
Allelopathic Potential of Marsdenia tenacissima (Roxb.) Moon against Four Test Plants and the Biological Activity of Its Allelopathic Novel Compound, 8-Dehydroxy-11β-O-Acetyl-12β-O-Tigloyl-17β-Marsdenin

Abstract
:1. Introduction
2. Results
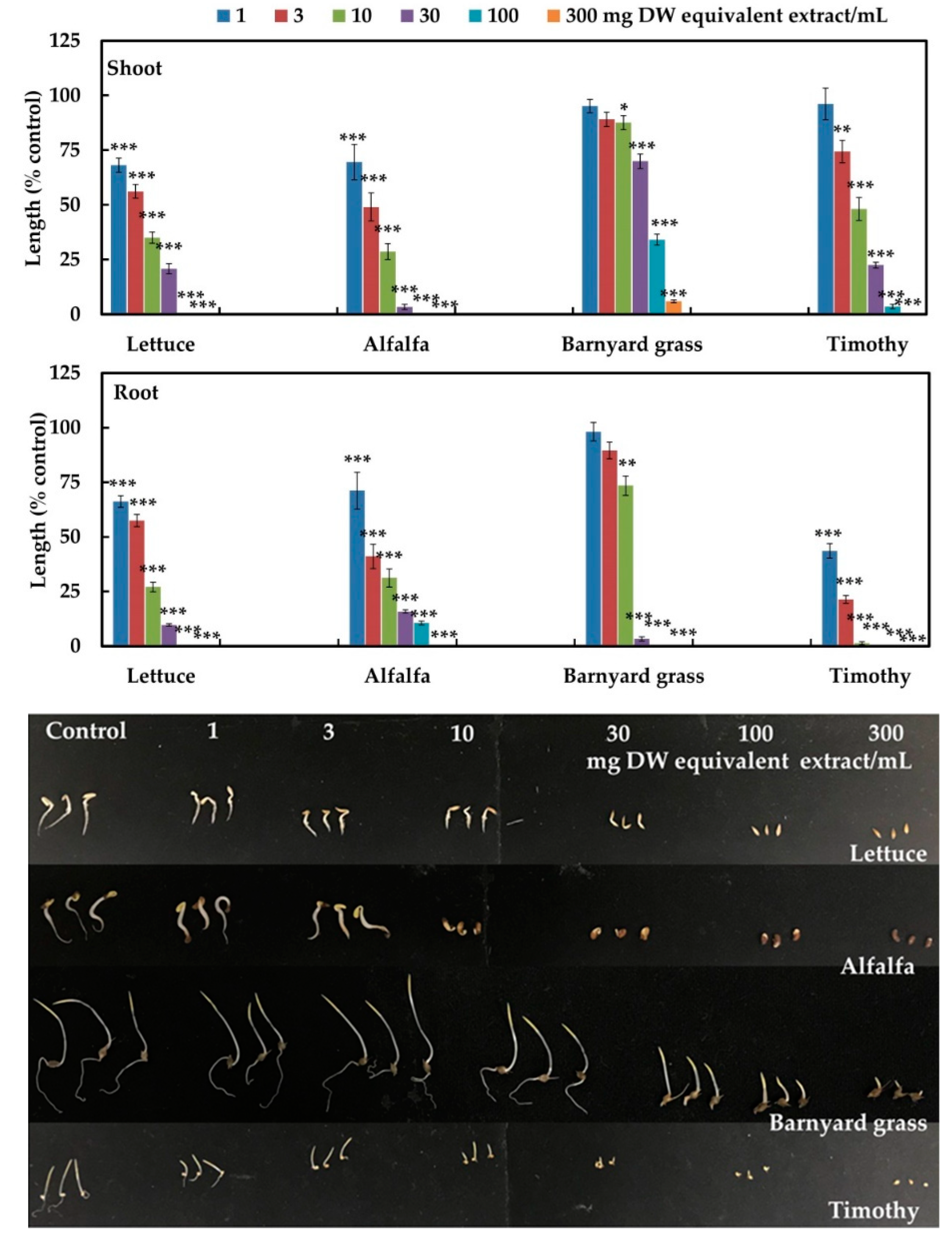
2.1. Allelopathic Activity of the Marsdenia tenacissima
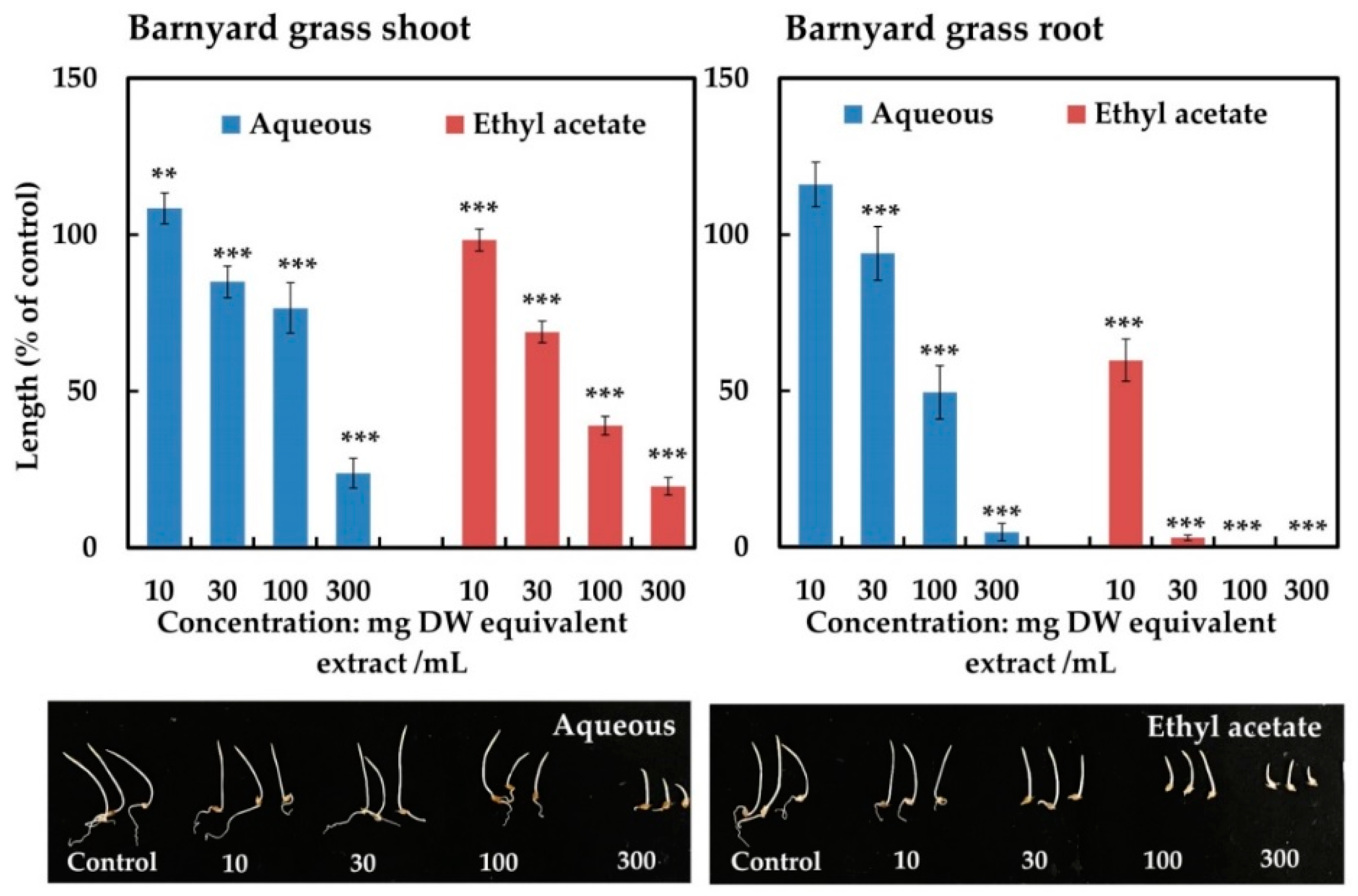
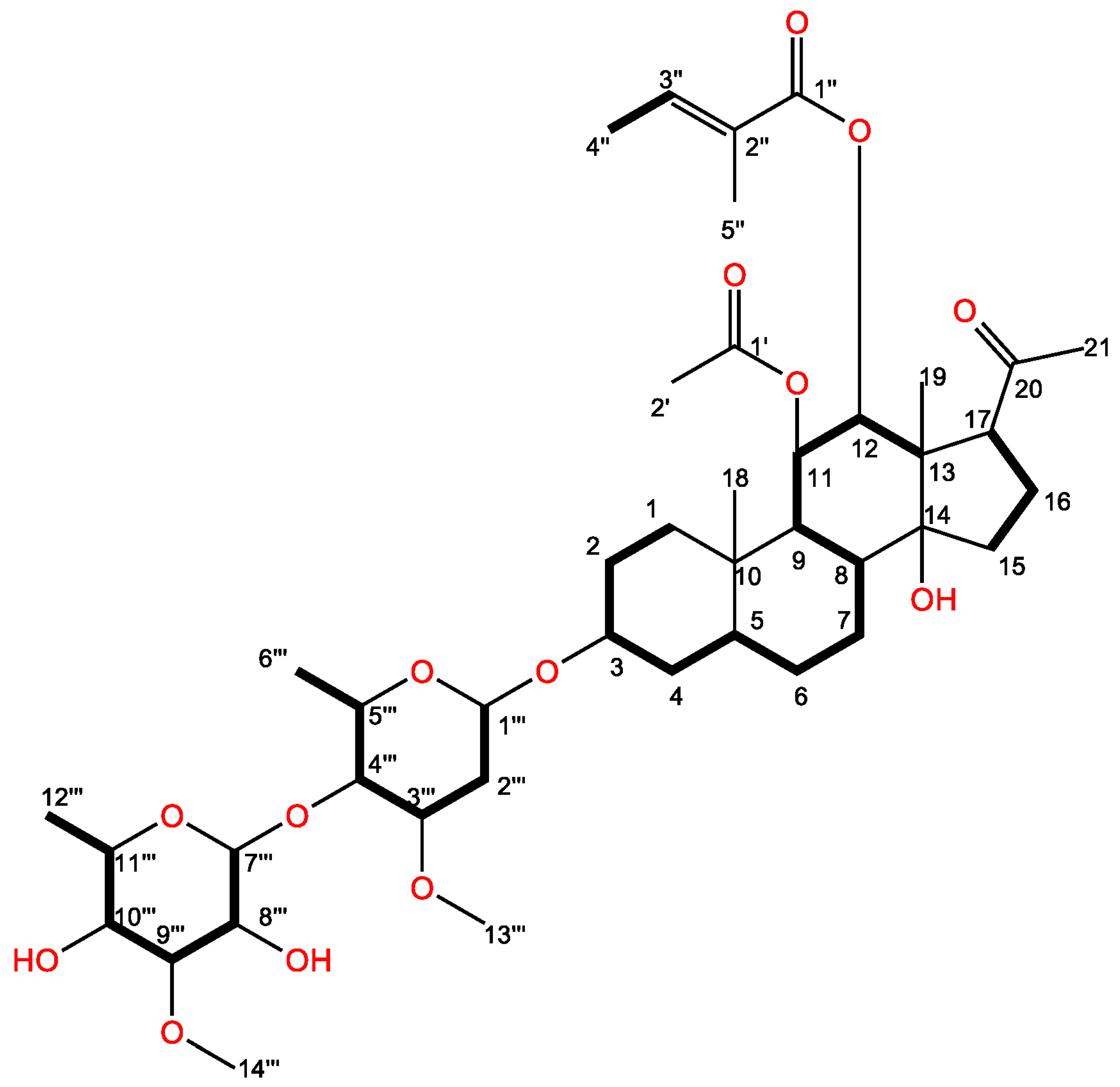
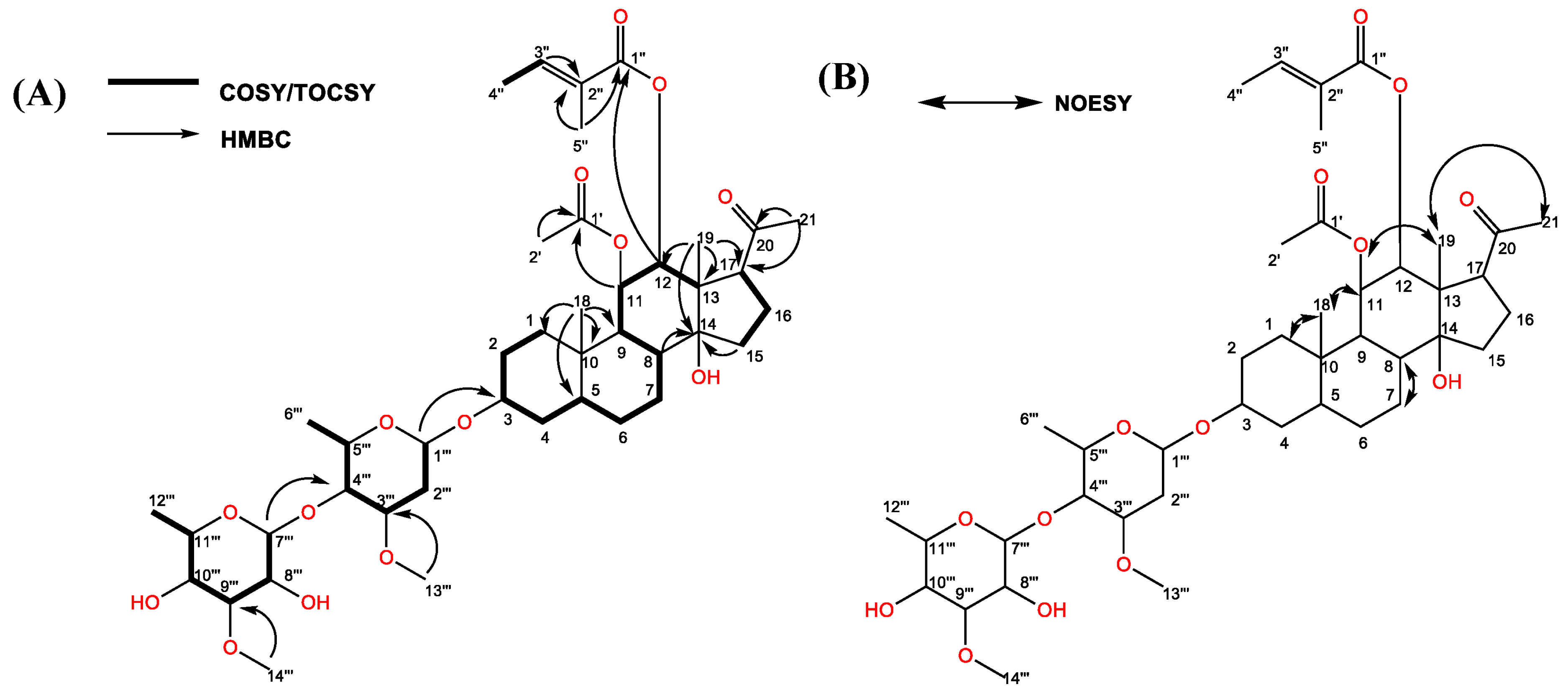
2.2. Isolation and Identification of the Active Substance
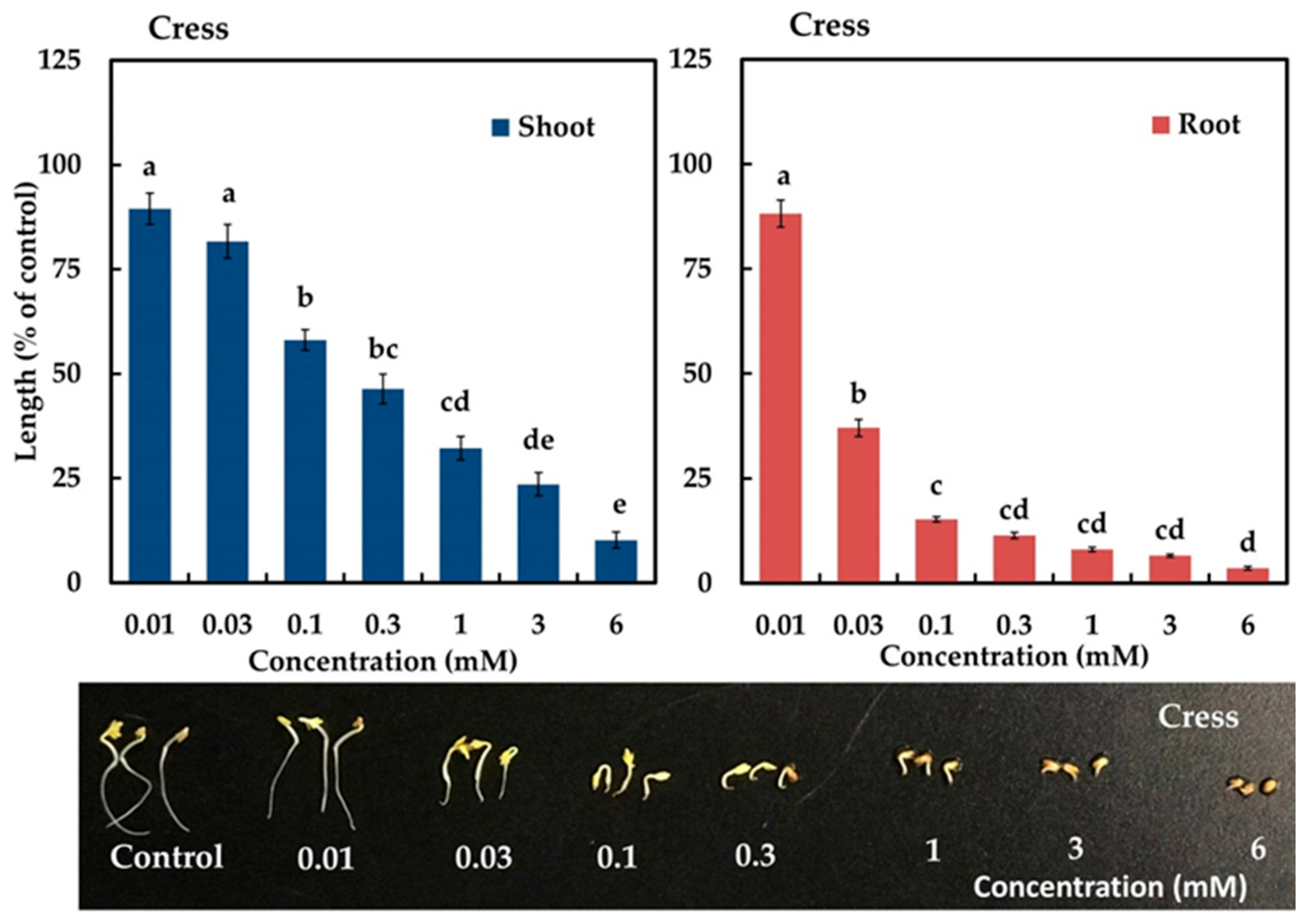
2.3. Biological Activity of the Active Compound
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Growth Bioassay
4.3. Purification of the Active Substance
4.4. Biological Activity of the Characterized Compound
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasan, M.; Ahmad-Hamdani, M.; Rosli, A.; Hamdan, H. Bioherbicides: An Eco-Friendly Tool for Sustainable Weed Management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef]
- Monteiro, A.L.; Souza, M.D.F.; Lins, H.A.; Teófilo, T.M.D.S.; Júnior, A.P.B.; Silva, D.V.; Mendonça, V. A new alternative to determine weed control in agricultural systems based on artificial neural networks (ANNs). Field Crop. Res. 2021, 263, 108075. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Qasem, J.R.; Foy, C.L. Weed allelopathy, its ecological impacts and future prospects: A review. J. Crop Prod. 2001, 4, 43–119. [Google Scholar] [CrossRef]
- Christensen, S.; Søgaard, H.T.; Kudsk, P.; Nørremark, M.; Lund, I.; Nadimi, E.; Jørgensen, R.N. Site-specific weed control technologies. Weed Res. 2009, 49, 233–241. [Google Scholar] [CrossRef]
- Pavlović, D.; Vrbničanin, S.; Andelković, A.; Božić, D.; Rajković, M.; Malidža, G. Non-chemical weed control for plant health and environment: Ecological integrated weed management (EIWM). Agronomy 2022, 12, 1091. [Google Scholar] [CrossRef]
- Xuan, T.D.; Tsuzuki, E. Allelopathic plants: Buckwheat. Allelopath. J. 2004, 13, 137–148. [Google Scholar]
- Farooq, N.; Abbas, T.; Tanveer, A.; Jabran, K. Allelopathy for Weed Management. In Co-Evolution of Secondary Metabolites. Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; ISBN 978-3-319-96396-9. [Google Scholar] [CrossRef]
- Putnam, A.R. Allelochemicals from Plants as Herbicides. Weed Technol. 1988, 2, 510–518. [Google Scholar] [CrossRef]
- Weston, L.A. Utilization of Allelopathy for Weed Management in Agroecosystems. Agron. J. 1996, 88, 860–866. [Google Scholar] [CrossRef]
- Einhellig, F.A. Interactions Involving Allelopathy in Cropping Systems. Agron. J. 1996, 88, 886–893. [Google Scholar] [CrossRef]
- Seigler, D.S. Chemistry and Mechanisms of Allelopathic Interactions. Agron. J. 1996, 88, 876–885. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources of herbicides: Current status and future trends. Weed Res. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Hussain, M.I.; Danish, S.; Sánchez-Moreiras, A.M.; Vicente, Ó.; Jabran, K.; Chaudhry, U.K.; Branca, F.; Reigosa, M.J. Unraveling Sorghum Allelopathy in Agriculture: Concepts and Implications. Plants 2021, 10, 1795. [Google Scholar] [CrossRef] [PubMed]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: A review. Agron. Sustain. Dev. 2011, 32, 273–303. [Google Scholar] [CrossRef] [Green Version]
- Scognamiglio, M.; D’abrosca, B.; Esposito, A.; Pacifico, S.; Monaco, P.; Fiorentino, A. Plant growth inhibitors: Allelopathic role or phytotoxic effects? Focus on Mediterranean biomes. Phytochem. Rev. 2013, 12, 803–830. [Google Scholar] [CrossRef]
- Scognamiglio, M.; Esposito, A.; D’abrosca, B.; Pacifico, S.; Fiumano, V.; Tsafantakis, N.; Monaco, P.; Fiorentino, A. Isolation, distribution and allelopathic effect of caffeic acid derivatives from Bellis perennis L. Biochem. Syst. Ecol. 2012, 43, 108–113. [Google Scholar] [CrossRef]
- Narwal, S.S. Allelopathy in weed management. In Allelopathy Update Basic and Applied Aspects; Narwal, S.S., Ed.; Science Publishers Inc.: Enfield, NH, USA, 1999; Volume 2, pp. 203–254. [Google Scholar]
- Kato-Noguchi, H.; Salam, A.; Ohno, O.; Suenaga, K. Nimbolide B and Nimbic Acid B, Phytotoxic Substances in Neem Leaves with Allelopathic Activity. Molecules 2014, 19, 6929–6940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyaw, E.H.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Assessment of the Phytotoxic Potential of Dregea volubilis (L.f.) Benth. ex Hook.f. and Identification of its Phytotoxic Substances for Weed Control. Agriculture 2022, 12, 1826. [Google Scholar] [CrossRef]
- Krumsri, R.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Assessment of Allelopathic Potential of Senna garrettiana Leaves and Identification of Potent Phytotoxic Substances. Agronomy 2022, 12, 139. [Google Scholar] [CrossRef]
- Wang, P.; Yang, J.; Zhu, Z.; Zhang, X. Marsdenia tenacissima: A review of traditional uses, phytochemistry and pharmacology. Am. J. Chin. Med. 2018, 46, 1–32. [Google Scholar] [CrossRef]
- Tiwari, V.; Singh, A.; Tiwari, A. Phytopharmacological overview on controversial drug: Murva. Tradit. Folk. Herb. Med. Recent Res. 2018, 2, 475–526. [Google Scholar]
- Kumar, G.; Banu, G.; Murugesan, A.G.; Pandian, M.R. Effect of Helicteres isora Bark Extract on Protein Metabolism and Marker Enzymes in Streptozotocin-Induced Diabetic Rats. Iran. J. Pharm. Sci. 2007, 6, 123–129. [Google Scholar] [CrossRef]
- Li, H.-T.; Kang, L.-P.; Guo, B.-L.; Zhang, Z.-L.; Guan, Y.-H.; Pang, X.; Peng, C.-Z.; Ma, B.-P.; Zhang, L.-X. Original plant identification of Dai nationality herb. China J. Chin. Mater. Medica 2014, 39, 1525–1529. [Google Scholar]
- Tripathi, M.; Shivhare, D.; Tiwari, A.; Ahirwar, P.; Pathak, S. Pharmacognostical evaluation of Marsdenia tenacissima Wight. & Arn. Root. Int. J. Rec. Biotech. 2014, 2, 18–23. [Google Scholar]
- Mondal, M.; Saha, S.; Hossain, M.; Al Foyjul, I.; Sarkar, C.; Hossain, S.; Khalipha, A.B.R.; Kundu, S.K. Phytochemical profiling and evaluation of bioactivities of methanolic and ethyl acetate extracts of Marsdenia tenacissima leaves. J. Herbs Spices Med. Plants 2020, 26, 405–422. [Google Scholar] [CrossRef]
- Huang, X.-D.; Liu, T.; Wang, S. Two New Polyoxypregnane Glycosides from Marsdenia tenacissima. Helv. Chim. Acta 2009, 92, 2111–2117. [Google Scholar] [CrossRef]
- Keyoumu, S.; Ma, L.Y.; Tang, Y. Clinical observation of Xiaoaiping injection combined with chemotherapy on the treatment of advanced gastric cancer. J. Basic Clin. Oncol. 2012, 25, 397–399. [Google Scholar]
- Pang, X.; Kang, L.-P.; Fang, X.-M.; Yu, H.-S.; Han, L.-F.; Zhao, Y.; Zhang, L.-X.; Yu, L.-Y.; Ma, B.-P. C21 steroid derivatives from the Dai herbal medicine Dai-Bai-Jie, the dried roots of Marsdenia tenacissima, and their screening for anti-HIV activity. J. Nat. Med. 2017, 72, 166–180. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.F.; Wang, J.; Yang, J.; Min, Y. Chemical constituents from Marsdenia tenacissima and their anti-tumor activities. Chin. Tradit. Pat. Med. 2017, 39, 334–338. [Google Scholar]
- Cheng, G.L.; Kong, L.Y.; Zang, C. Progress in chemical and pharmacological studies on Marsdenia tenacissima. Pharm. Clin. Res. 2009, 17, 135–138. [Google Scholar]
- Moh, S.M.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathic activity of a novel compound, 5,6-dihydrogen-11α-O-acetyl-12β-O-tigloyl-17β-marsdenin, and a known steroidal glycoside from the leaves of Marsdenia tenacissima (Roxb.) Moon. Agronomy 2022, 12, 1536. [Google Scholar] [CrossRef]
- Al-Harbi, N.A. Allelopathic effect of Calotropis procera, Hyoscyamus muticus and Pulicaria undulata extracts on seed germination of Portulaca oleracea and Chenopodium murale. Pak. J. Biol. Sci. 2020, 23, 1260–1266. [Google Scholar]
- Moh, S.M.; Noguchi, H.K. Efficacy of Ochna integerrima (Lour.) Merr leaf extracts against seedling growth of six important plants. Aust. J. Crop Sci. 2022, 16, 555–561. [Google Scholar] [CrossRef]
- Bari, I.N.; Kato-Noguchi, H. Phytotoxic effects of Cerbera manghas L. leaf extracts on seedling elongation of four monocot and four dicot test species. Acta Agrobot. 2017, 70, 1–7. [Google Scholar] [CrossRef]
- Das, K.R.; Kato-Noguchi, H. Assessment of allelopathic potential of Coccinia grandis L. on eight test plant species. Res. Crop. 2018, 19, 769–774. [Google Scholar]
- Poonpaiboonpipat, T.; Krumsri, R.; Kato-Noguchi, H. Allelopathic and herbicidal effects of crude extract from Chromolaena odorata (L.) R.M.King and H.Rob. on Echinochloa crus-galli and Amaranthus viridis. Plants 2021, 10, 1609. [Google Scholar] [CrossRef] [PubMed]
- Hossen, K.; Asato, Y.; Teruya, T.; Kato-Noguchi, H. Identification of four allelopathic compounds including a novel compound from Elaeocarpus floribundus Blume and determination of their allelopathic activity. J. Environ. Manag. 2023, 326, 116728. [Google Scholar] [CrossRef]
- Bari, I.N.; Kato-Noguchi, H.; Iwasaki, A.; Suenaga, K. Allelopathic Potency and an Active Substance from Anredera cordifolia (Tenore) Steenis. Plants 2019, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Rob, M.M.; Iwasaki, A.; Suzuki, R.; Suenaga, K.; Kato-Noguchi, H. Garcienone, a novel compound involved in allelopathic activity of Garcinia xanthochymus Hook. Plants 2019, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Lun, T.L.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Two Allelopathic Substances from Plumbago rosea Stem Extracts and Their Allelopathic Effects. Agronomy 2022, 12, 2020. [Google Scholar] [CrossRef]
- Deng, J.; Liao, Z.; Chen, D. Marsdenosides A–H, polyoxypregnane glycosides from Marsdenia tenacissima. Phytochemistry 2005, 66, 1040–1051. [Google Scholar] [CrossRef]
- Wang, S.; Lai, Y.-H.; Tian, B.; Yang, L. Two New C21 Steroidal Glycosides from Marsdenia tenacissima (ROXB.) WIGHT et ARN. Chem. Pharm. Bull. 2006, 54, 696–698. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; To, K.K.-W.; Wang, Y.-Z.; Yin, C.; Tang, C.; Chai, S.; Ke, C.-Q.; Lin, G.; Ye, Y. Polyoxypregnane Steroids from the Stems of Marsdenia tenacissima. J. Nat. Prod. 2014, 77, 2044–2053. [Google Scholar] [CrossRef]
- Warashina, T.; Noro, T. Steroidal glycosides from the aerial part of Asclepias incarnata. Phytochemistry 2000, 53, 485–498. [Google Scholar] [CrossRef]
- Liu, P.; Xu, D.-W.; Li, R.-T.; Wang, S.-H.; Hu, Y.-L.; Shi, S.-Y.; Li, J.-Y.; Huang, Y.-H.; Kang, L.-W.; Liu, T.-X. A Combined Phytochemistry and Network Pharmacology Approach to Reveal Potential Anti-NSCLC Effective Substances and Mechanisms in Marsdenia tenacissima (Roxb.) Moon (Stem). Front. Pharmacol. 2021, 12, 518406. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Anticancer properties of caudatin and related C-21 steroidal glycosides from Cynanchum plants. Steroids 2021, 172, 108855. [Google Scholar] [CrossRef]
- Matsuo, Y.; Shinoda, D.; Nakamaru, A.; Kamohara, K.; Sakagami, H.; Mimaki, Y. Steroidal Glycosides from Convallaria majalis Whole Plants and Their Cytotoxic Activity. Int. J. Mol. Sci. 2017, 18, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.-Q.; Zhang, R.-R.; Wang, J.; Ma, Y.; Li, W.-X.; Jiang, R.W.; Cai, S.-H. Asclepiasterol, a novel C21 steroidal glycoside derived from Asclepias curassavica, reverses tumor multidrug resistance by down-regulating P-glycoprotein expression. Oncotarget 2016, 7, 31466–31483. [Google Scholar] [CrossRef] [PubMed]
- Timité, G.; Mitaine-Offer, A.-C.; Miyamoto, T.; Tanaka, C.; Mirjolet, J.-F.; Duchamp, O.; Lacaille-Dubois, M.-A. Structure and cytotoxicity of steroidal glycosides from Allium schoenoprasum. Phytochemistry 2013, 88, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Kenji, G.M.; Nakajima, S.; Baba, N.; Iwasa, J. Biologically Active Substance from Kenyan Plant, Vernonia hindii S. Moore (Asteraceae); Scientific Report of the Faculty of Agriculture, Okayama University: Okayama, Japan, 1998; Volume 87, pp. 17–21. [Google Scholar]
- Liu, X.-J.; Shi, Y.; Jia, S.-H.; Deng, Y.-L.; Lv, F.; Dai, R.-J. Six new C-21 steroidal glycosides from Dregea sinensis Hemsl. J. Asian Nat. Prod. Res. 2017, 19, 745–753. [Google Scholar] [CrossRef]
- Panda, N.; Sukdeb Banerjee, S.; Mandal, N.B.; Sahu, N.P. Pregnane glycosides. Nat. Prod. Com. 2006, 1, 665–695. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Plant | I50 Value (mg DW Equivalent Extract/mL) | ||
|---|---|---|---|
| Shoot | Root | ||
| Dicots | Lettuce | 3.8 d | 3.2 d |
| Alfalfa | 3.5 d | 3.9 d | |
| Monocots | Barnyard grass | 54.2 a | 12.6 db |
| Timothy | 9.1 c | 0.7 e | |
| Position | δC, (Type) a | δH (J in Hz) b | COSY/TOCSY | Selected HMBC | Selected NOESY |
|---|---|---|---|---|---|
| 1a | 39.0, CH2 | 1.22, m | 2a, 2b | 18 | |
| 1b | 1.56, m | 2b | 18 | ||
| 2a | 30.7, CH2 | 1.28, m | 1a, 1b, 3 | ||
| 2b | 1.77, m | 1a, 1b, 3 | |||
| 3 | 77.9, CH | 3.61, m | 2a, 2b, 4a, 4b | ||
| 4a | 36.0, CH2 | 1.22, m | 3, 5 | ||
| 4b | 1.66, m | 3, 5 | |||
| 5 | 45.7, CH | 1.23, m | 4a, 4b, 6a, 6b | ||
| 6a | 30.2, CH2 | 1.32, m | 5, 7a, 7b | ||
| 6b | 1.38, m | 5, 7a, 7b | |||
| 7a | 29.1, CH2 | 1.16, m | 6a, 6b, 8 | ||
| 7b | 2.13, m | 6a, 6b, 8 | |||
| 8 | 40.8, CH | 1.77, m | 7a, 7b, 9 | 14 | |
| 9 | 51.0, CH | 1.45, m | 8, 11 | ||
| 10 | 38.7, C | ||||
| 11 | 72.5, CH | 5.82, dd (10.1, 10.1) | 9, 12 | 1′ | 18, 19 |
| 12 | 78.7, CH | 4.86, d (10.1) | 11 | 1″ | |
| 13 | 55.6, C | ||||
| 14 | 85.2, C | ||||
| 15a | 34.4, CH2 | 1.87, m | 16a, 16b | 14 | |
| 15b | 2.11, m | 16a, 16b | 14 | ||
| 16a | 24.9, CH2 | 1.99, m | 15a, 15b, 17 | ||
| 16b | 2.08, m | 15a, 15b, 17 | |||
| 17 | 59.0, CH | 2.89, dd (4.8, 9.6) | 16a, 16b | ||
| 18 | 12.6, CH3 | 0.94, s | 1, 5, 9, 10 | 1a, 1b, 11 | |
| 19 | 11.7, CH3 | 1.04, s | 12, 13, 14, 17 | 11, 21 | |
| 20 | 216.0, C | ||||
| 21 | 32.0, CH3 | 2.12, s | 17, 20 | 19 | |
| Acetyl | |||||
| 1′ | 172.1, C | ||||
| 2′ | 21.6, CH3 | 1.82, s | 1′ | ||
| Tigloyl | |||||
| 1″ | 169.1, C | ||||
| 2″ | 129.1, C | ||||
| 3″ | 140.6, CH | 6.93, brq | 4″ | 2″ | |
| 4″ | 14.6, CH3 | 1.85, brs | 3″ | ||
| 5″ | 12.1, CH3 | 1.85, s | 1″, 2″ | ||
| Cym I | |||||
| 1′″ | 98.6, CH | 4.64, dd (1.6, 9.6) | 2′″a, 2′″b | 3 | |
| 2′″a | 37.9, CH2 | 1.34, m | 1′″, 3′″ | ||
| 2′″b | 2.23, m | 1′″, 3′″ | |||
| 3′″ | 80.5, CH | 3.37, m | 2′″, 4′″ | ||
| 4′″ | 84.0, CH | 3.17, m | 3′″, 5′″ | ||
| 5′″ | 72.5, CH | 3.66, m | 4′″, 6′″ | ||
| 6′″ | 18.8, CH3 | 1.35, d (5.9) | 5′″ | ||
| 13′″ | 57.4, CH3 | 3.40, s | 3′″ | ||
| Cym II | |||||
| 7′″ | 102.2, CH | 4.72, d (8.1) | 8′″ | 4′″ | |
| 8′″ | 73.6, CH | 3.31, m | 7′″, 9′″ | ||
| 9′″ | 84.0, CH | 3.62, m | 8′″, 10′″ | ||
| 10′″ | 75.0, CH | 3.17, m | 9′″, 11′″ | ||
| 11′″ | 71.3, CH | 3.37, m | 10′″, 12′″ | ||
| 12′″ | 18.2, CH3 | 1.23, d (6.3) | 11′″ | ||
| 14′″ | 62.5, CH3 | 3.59, s | 9′″ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moh, S.M.; Kurisawa, N.; Suenaga, K.; Kato-Noguchi, H. Allelopathic Potential of Marsdenia tenacissima (Roxb.) Moon against Four Test Plants and the Biological Activity of Its Allelopathic Novel Compound, 8-Dehydroxy-11β-O-Acetyl-12β-O-Tigloyl-17β-Marsdenin. Plants 2023, 12, 1663. https://doi.org/10.3390/plants12081663
Moh SM, Kurisawa N, Suenaga K, Kato-Noguchi H. Allelopathic Potential of Marsdenia tenacissima (Roxb.) Moon against Four Test Plants and the Biological Activity of Its Allelopathic Novel Compound, 8-Dehydroxy-11β-O-Acetyl-12β-O-Tigloyl-17β-Marsdenin. Plants. 2023; 12(8):1663. https://doi.org/10.3390/plants12081663
Chicago/Turabian StyleMoh, Seinn Moh, Naoaki Kurisawa, Kiyotake Suenaga, and Hisashi Kato-Noguchi. 2023. "Allelopathic Potential of Marsdenia tenacissima (Roxb.) Moon against Four Test Plants and the Biological Activity of Its Allelopathic Novel Compound, 8-Dehydroxy-11β-O-Acetyl-12β-O-Tigloyl-17β-Marsdenin" Plants 12, no. 8: 1663. https://doi.org/10.3390/plants12081663