
Effects of Ca Sprays on Fruit Ca Content and Yield of Tomato Variety Susceptible to Blossom-End Rot

Abstract
:1. Introduction
2. Materials and Methods
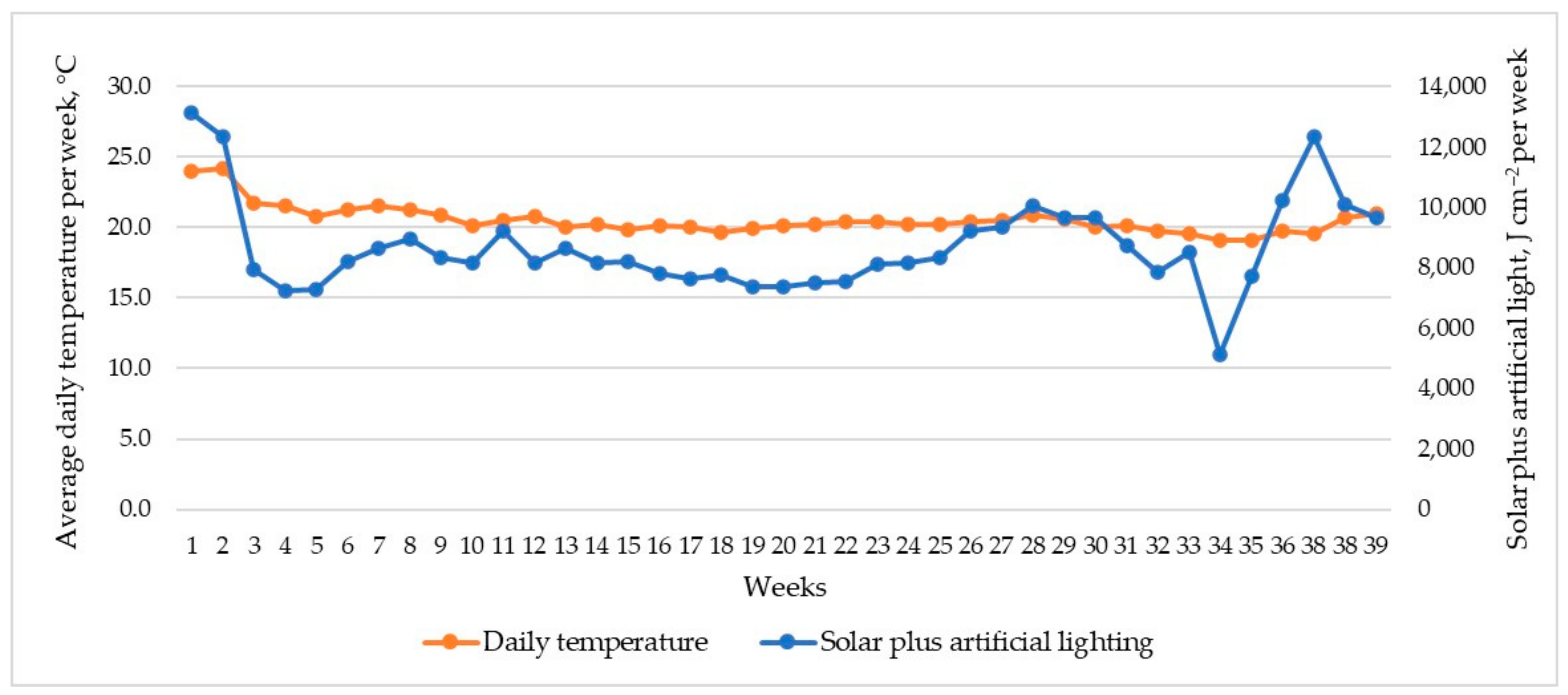
2.1. Study Site and Plant Growth Conditions
2.2. Ca Preparations and Plant Treatments
- Control—deionized water;
- An aqueous 0.2% solution of Brexil Duo (Ca 12.9%, Mg 2.4%, B 0.5%, Cu 0.5%, Mn 2%, Zn 2%; water-soluble foliar fertiliser, nutrients complexed with lignin sulphonate, Valagro SDS, Atessa, Italy);
- An aqueous 0.2% solution of Greenstim (Ca 0%, a preparation promoting Ca uptake, 97% glycine betaine; Verdera, Espoo, Finland);
- An aqueous 0.5% solution of Calmax zero N (Ca 7.13%, B 0.33%, Cu 0.04%, Fe 0.05%, Mn 0.1%, Mo 0.001%, Zn 0.02%; concentrated suspension containing CaO, micronutrients chelated with EDTA; Omex Agrifluids Ltd., King’s Lynn, UK);
- An aqueous 0.5% solution of Ca(NO3)2 (Ca 19%; Yara Latvia Ltd., Riga, Latvia);
- An aqueous 0.5% solution of CaCl2 (Ca 28%; Yara Latvia Ltd., Riga, Latvia).
2.3. Sampling and Tissue Analysis
2.4. Measurement of Yield Parameters
2.5. Statistical Analysis
3. Results
3.1. Nutrient Status
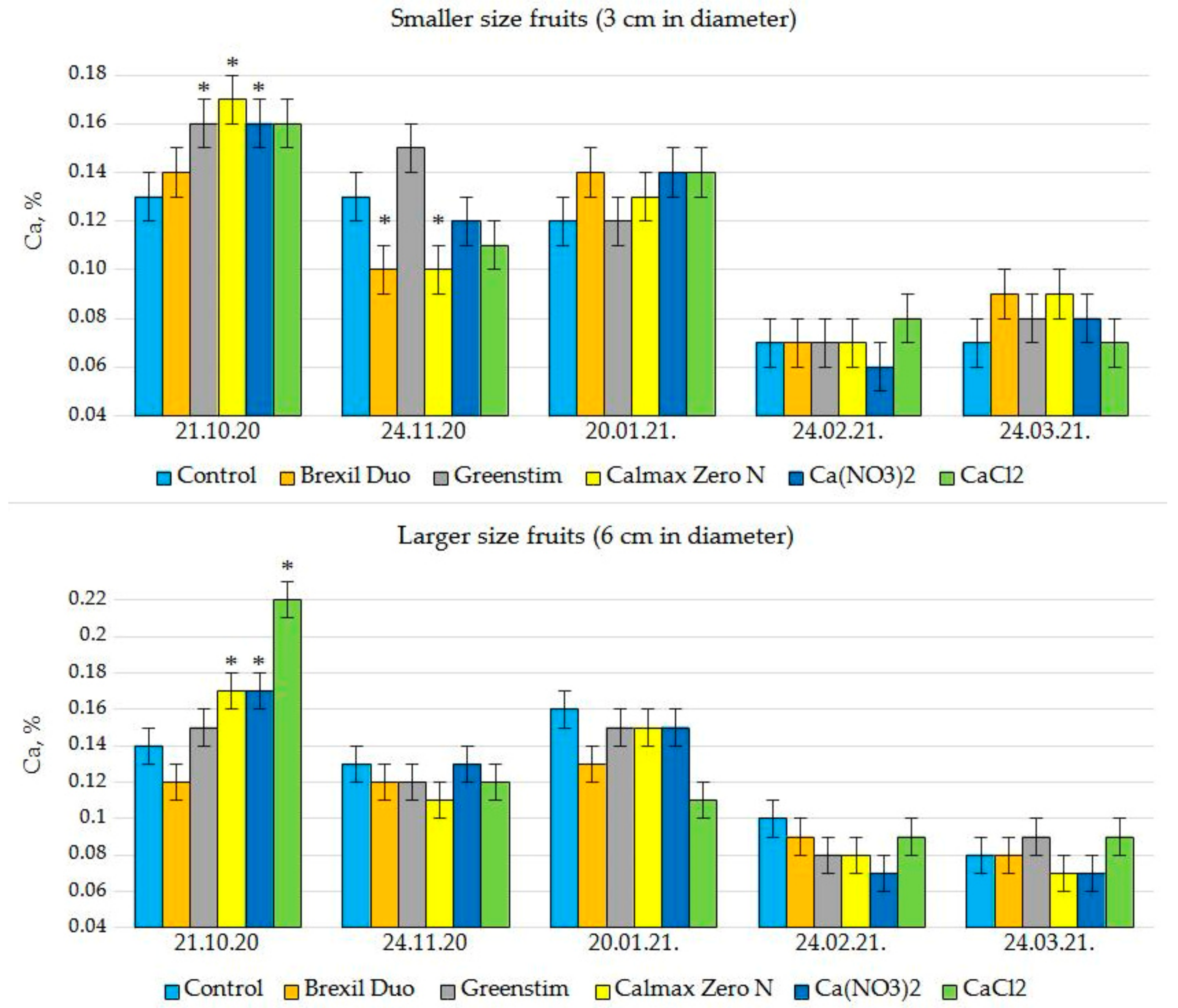
3.2. Ca Content in Tomato Fruits
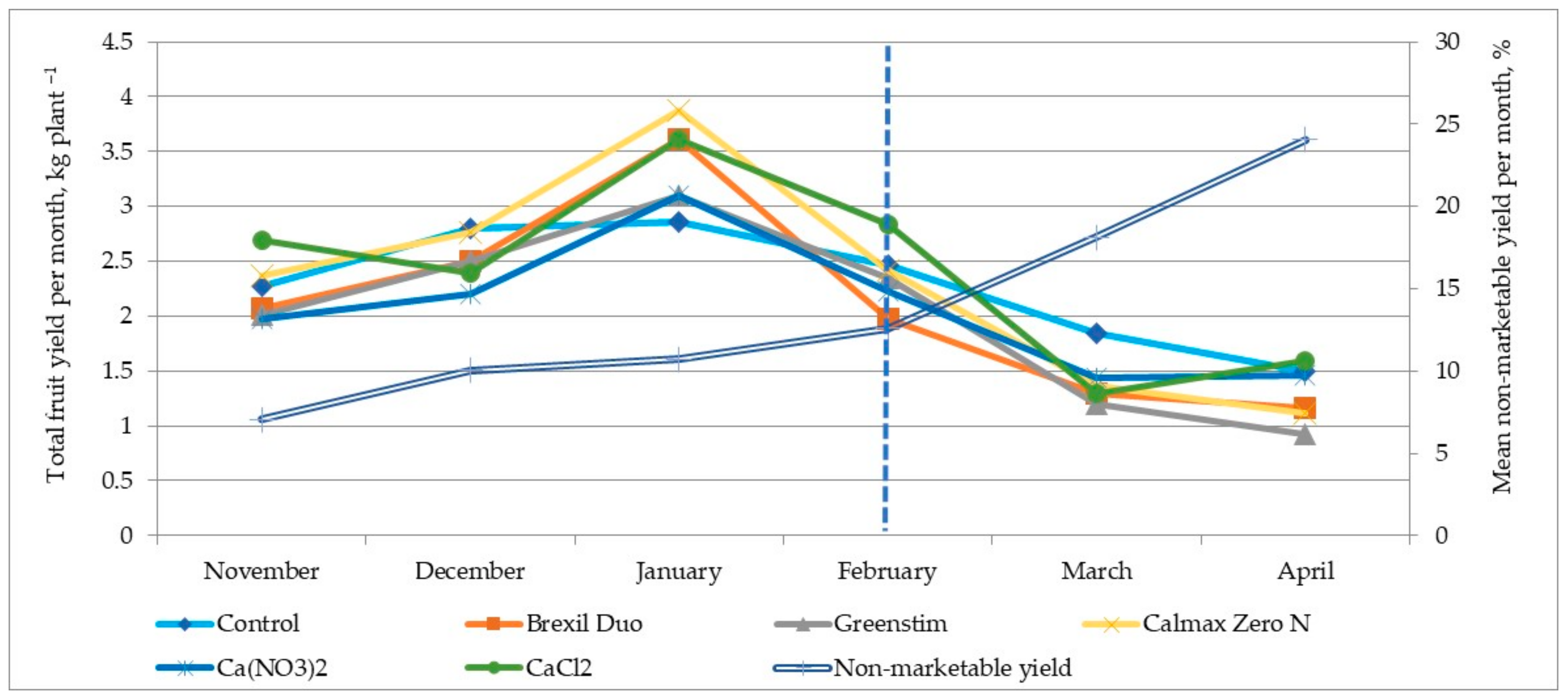
3.3. Yield and BER Incidence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hagassou, D.; Francia, E.; Ronga, D.; Buti, M. Blossom end-rot in tomato (Solanum lycopersicum L.): A multi-disciplinary overview of inducing factors and control strategies. Sci. Hortic. 2019, 249, 49–58. [Google Scholar] [CrossRef]
- Olle, M.; Williams, I.H. Physiological disorders in tomato and some methods to avoid them. J. Hortic. Sci. Biotechnol. 2017, 92, 223–230. [Google Scholar] [CrossRef]
- Saure, M.C. Blossom-end rot of tomato (Lycopersicon esculentum Mill.)—A calcium or a stress-related disorder? Sci. Hortic. 2001, 90, 193–208. [Google Scholar] [CrossRef]
- Riboldi, L.B.; da Cruz Araújo, S.H.; de Freitas, S.T.; Castro, P.R.C. Fruit shape regulates susceptibility of tomato to blossom-end rot. Acta Sci. Agron. 2020, 42, e42487. [Google Scholar] [CrossRef] [Green Version]
- De Freitas, S.T.; Mitcham, E.J. Factors involved in fruit calcium deficiency disorders. Hort. Rev. 2012, 40, 107–146. [Google Scholar] [CrossRef]
- Ho, L.C.; White, P.J. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Ann. Bot. 2005, 95, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. (Ed.) Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Saure, M.C. Why calcium deficiency is not the cause of blossom-end rot in tomato and pepper fruit—A reappraisal. Sci. Hortic. 2014, 174, 151–154. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [Green Version]
- Jürkenbeck, K.; Spiller, A.; Meyerding, S.G.H. Tomato attributes and consumer preferences—A consumer segmentation approach. Br. Food J. 2019, 122, 328–344. [Google Scholar] [CrossRef]
- Birlanga, V.; Acosta-Motos, J.R.; Pérez-Pérez, J.M. Mitigation of calcium-related disorders in soilless production systems. Agronomy 2022, 12, 644. [Google Scholar] [CrossRef]
- Vinh, T.D.; Yoshida, Y.; Ooyama, M.; Goto, T.; Yasuba, K.; Tanaka, Y. Comparative analysis on blossom-end rot incidence in two tomato cultivars in relation to calcium nutrition and fruit growth. Hortic. J. 2018, 87, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.G.; Klee, H.; Tieman, D. Field evaluation of CRISPR-Cas9-driven brachytic and jointless pedicel tomatoes identifies an association between the high extra-large-sized fruit yield of the brachytic-mediated shortened tomato and the jointless2. Hortic. Environ. Biotechnol. 2022, 63, 1–6. [Google Scholar] [CrossRef]
- Sierra-Orozco, E.; Shekasteband, R.; Illa-Berenguer, E.; Snouffer, A.; van der Knaap, E.; Lee, T.G.; Hutton, S.F. Identification and characterization of GLOBE, a major gene controlling fruit shape and impacting fruit size and marketability in tomato. Hortic. Res. 2021, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Reitz, N.F.; Shackel, K.A.; Mitcham, E.J. Differential effects of excess calcium applied to whole plants vs. excised fruit tissue on blossom-end rot in tomato. Sci. Hortic. 2021, 290, 110514. [Google Scholar] [CrossRef]
- Liebisch, F.; Max, J.F.J.; Heine, G.; Horst, W.J. Blossom-end rot and fruit cracking of tomato grown in net-covered greenhouses in Central Thailand can partly be corrected by calcium and boron sprays. J. Plant. Nutr. Soil Sci. 2009, 172, 140–150. [Google Scholar] [CrossRef]
- Alonso, T.A.S.; Barreto, R.F.; Prado, R.M.; Souza Junior, J.P.; Carvalho, R.F. Silicon spraying alleviates calcium deficiency in tomato plants, but Ca-EDTA is toxic. J. Plant Nutr. Soil Sci. 2020, 183, 659–664. [Google Scholar] [CrossRef]
- Mazumder, M.N.N.; Misran, A.; Ding, P.; Wahab, P.E.M.; Mohamad, A. Preharvest foliar spray of calcium chloride on growth, yield, quality, and shelf life extension of different lowland tomato varieties in Malaysia. Horticulturae 2021, 7, 466. [Google Scholar] [CrossRef]
- Peet, M.M. Irrigation and fertilization. In Tomatoes; Heuvelink, E., Ed.; CABI: Wallingford, UK, 2005; pp. 171–198. [Google Scholar]
- Rab, A.; Haq, I.U. Foliar application of calcium chloride and borax influences plant growth, yield, and quality of tomato (Lycopersicon esculentum Mill.) fruit. Turk. J. Agric. For. 2012, 36, 695–701. [Google Scholar] [CrossRef]
- Cekstere, G.; Osvalde, A.; Karlsons, A.; Strauts, G. Nutrient status of tomatoes grown in hydroponics during the crop cycle as a factor involved in fruit yield and quality. Acta Hortic. 2021, 1321, 109–116. [Google Scholar] [CrossRef]
- Dry Farm Tomato Variety Trial 2020: High Performing Variety Descriptions. Available online: https://horticulture.oregonstate.edu/sites/agscid7/files/2._tomato_variety_trial_report_jan_31_2021.pdf (accessed on 8 February 2023).
- Heuvelink, E. (Ed.) Tomatoes, 2nd ed.; Crop Production Science in Horticulture Series, 27; CABI International: Oxfordshire, UK, 2018; p. 388.
- Microwave Plasma Atomic Emission Spectroscopy (MP-AES). Application eHandbook, Agilent Technologies. Available online: https://www.agilent.com/cs/library/applications/5991-7282EN_MP-AES-eBook.pdf (accessed on 23 July 2020).
- Cekstere, G.; Osvalde, A.; Elferts, D.; Rose, C.; Lucas, F.; Vollenweider, P. Salt accumulation and effects within foliage of Tilia x vulgaris trees from the street greenery of Riga, Latvia. Sci. Total Environ. 2020, 747, 140921. [Google Scholar] [CrossRef]
- Nutritional Recommendations for Tomato. Available online: https://www.haifa-group.com/crop-guide/vegetables/tomato/crop-guide-tomato-plant-nutrition (accessed on 15 March 2020).
- Brust, G. Nutrient Problems and Their Management in Tomatoes, University of Marlyand Extension. Available online: https://extension.umd.edu/sites/extension.umd.edu/files/2021-06/NutrientProblemsAndTheirMgtInTomatoes.pdf (accessed on 13 December 2020).
- Campbell, C.R. (Ed.) Tomato, greenhouse. In Reference Sufficiency Ranges for Plant Analysis in the Southern Region of the United States; Southern Region Agricultural Experiment Station: Fayetteville, AR, USA, 2000; Volume 394, pp. 79–80. [Google Scholar]
- Fanasca, S.; Rouphael, Y.; Cardarelli, M.; Colla, G. The influence of K:Ca:Mg:Na ratio and total concentration on yield and fruit quality of soilless-grown tomatoes: A modelling approach. Acta Hortic. 2005, 697, 345–350. [Google Scholar] [CrossRef]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Pujos, A.; Morard, P. Effects of potassium deficiency on tomato growth and mineral nutrition at the early production stage. Plant Soil 1997, 189, 189–196. [Google Scholar] [CrossRef]
- Bar-Tal, A.; Yermiyahu, U.; Ben-Gal, A.; Schwartz, A.; Faingold, I.; Seligmann, R. Optimization of calcium and magnesium concentrations for fertigation of tomato with desalinated water. Isr. J. Plant Sci. 2017, 64, 80–91. [Google Scholar]
- Hao, H.; Papadopoulos, A.P. Effects of calcium and magnesium on growth, fruit yield and quality in a fall greenhouse tomato crop grown on rockwool. Can. J. Plant Sci. 2003, 83, 903–912. [Google Scholar]
- Pečinar, I.; Quarrie, S.P.; Bertin, N.; Rančič, D.; Savič, S.; Jovanovič, Z.; Stikič, R. Tomato fruit development in response to different irrigation practices: Developmental study of pericarp cell layers. Biol. Life Sci. Forum 2021, 4, 105. [Google Scholar]
- Rached, M.; Pierre, B.; Yves, G.; Matsukura, C.; Ariizumi, T.; Ezura, H.; Fukuda, N. Differences in blossom-end rot resistance in tomato cultivars is associated with total ascorbate rather than calcium concentration in the distal end part of fruits per se. Hortic. J. 2018, 87, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Melo, R.O.; Martinez, H.E.P.; Rocha, B.C.P.; Garcia, J.E. Production and quality of sweet grape tomato in response to foliar calcium fertilization. Rev. Ceres 2022, 69, 48–54. [Google Scholar] [CrossRef]
- Farm Solution Catalogue 2023. Available online: https://valagrocatalogue.com/ (accessed on 15 March 2023).
- Peyvast, G.; Olfati, J.A.; Ramezani-Kharazi, P.; Kamari-Shahmalek, S. Uptake of calcium nitrate and potassium phosphate from foliar fertilization by tomato. J. Hortic. For. 2009, 1, 7–13. [Google Scholar]
- Mäkelä, P.; Jokinen, K.; Kontturi, M.; Peltonen-Sainio, P.; Pehu, E.; Somersalo, S. Foliar application of glycinebetaine—A novel product from sugar beet—As an approach to increase tomato yield. Ind. Crops Prod. 1998, 7, 139–148. [Google Scholar] [CrossRef]
- Li, M.; Zhi, H. Influence of preharvest and postharvest applications of glycine betaine on fruit quality attributes and storage disorders of ‘Lapins’ and ‘Regina’ cherries. Hortscience 2019, 59, 1540–1545. [Google Scholar] [CrossRef] [Green Version]
- Zuzunaga-Rosas, J.; González-Orenga, S.; Tofei, A.M.; Boscaiu, M.; Moreno-Ramón, H.; Ibáñez-Asensio, S.; Vicente, O. Effect of a biostimulant based on polyphenols and glycine betaine on tomato plants’ responses to salt stress. Agronomy 2022, 12, 2142. [Google Scholar] [CrossRef]
- Indeche, A.K.; Yoshida, Y.; Goto, T.; Yasuba, K.; Yoshiyuki, T. Effect of defoliation on blossom-end rot incidence and calcium transport into fruit of tomato cultivars under moderate water stress. Hortic. J. 2020, 89, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Indeche, A.K.; Yoshida, Y.; Miyaji, D.; Goto, T.; Yasuba, K.; Tanakac, Y. Defoliating to 12–15 leaves increases calcium concentration and decreases blossom-end rot incidence in fruit of tomato plant grown under moderate water stress. Sci. Rep. Fac. Agric. Okayama Univ. 2020, 109, 21–27. [Google Scholar]
- De Freitas, S.T.; Jiang, C.Z.; Mitcham, E.J. Mechanisms involved in calcium deficiency development in tomato fruit in response to gibberellins. J. Plant Growth Regul. 2012, 31, 221–234. [Google Scholar] [CrossRef]
- De Freitas, S.T.; McElrone, A.J.; Shackel, K.A.; Mitcham, E.J. Calcium partitioning and allocation and blossom-end rot development in tomato plants in response to whole-plant and fruit-specific abscisic acid treatments. J. Exp. Bot. 2014, 65, 235–247. [Google Scholar] [CrossRef]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Foliar applications of abscisic acid decrease the incidence of blossom-end rot in tomato fruit. Sci. Hortic. 2014, 179, 356–362. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nutrient | Young Leaves (n = 14) | Older Leaves (n = 14) | Sufficiency Ranges in Tomato Leaves | ||
|---|---|---|---|---|---|
| Range | Mean ± SE | Range | Mean ± SE | ||
| N | 3.50–5.15 | 4.01 ± 0.15b 1 | 3.20–4.00 | 3.44 ± 0.07a | 3.50–6.00 |
| P | 0.46–0.66 | 0.52 ± 0.02a | 0.57–0.89 | 0.76 ± 0.04b | 0.30–1.00 |
| K | 2.69–6.50 | 4.25 ± 0.29a | 3.07–7.20 | 4.91 ± 0.39a | 3.50–6.00 |
| Ca | 1.55–5.59 | 2.87 ± 0.32a | 4.95–7.30 | 6.13 ± 0.17b | 1.20–4.00 |
| Mg | 0.27–0.55 | 0.37 ± 0.02a | 0.25–0.58 | 0.41 ± 0.03a | 0.30–1.00 |
| S | 0.70–2.08 | 1.28 ± 0.10a | 1.88–3.63 | 2.67 ± 0.18b | 0.40–1.00 |
| Nutrient | Young Leaves (n = 14) | Old Leaves (n = 14) | Sufficiency Ranges in Tomato Leaves | ||
|---|---|---|---|---|---|
| Range | Mean ± SE | Range | Mean ± SE | ||
| Fe | 133–323 | 209.38 ± 16.15a 1 | 180–549 | 326.36 ± 38.05b | 100–300 |
| Mn | 68–132 | 93.77 ± 5.22a | 127–300 | 188.45 ± 14.05b | 50–200 |
| Zn | 14.0–31.5 | 19.79 ± 1.26a | 9.5–35.0 | 19.18 ± 2.15a | 25–80 |
| Cu | 10.5–22.0 | 16.43 ± 1.04b | 6.5–21.5 | 11.30 ± 1.38a | 6.0–25.0 |
| Mo | 1.50–5.00 | 3.40 ± 0.23a | 2.88–8.00 | 4.83 ± 0.47b | 1.0–5.0 |
| B | 24–54 | 32.62 ± 2.51a | 38–84 | 60.73 ± 4.46b | 25–75 |
| N | P | K | Ca | Mg | S | Fe | Mn | Zn | Cu | Mo | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | 0.835 * | 1.000 | |||||||||
| K | 0.577 * | 0.627 * | 1.000 | ||||||||
| Ca | 0.742 * | 0.459 | 0.522 * | 1.000 | |||||||
| Mg | 0.808 * | 0.486 | 0.443 | 0.798 * | 1.000 | ||||||
| S | 0.567 * | 0.648 * | 0.616 * | 0.656 * | 0.423 | 1.000 | |||||
| Fe | −0.434 | −0.161 | 0.282 | −0.430 | −0.455 | 0.059 | 1.000 | ||||
| Mn | −0.522 | −0.486 | −0.257 | −0.420 | −0.536 * | −0.271 | 0.420 | 1.000 | |||
| Zn | −0.106 | −0.056 | −0.046 | −0.142 | 0.117 | −0.167 | 0.143 | 0.190 | 1.000 | ||
| Cu | 0.237 | 0.345 | 0.193 | 0.184 | 0.347 | 0.190 | −0.090 | −0.293 | 0.636 * | 1.000 | |
| Mo | 0.501 * | 0.644 * | 0.247 | 0.252 | 0.214 | 0.677 * | 0.072 | −0.278 | 0.084 | 0.186 | 1.000 |
| B | 0.352 | 0.442 | 0.475 | 0.334 | 0.198 | 0.366 | 0.279 | −0.115 | −0.246 | 0.130 | 0.095 |
| N | P | K | Ca | Mg | S | Fe | Mn | Zn | Cu | Mo | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P | −0.402 | 1.000 | |||||||||
| K | −0.048 | 0.772 * | 1.000 | ||||||||
| Ca | 0.186 | 0.284 | 0.547 | 1.000 | |||||||
| Mg | 0.606 * | −0.596 * | −0.255 | −0.134 | 1.000 | ||||||
| S | 0.028 | 0.719 * | 0.747 * | 0.060 | −0.254 | 1.000 | |||||
| Fe | −0.165 | 0.790 * | 0.862 * | 0.275 | −0.064 | 0.666 * | 1.000 | ||||
| Mn | 0.301 | 0.334 | 0.555 * | 0.289 | 0.172 | 0.367 | 0.535 * | 1.000 | |||
| Zn | 0.290 | 0.086 | 0.374 | 0.176 | 0.508 * | 0.399 | 0.354 | 0.407 | 1.000 | ||
| Cu | 0.372 | 0.374 | 0.600 * | 0.227 | 0.367 | 0.699 * | 0.638 * | 0.583 * | 0.735 * | 1.000 | |
| Mo | −0.355 | 0.451 | 0.324 | −0.058 | −0.455 | 0.259 | 0.264 | 0.045 | 0.131 | −0.064 | 1.000 |
| B | −0.180 | 0.763 * | 0.512 | 0.334 | −0.616 | 0.531 | 0.424 | 0.273 | −0.027 | 0.342 | 0.474 |
| Treatment | Mean Total Yield, kg Plant−1 | Compared to the Control, % |
|---|---|---|
| Control | 15.31 ± 1.07 | 100 |
| Brexil Duo 0.2% | 13.65 ± 1.09 | 89 |
| Greenstim 0.2% | 13.62 ± 1.12 | 89 |
| Calmax Zero N 0.5% | 15.16 ± 1.39 | 99 |
| Ca(NO3)2 0.5% | 13.74 ± 1.04 | 90 |
| CaCl2 0.5% | 15.83 ± 1.29 | 103 |
| Nutrient | Smaller Size Fruits (3 cm in Diameter) | Larger Size Fruits (6 cm in Diameter) | ||
|---|---|---|---|---|
| October–January | February–March | October–January | February–March | |
| K | 4.10 ± 0.18a 1 | 4.33 ± 0.10a | 4.01 ± 0.19a | 4.39 ± 0.14a |
| Mg | 0.20 ± 0.01a | 0.19 ± 0.01a | 0.17 ± 0.01a | 0.17 ± 0.01a |
| Ca | 0.13 ± 0.005b | 0.08 ± 0.003a | 0.14 ± 0.006b | 0.08 ± 0.003a |
| Treatment | Smaller Size Fruits (3 cm in Diameter) | Larger Size Fruits (6 cm in Diameter) | ||
|---|---|---|---|---|
| October–January | February–March | October–January | February–March | |
| Control | 34.19 ± 4.46a 1 A 2 | 71.64 ± 1.79abB | 29.03 ± 2.97aA | 54.90 ± 1.10aB |
| Brexil Duo, 0.2% | 33.89 ± 0.97aA | 56.82 ± 9.04aB | 32.30 ± 4.01aA | 52.83 ± 3.95aB |
| Greenstim, 0.2% | 30.64 ± 5.36aA | 64.68 ± 6.18aB | 30.32 ± 2.62aA | 56.85 ± 5.40aB |
| Calmax Zero N, 0.5% | 32.08 ± 3.13aA | 54.36 ± 9.36aB | 29.07 ± 0.89aA | 53.85 ± 6.72aB |
| Ca(NO3)2, 0.5% | 30.60 ± 2.25aA | 60.65 ± 8.02aB | 27.36 ± 1.56aA | 65.50 ± 2.36bB |
| CaCl2, 0.5% | 32.18 ± 2.04aA | 58.93 ± 2.93aB | 30.74 ± 6.46aA | 49.78 ± 4.33aB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karlsons, A.; Osvalde, A.; Cekstere, G.; Āboliņa, L. Effects of Ca Sprays on Fruit Ca Content and Yield of Tomato Variety Susceptible to Blossom-End Rot. Plants 2023, 12, 1640. https://doi.org/10.3390/plants12081640
Karlsons A, Osvalde A, Cekstere G, Āboliņa L. Effects of Ca Sprays on Fruit Ca Content and Yield of Tomato Variety Susceptible to Blossom-End Rot. Plants. 2023; 12(8):1640. https://doi.org/10.3390/plants12081640
Chicago/Turabian StyleKarlsons, Andis, Anita Osvalde, Gunta Cekstere, and Laura Āboliņa. 2023. "Effects of Ca Sprays on Fruit Ca Content and Yield of Tomato Variety Susceptible to Blossom-End Rot" Plants 12, no. 8: 1640. https://doi.org/10.3390/plants12081640