX010389 |
| ICMP 18686 | Pyrus pyrifolia | Japan | JX010243 | JX009913 | JX009684 | JX009519 | JX009789 | JX010390 | |
| HQ2-1 | S. babylonica | China | OQ253546 | OQ428578 | OQ428572 | OQ428569 | OQ428575 | OQ428581 | |
| HQ2-6 | S. babylonica | China | OQ243538 | OQ428579 | OQ428573 | OQ428570 | OQ428576 | OQ428582 | |
| WH2-9 | S. babylonica | China | OQ253555 | OQ428580 | OQ428574 | OQ428571 | OQ428577 | OQ428583 | |
| C. aeschynomenes | ICMP 17673 * | Aeschynomene virginica | USA | JX010176 | JX009930 | JX009721 | JX009483 | JX009799 | JX010392 |
| C. alatae | ICMP 17919 * | Dioscorea alata | India | JX010190 | JX009990 | JX009738 | JX009471 | JX009837 | JX010383 |
| C. alienum | ICMP 18691 | Persea americana | Australia | JX010217 | JX010018 | JX009664 | JX009580 | JX009754 | JX010385 |
| ICMP 12071 * | Malus domestica | New Zealand | JX010251 | JX010028 | JX009654 | JX009572 | JX009882 | JX010411 | |
| C. aotearoa | ICMP 18532 | Vitex lucens | New Zealand | JX010220 | JX009906 | JX009614 | JX009544 | JX009764 | JX010421 |
| ICMP 18537 * | Coprosma sp. | New Zealand | JX010205 | JX010005 | JX009611 | JX009564 | JX009853 | JX010420 | |
| C. asianum | ICMP 18580 * | Coffea arabica | Thailand | FJ972612 | JX010053 | FJ917506 | JX009584 | JX009867 | JX010406 |
| C. boninense | CBS 123755 * | Crinum asiaticum var. sinicum | Japan | JX010292 | JX009905 | -- | JX009583 | JX009827 | -- |
| C. clidemiae | ICMP 18706 | Vitis sp. | USA | JX010274 | JX009909 | JX009639 | JX009476 | JX009777 | JX010439 |
| ICMP 18658 * | Clidemia hirta | USA, Hawaii | JX010265 | JX009989 | JX009645 | JX009537 | JX009877 | JX010438 | |
| C. cordylinicola | ICMP 18579 * | Cordyline fruticosa | Thailand | JX010226 | JX009975 | HM470238 | HM470235 | JX009864 | JX010440 |
| C. fructicola | ICMP 18581 * | Coffea arabica | Thailand | JX010165 | JX010033 | FJ917508 | FJ907426 | JX009866 | JX010405 |
| ICMP 18727 | Fragaria × ananassa | USA | JX010179 | JX010035 | JX009682 | JX009565 | JX009812 | JX010394 | |
| SD1-6 | S. babylonica | China | OQ253556 | OQ428565 | OQ428561 | OQ428559 | OQ428563 | OQ428567 | |
| SD1-9 | S. babylonica | China | OQ253557 | OQ428566 | OQ428562 | OQ428560 | OQ428564 | OQ428568 | |
| C. fructicola (syn. C. ignotum) | CBS 125397 (*) | Tetragastris panamensis | Panama | JX010173 | JX010032 | JX009674 | JX009581 | JX009874 | JX010409 |
| C. gloeosporioides | ICMP 17821 * | Citrus sinensis | Italy | JX010152 | JX010056 | JX009731 | JX009531 | JX009818 | JX010445 |
| ICMP 18694 | Mangifera indica | South Africa | JX010155 | JX009980 | JX009729 | JX009481 | JX009796 | -- | |
| ICMP 18678 | Pueraria lobata | USA | JX010150 | JX010013 | JX009733 | JX009502 | JX009790 | -- | |
| ICMP 18695 | Citrus sp. | USA | JX010153 | JX009979 | JX009735 | JX009494 | JX009779 | -- | |
| WH2-4 | S. babylonica | China | OQ243548 | OQ428555 | OQ428551 | OQ428549 | OQ428553 | OQ428557 | |
| NL1-7 | S. babylonica | China | ON870951 | ON858480 | ON858478 | ON858477 | ON858479 | ON858481 | |
| MXL1-7 | S. babylonica | China | OQ253571 | OQ428556 | OQ428552 | OQ428550 | OQ428554 | OQ428558 | |
| C. gloeosporioides (syn. Gloeosporium pedemontanum) | ICMP 19121 (*) | Citrus limon | Italy | JX010148 | JX010054 | JX009745 | JX009558 | JX009903 | -- |
| C. horii | ICMP 10492 * | Diospyros kaki | Japan | GQ329690 | GQ329681 | JX009604 | JX009438 | JX009752 | JX010450 |
| C. kahawae subsp. ciggaro | ICMP 18539 * | Olea europaea | Australia | JX010230 | JX009966 | JX009635 | JX009523 | JX009800 | JX010434 |
| ICMP 18534 | Kunzea ericoides | New Zealand | JX010227 | JX009904 | JX009634 | JX009473 | JX009765 | JX010427 | |
| C. musae | CBS 116870 * | Musa sp. | USA | JX010146 | JX010050 | JX009742 | JX009433 | JX009896 | HQ596280 |
| C. nupharicola | ICMP 18187 * | Nuphar lutea subsp. polysepala | USA | JX010187 | JX009972 | JX009663 | JX009437 | JX009835 | JX010398 |
| CBS 472.96 | Nymphaea ordorata | USA | JX010188 | JX010031 | JX009662 | JX009582 | JX009836 | JX010399 | |
| C. psidii | CBS 145.29 * | Psidium sp. | Italy | JX010219 | JX009967 | JX009743 | JX009515 | JX009901 | JX010443 |
| C. queenslandicum | ICMP 1778 * | Carica papaya | Australia | JX010276 | JX009934 | JX009691 | JX009447 | JX009899 | JX010414 |
| ICMP 18705 | Coffea sp. | Fiji | JX010185 | JX010036 | JX009694 | JX009490 | JX009890 | JX010412 | |
| C. salsolae | ICMP 19051 * | Salsola tragus | Hungary | JX010242 | JX009916 | JX009696 | JX009562 | JX009863 | JX010403 |
| C. siamense | ICMP 18121 | Dioscorea rotundata | Nigeria | JX010245 | JX009942 | JX009715 | JX009460 | JX009845 | JX010402 |
| ICMP 18578 * | Coffea arabica | Thailand | JX010171 | JX009924 | FJ917505 | FJ907423 | JX009865 | JX010404 | |
| YH2-2 | S. babylonica | China | OQ243534 | OQ428605 | OQ428591 | OQ428584 | OQ428598 | OQ428612 | |
| YH2-3 | S. babylonica | China | OQ253535 | OQ428606 | OQ428592 | OQ428585 | OQ428599 | OQ428613 | |
| YH2-5 | S. babylonica | China | OQ253537 | OQ428607 | OQ428593 | OQ428586 | OQ428600 | OQ428614 | |
| YH2-6 | S. babylonica | China | OQ253536 | OQ428608 | OQ428594 | OQ428587 | OQ428601 | OQ428615 | |
| WH2-7 | S. babylonica | China | OQ253552 | OQ428609 | OQ428595 | OQ428588 | OQ428602 | OQ428616 | |
| NL1-10 | S. babylonica | China | ON908707 | ON858485 | ON858483 | ON858482 | ON858484 | ON858486 | |
| NL1-13 | S. babylonica | China | ON870949 | ON858490 | ON858488 | ON858487 | ON858489 | ON858491 | |
| MXL1-1 | S. babylonica | China | OQ253561 | OQ428610 | OQ428596 | OQ428589 | OQ428603 | OQ428617 | |
| MXL1-10 | S. babylonica | China | OQ253562 | OQ428611 | OQ428597 | OQ428590 | OQ428604 | OQ428618 | |
| C. siamense (syn. C. hymenocallidis) | CBS 125378 (*) | Hymenocallis americana | China | JX010278 | JX010019 | JX009709 | GQ856775 | GQ856730 | JX010410 |
| C. siamense (syn. C. jasmini-sambac) | CBS 130420 (*) | Jasminum sambac | Vietnam | HM131511 | HM131497 | JX009713 | HM131507 | JX009895 | JX010415 |
| C. theobromicola | CBS 124945 *, ICMP 18649 | Theobroma cacao | Panama | JX010294 | JX010006 | JX009591 | JX009444 | JX009869 | JX010447 |
| C. theobromicola (syn. C. fragariae) | CBS 142.31 (*) | Fragaria × ananassa | USA | JX010286 | JX010024 | JX009592 | JX009516 | JX009830 | JX010373 |
| C. ti | ICMP 5285 | Cordyline australis | New Zealand | JX010267 | JX009910 | JX009650 | JX009553 | JX009897 | JX010441 |
| ICMP 4832 * | Cordyline sp. | New Zealand | JX010269 | JX009952 | JX009649 | JX009520 | JX009898 | JX010442 | |
| C. tropicale | ICMP 18672 | Litchi chinensis | Japan | JX010275 | JX010020 | JX009722 | JX009480 | JX009826 | JX010396 |
| CBS 124949 * | Theobroma cacao | Panama | JX010264 | JX010007 | JX009719 | JX009489 | JX009870 | JX010407 | |
| C. xanthorrhoeae | BRIP 45094 * | Xanthorrhoea preissii | Australia | JX010261 | JX009927 | JX009653 | JX009478 | JX009823 | JX010448 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Li, D.; Si, Y.; Ju, Y.; Zhu, L. Colletotrichum Species Associated with Anthracnose in Salix babylonica in China. Plants 2023, 12, 1679. https://doi.org/10.3390/plants12081679
Zhang M, Li D, Si Y, Ju Y, Zhu L. Colletotrichum Species Associated with Anthracnose in Salix babylonica in China. Plants. 2023; 12(8):1679. https://doi.org/10.3390/plants12081679
Chicago/Turabian StyleZhang, Mengyu, Dewei Li, Yuanzhi Si, Yue Ju, and Lihua Zhu. 2023. "Colletotrichum Species Associated with Anthracnose in Salix babylonica in China" Plants 12, no. 8: 1679. https://doi.org/10.3390/plants12081679