
Effects of Exogenous Sodium Nitroprusside Spraying on Physiological Characteristics of Soybean Leaves at the Flowering Stage under Drought Stress
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
Plant Materials and Test Design
3. Test Methods
3.1. Determination of the NO Content
3.2. Determination of Antioxidant Enzyme Activity
3.3. Determination of Contents of Osmoregulatory Substances
3.4. Determination of NiR and NR Activity
3.5. Determination of MDA Content
3.6. Data Analysis
4. Results and Analysis
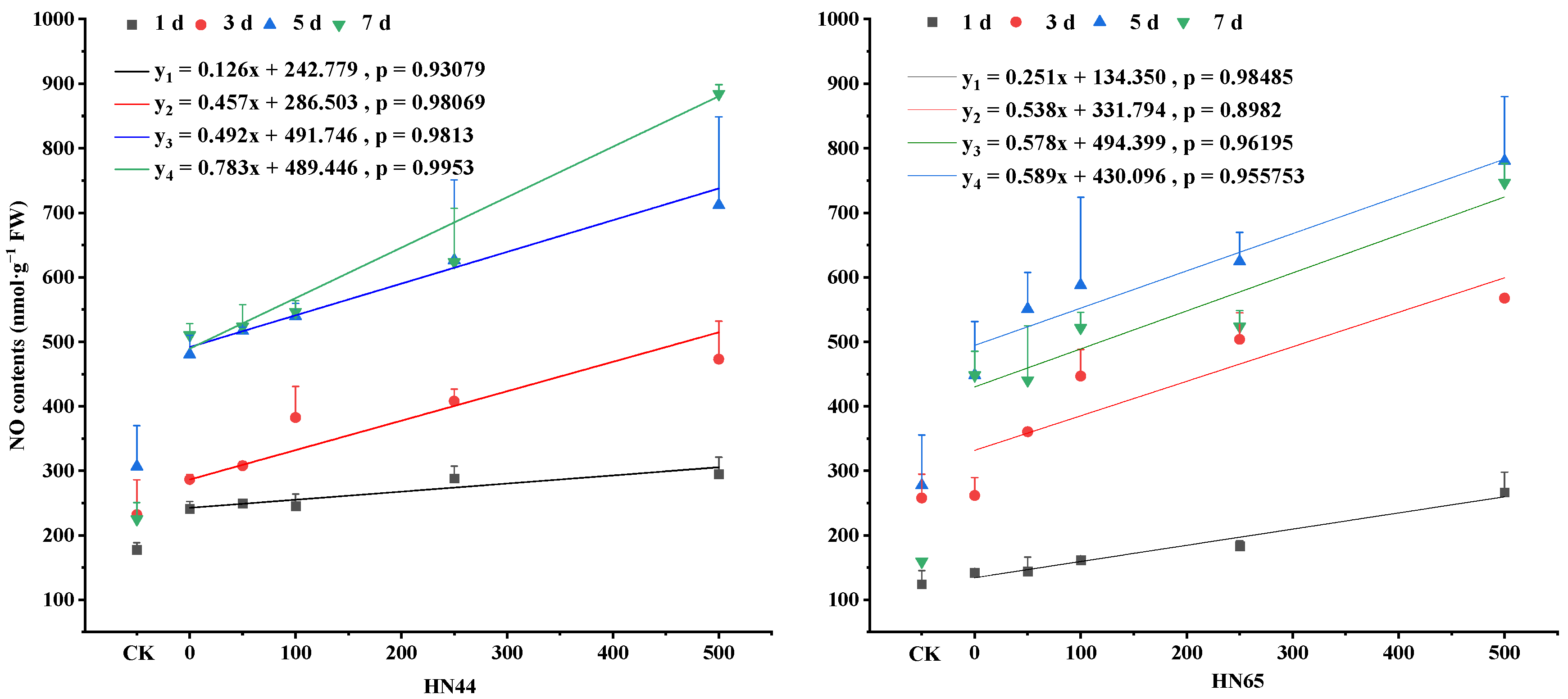
4.1. Changes in the NO Content in Soybean Leaves
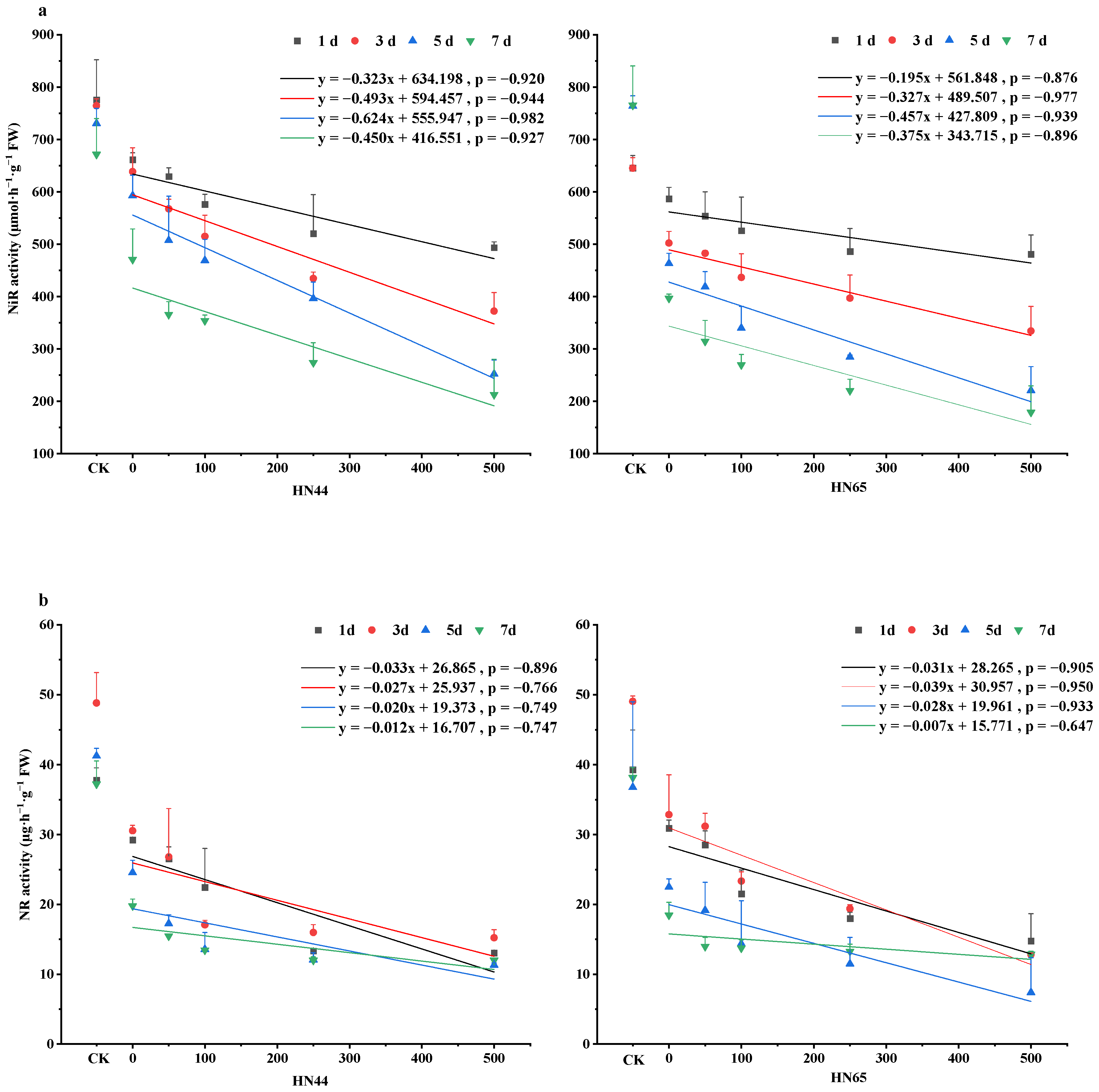
4.2. Changes in NiR and NR Activity in Soybean Leaves
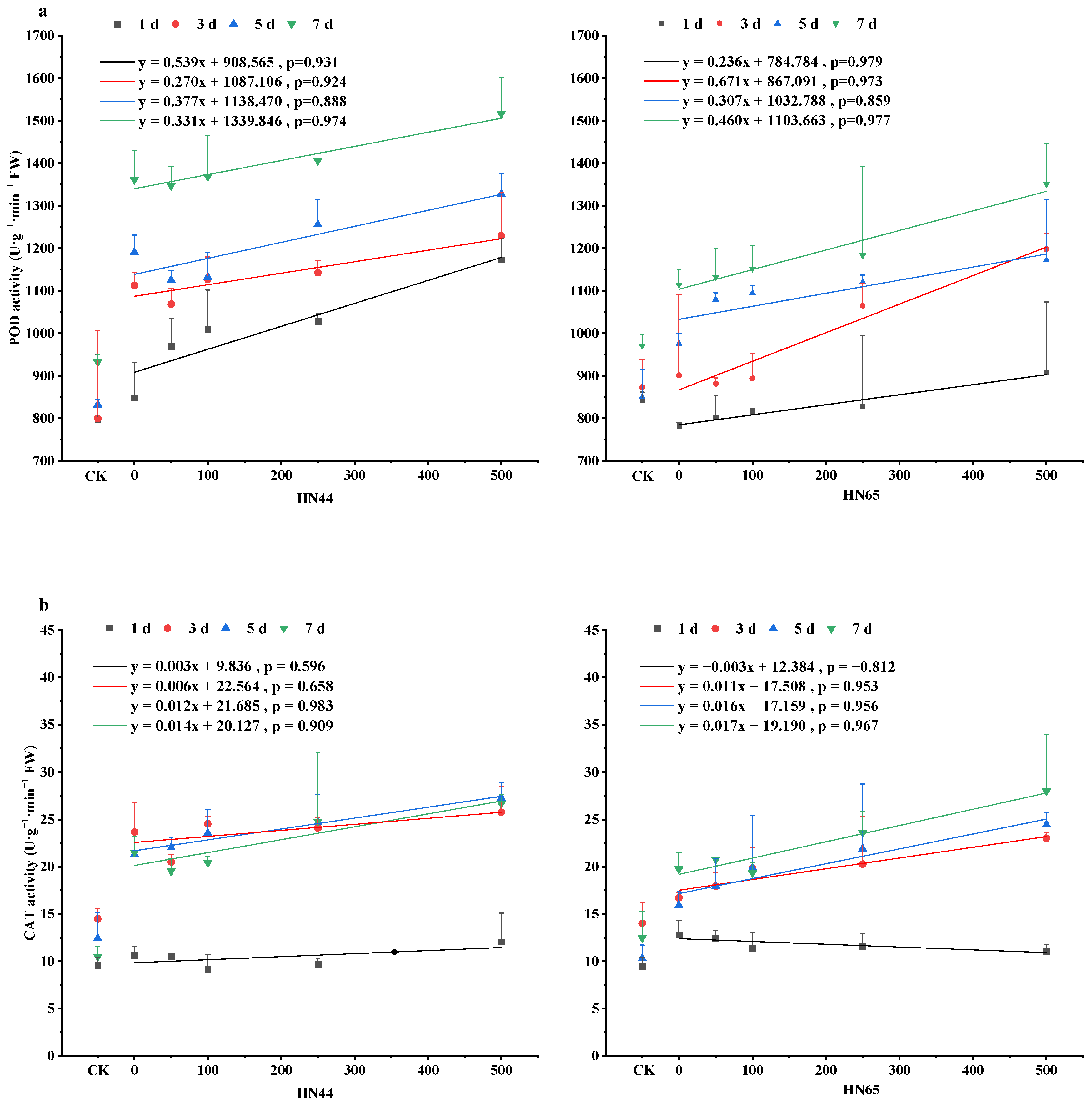
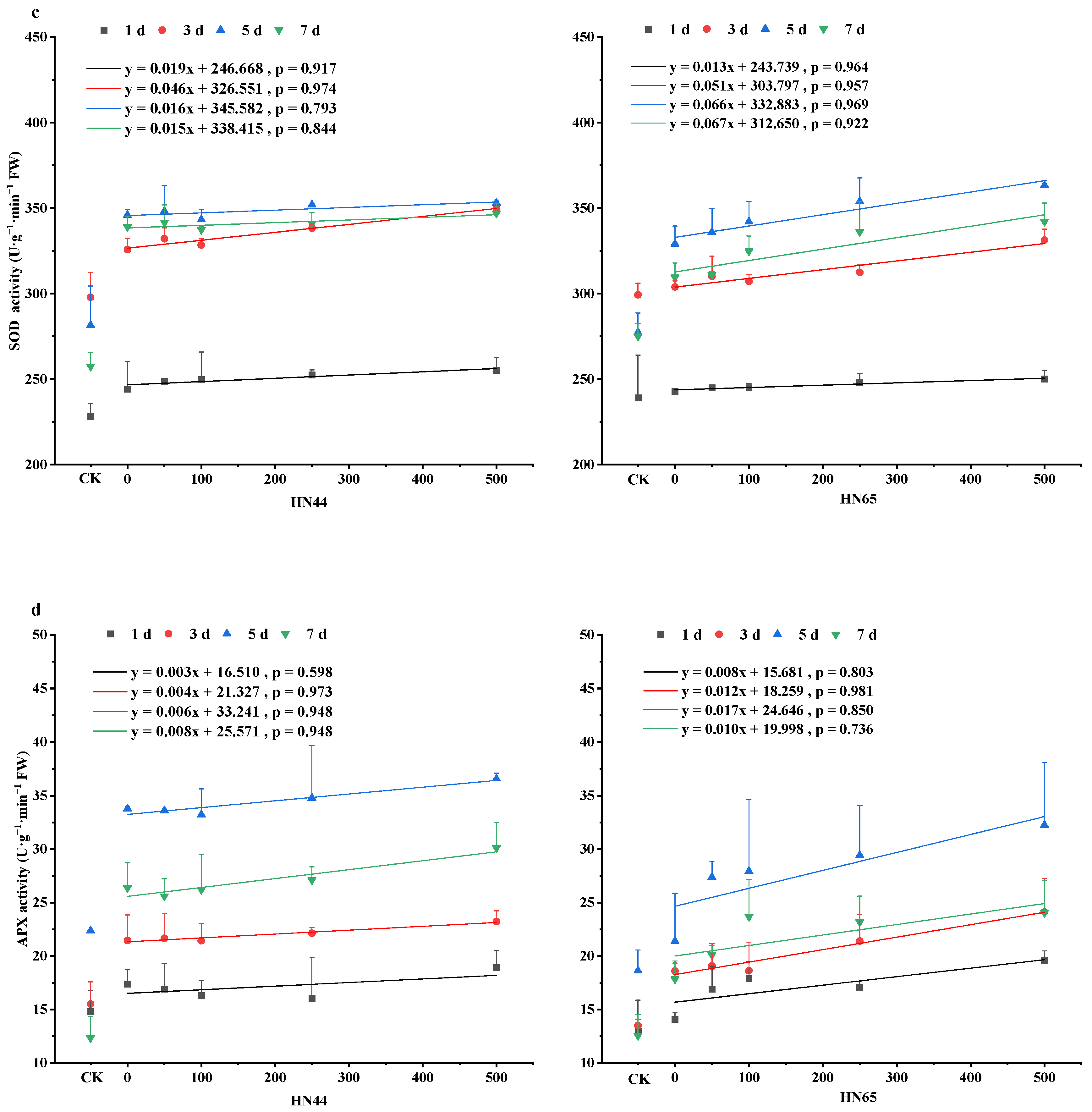
4.3. Changes in Antioxidant Enzyme Activity in Soybean Leaves
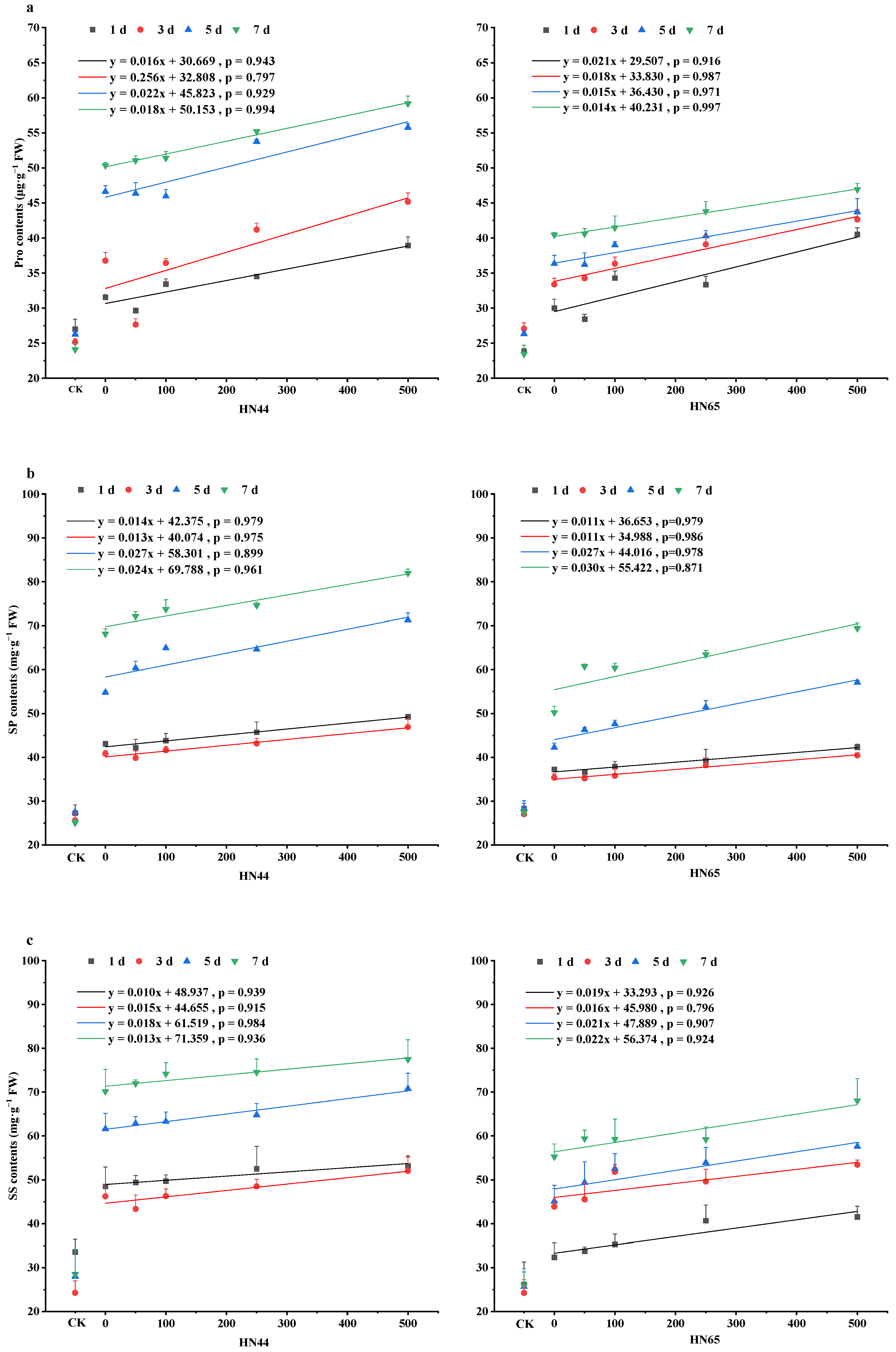
4.4. Changes in Osmotic Adjustment Ability in Soybean Leaves
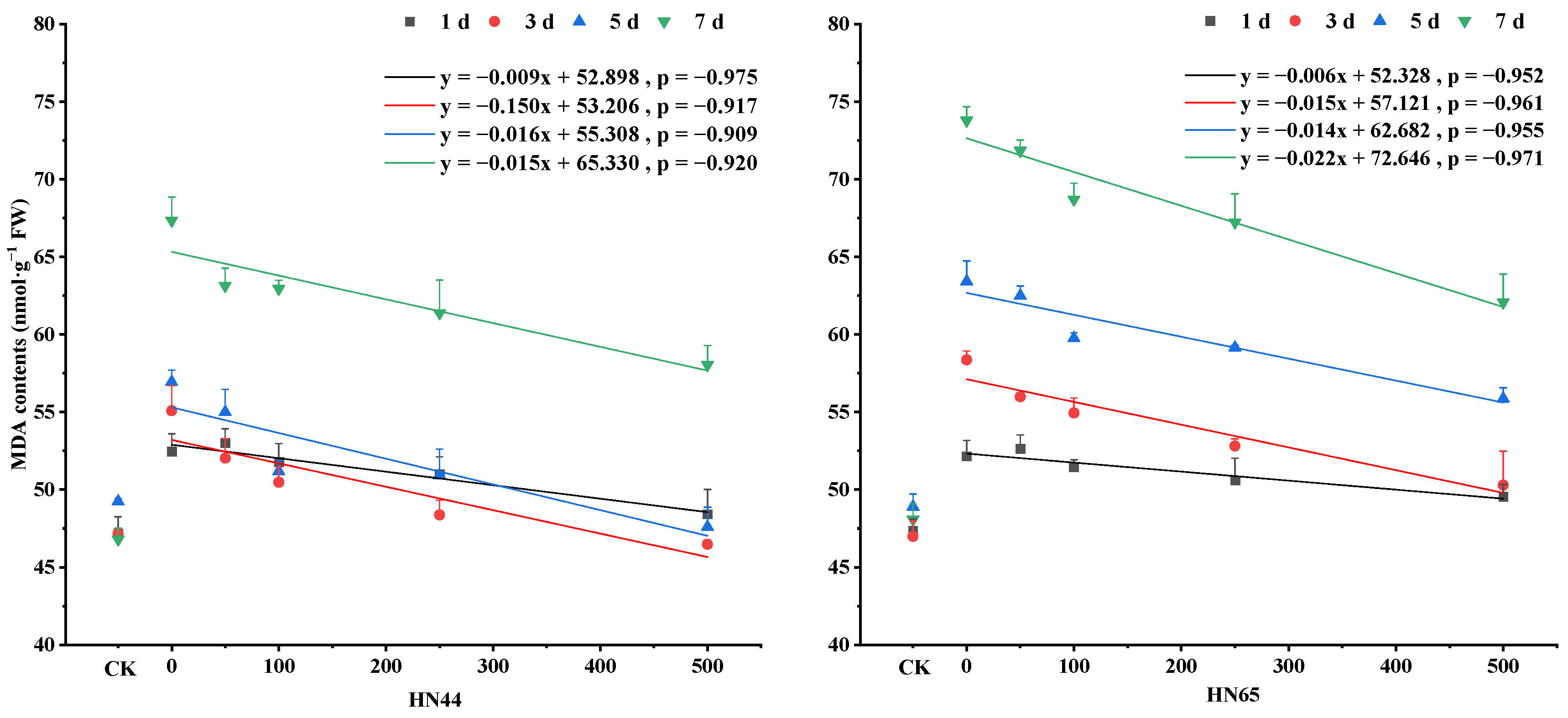
4.5. Changes in MDA Contents under Different SNP Treatments
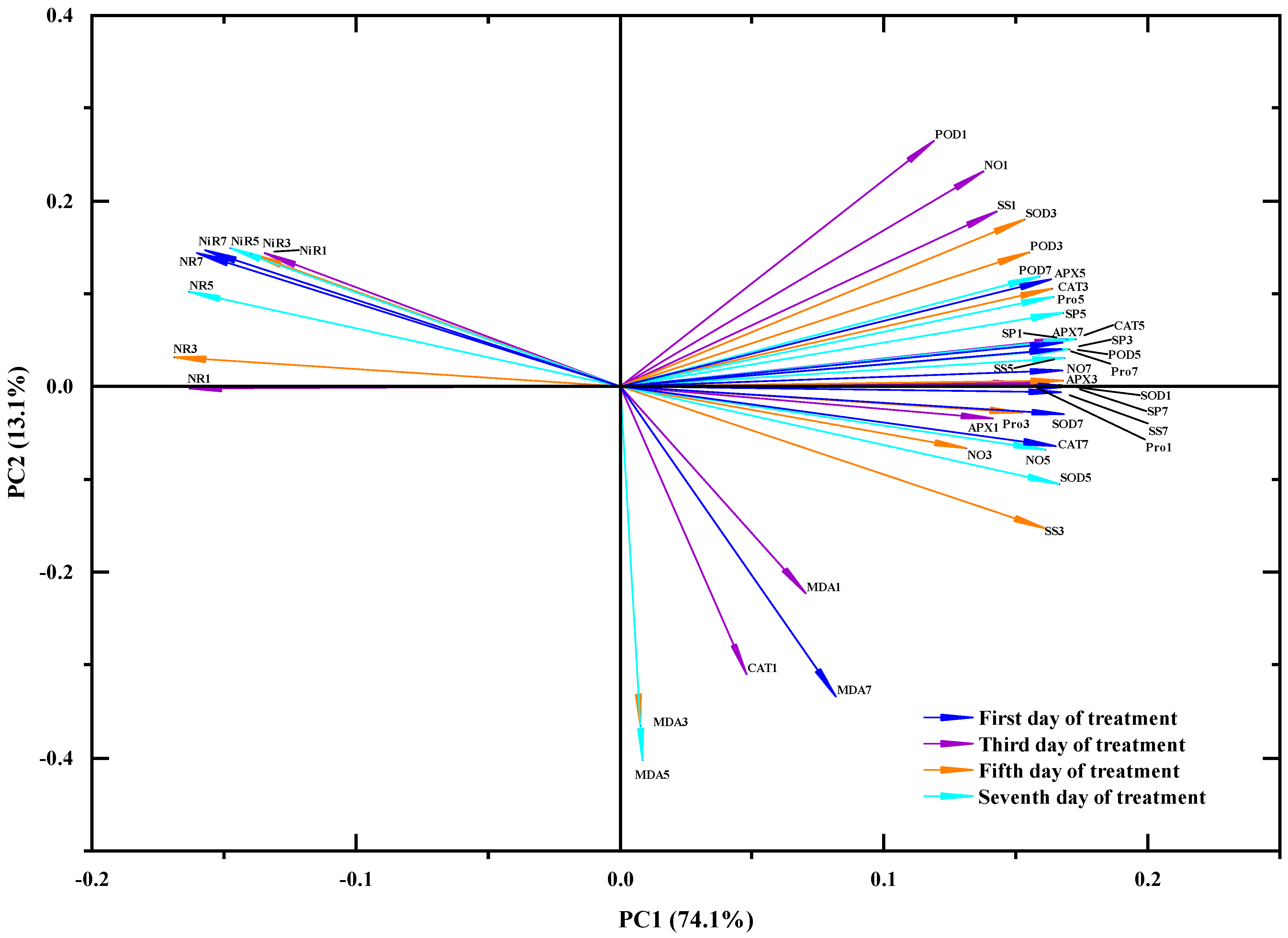
4.6. Principal Component Analysis(PCA) and Multifactor Analysis of Variance
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. Disasters accompanied by new threats have severely impacted the agricultural food system. In World Agriculture; FAO: Rome, Italy, 2021; pp. 104–105. [Google Scholar]
- Dong, S.; Jiang, Y.; Dong, Y.; Wang, L.; Wang, W.; Ma, Z.; Yan, C.; Ma, C.; Liu, L. A study on soybean responses to drought stress and rehydration. Saudi J. Biol. Sci. 2019, 26, 2006–2017. [Google Scholar] [CrossRef]
- Rasheed, A.; Mahmood, A.; Maqbool, R.; Albaqami, M.; Sher, A.; Sattar, A.; Bakhsh, G.; Nawaz, M.; Hassan, M.U.; Al-Yahyai, R.; et al. Key insights to develop drought-resilient soybean: A review. J. King Saud Univ.-Sci. 2022, 34, 102089. [Google Scholar] [CrossRef]
- Mohd Amnan, M.A.; Pua, T.L.; Lau, S.E.; Tan, B.C.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Komatsu, S. Osmotic stress in banana is relieved by exogenous nitric oxide. PeerJ 2021, 9, e10879. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.-E.; Hamdan, M.F.; Pua, T.-L.; Saidi, N.B.; Tan, B.C. Plant Nitric Oxide Signaling under Drought Stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef]
- Jinghui, G.A.O.; Yangxiu, M.U.; Yueli, Z.; Zhisong, G.E.; Wen, Y.A.N.; Huaimei, S. Effects of Exogenous Nitric Oxide on the Growth and Physiological Characteristics of Perennial Ryegrass Seedlings under Osmotic Stress. Acta Agrestia Sin. 2011, 19, 625–630. [Google Scholar]
- Guo, X.; Huang, D.; Jing, G.; Feng, J.; Zhu, S. Nitric oxide-mediated DNA methylation enhances cold resistance in postharvest peach fruit. Food Chem. 2022, 404 Pt B, 134660. [Google Scholar] [CrossRef]
- Kovacs, I.; Lindermayr, C. Nitric oxide-based protein modification: Formation and site-specificity of protein S-nitrosylation. Front. Plant Sci. 2013, 4, 137. [Google Scholar] [CrossRef] [Green Version]
- Egbichi, I.; Keyster, M.; Jacobs, A.; Klein, A.; Ludidi, N. Modulation of antioxidant enzyme activities and metabolites ratios by nitric oxide in short-term salt stressed soybean root nodules. S. Afr. J. Bot. 2013, 88, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Corpas, F.J.; Ahmad, P. Nitric oxide, salicylic acid and oxidative stress: Is it a perfect equilateral triangle? Plant Physiol. Biochem. 2022, 184, 56–64. [Google Scholar] [CrossRef]
- Zhou, Q.; Song, S.; Wang, X.; Yan, C.; Ma, C.M.; Dong, S.K. Effects of drought stress on flowering soybean physiology under different soil conditions. Plant Soil Environ. 2022, 68, 487–498. [Google Scholar] [CrossRef]
- Imran, M.; Shazad, R.; Bilal, S.; Imran, Q.M.; Khan, M.; Kang, S.-M.; Khan, A.L.; Yun, B.-W.; Lee, I.-J. Exogenous Melatonin mediates the regulation of endogenous nitric oxide in Glycine max L. to reduce effects of drought stress. Environ. Exp. Bot. 2021, 188, 104511. [Google Scholar] [CrossRef]
- Alnusairi, G.S.H.; Mazrou, Y.S.A.; Qari, S.H.; Elkelish, A.A.; Soliman, M.H.; Eweis, M.; Abdelaal, K.; El-Samad, G.A.; Ibrahim, M.F.M.; ElNahhas, N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants 2021, 10, 1693. [Google Scholar] [CrossRef] [PubMed]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. NO to drought-multifunctional role of nitric oxide in plant drought: Do we have all the answers? Plant Sci. 2015, 239, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Procházková, D.; Haisel, D.; Pavlíková, D. Nitric oxide biosynthesis in plants—The short overview. Plant Soil Environ. 2014, 60, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Toffanin, A.; Wu, Q.; Maskus, M.; Caselia, S.; Abruna, H.D.; Shapleigh, J.P. Characterization of the gene encoding nitrite reductase and the physiological consequences of its expression in the nondenitrifying Rhizobium "hedysari" strain HCNT1. Appl. Environ. Microbiol. 1996, 62, 4019–4025. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Du, S. Properties of nitrogen fertilization are decisive in determining the effects of elevated atmospheric CO2 on the activity of nitrate reductase in plants. Plant Signal. Behav. 2016, 11, e1165380. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.F.; Wang, X.L.; Pan, J.; Deng, W.H.; Li, Y.C. Role of Nitrate Reductase and Nitrite Reductase in NaCl Tolerance in Eelgrass (Zostera marina L.). Ecol. Chem. Eng. S-Chem. I Inz. Ekol. S 2022, 29, 111–125. [Google Scholar] [CrossRef]
- Rosales, E.P.; Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Nitric oxide inhibits nitrate reductase activity in wheat leaves. Plant Physiol. Biochem. 2011, 49, 124–130. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Pei, Y.; Dong, S.; Sun, C.; Zu, W.; Ruan, Y. Drought resistance identification of soybean germplasm resources at bud stage. J. Northeast. Agric. Univ. 2012, 43, 36–43. [Google Scholar]
- Waheed, H.; Javaid, M.M.; Shahid, A.; Ali, H.H.; Nargis, J.; Mehmood, A. Impact of foliar-applied Hoagland’s nutrient solution on growth and yield of mash bean (Vigna mungo L.) under different growth stages. J. Plant Nutr. 2019, 42, 1133–1141. [Google Scholar] [CrossRef]
- Xu, M.J.; Dong, J.F.; Zhu, M.Y. Nitric oxide mediates the fungal elicitor-induced hypericin production of Hypericum perforatum cell suspension cultures through a jasmonic-acid-dependent signal pathway. Plant Physiol. 2005, 139, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kirkham, M.B. Drought-Stress-Induced Changes in Activities of Superoxide Dismutase, Catalase, and Peroxidase in Wheat Species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1980, 22, 867–880. [Google Scholar]
- Spychalla, J.P.; Desborough, S.L. Superoxide Dismutase, Catalase, and alpha-Tocopherol Content of Stored Potato Tubers. Plant Physiol. 1990, 94, 1214–1218. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Z.; Zhou, Q.; Wang, X.; Song, S.; Dong, S. Physiological Response of Soybean Plants to Water Deficit. Front. Plant Sci. 2021, 12, 809692. [Google Scholar] [CrossRef]
- Du, Y.L.; Zhao, Q.; Chen, L.R.; Yao, X.D.; Xie, F.T. Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Li, S.; Tian, X.; Duan, L.; Wang, B.; Zhi, Z.; He, Z. Effect of plant growth regulator SHK-6 on nitrogen metabolism of soybean leaf. Soybean Sci. 2004, 23, 15–20. [Google Scholar]
- Wang, Q. Effects of Cr6+ Stress on Membrane Lipid Peroxidation and Activities of Endogenous Protective Enzymes during Germination of Soybean Seed. Seed 2006, 25, 15–18. [Google Scholar]
- Majeed, S.; Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Ejaz, S.; Ahmad, K.S.; Tauseef, S.; Farid, G.; Khalid, I.; Mehmood, K. Nitric oxide regulates water status and associated enzymatic pathways to inhibit nutrients imbalance in maize (Zea mays L.) under drought stress. Plant Physiol. Biochem. 2020, 155, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Kanahama, K.; Kanayama, Y. Involvement of nitric oxide in the inhibition of nitrogenase activity by nitrate in Lotus root nodules. J. Plant Physiol. 2010, 167, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Choudary, P.V. Product-mediated regulation reveals the polymorphic nature of the yeast assimilatory nitrate reductase. Microbios 1993, 74, 53–57. [Google Scholar]
- Dinler, B.S.; Antoniou, C.; Fotopoulos, V. Interplay between GST and nitric oxide in the early response of soybean (Glycine max L.) plants to salinity stress. J. Plant Physiol. 2014, 171, 1740–1747. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.H.M.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Jing, G.; Zhu, S. Nitric oxide (NO) involved in antioxidant enzyme gene regulation to delay mitochondrial damage in peach fruit. Postharvest Biol. Technol. 2022, 192, 111993. [Google Scholar] [CrossRef]
- Khademi Astaneh, R.; Bolandnazar, S.; Zaare Nahandi, F. Exogenous nitric oxide protect garlic plants against oxidative stress induced by salt stress. Plant Stress 2022, 5, 100101. [Google Scholar] [CrossRef]
- Xu, M.; Li, H.; Liu, Z.N.; Wang, X.H.; Xu, P.; Dai, S.J.; Cao, X.; Cui, X.Y. The soybean CBL-interacting protein kinase, GmCIPK2, positively regulates drought tolerance and ABA signaling. Plant Physiol. Biochem. 2021, 167, 980–989. [Google Scholar] [CrossRef]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Morris, P.; Ribeiro, D.; Wilson, I. Nitric oxide, stomatal closure, and abiotic stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.-X.; Li, X.; Li, C.; Zhao, L. The Role of Nitric Oxide in Plant Responses to Salt Stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef]
- Ji, Z.; Camberato, J.J.; Zhang, C.; Jiang, Y. Effects of 6-Benzyladenine, γ-Aminobutyric Acid, and Nitric Oxide on Plant Growth, Photochemical Efficiency, and Ion Accumulation of Perennial Ryegrass Cultivars to Salinity Stress. HortScience 2019, 54, 1418–1422. [Google Scholar] [CrossRef] [Green Version]
- Hogg, N.; Kalyanaraman, B. Nitric oxide and lipid peroxidation. Biochim. Biophys. Acta 1999, 1411, 378–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundararajan, S.; Shanmugam, R.; Rajendran, V.; Sivakumar, H.P.; Ramalingam, S. Sodium Nitroprusside and Putrescine Mitigate PEG-Induced Drought Stress in Seedlings of Solanum lycopersicum. J. Soil Sci. Plant Nutr. 2022, 22, 1019–1032. [Google Scholar] [CrossRef]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Kubiś, J. Involvement of nitric oxide in water stress-induced responses of cucumber roots. Plant Sci. 2009, 177, 682–690. [Google Scholar] [CrossRef]
- Wei, X.-H.; Wang, L.-M.; Long, R.-J.; Wang, G.-X. Effects of exogenous nitric oxide, salicylic acid and hydrogen peroxide on free amino acid and soluble protein contents in tobacco leaves. Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao = J. Plant Physiol. Mol. Biol. 2006, 32, 257–260. [Google Scholar]
- Hao, G.-P.; Du, X.-H.; Shi, R.-J. Exogenous nitric oxide accelerates soluble sugar, proline and secondary metabolite synthesis in Ginkgo biloba under drought stress. Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao = J. Plant Physiol. Mol. Biol. 2007, 33, 499–506. [Google Scholar]
- Nemati, F.; Ghanati, F.; Gavlighi, H.A.; Sharifi, M. Fructan dynamics and antioxidant capacity of 4-day-old seedlings of wheat (Triticum aestivum) cultivars during drought stress and recovery. Funct. Plant Biol. 2018, 45, 1000–1008. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constituents | PEG-6000 | SNP (μmol/L) | |
|---|---|---|---|
| Treatment | |||
| CK | 0 | 0 | |
| S0 | 15% | 0 | |
| S50 | 15% | 50 | |
| S100 | 15% | 100 | |
| S250 | 15% | 250 | |
| S500 | 15% | 500 | |
| Different Treatments | Days | Different Treatments and Days | |||||
|---|---|---|---|---|---|---|---|
| Variety | Index | F | Sig | F | Sig | F | Sig |
| HN44 | NO | 173.001 | <0.01 | 396.733 | <0.01 | 18.203 | <0.01 |
| NiR | 51.955 | <0.01 | 33.215 | <0.01 | 0.768 | 0.705 | |
| NR | 166.089 | <0.01 | 24.92 | <0.01 | 3.143 | 0.001 | |
| POD | 33.903 | <0.01 | 44.348 | <0.01 | 1.162 | 0.333 | |
| CAT | 18.854 | <0.01 | 63.081 | <0.01 | 1.995 | 0.036 | |
| SOD | 45.224 | <0.01 | 256.572 | <0.01 | 3.313 | 0.001 | |
| APX | 25.104 | <0.01 | 116.587 | <0.01 | 2.415 | 0.011 | |
| Pro | 757.019 | <0.01 | 1001.968 | <0.01 | 58.858 | <0.01 | |
| SP | 750.399 | <0.01 | 1102.429 | <0.01 | 50.863 | <0.01 | |
| SS | 130.832 | <0.01 | 129.369 | <0.01 | 5.773 | <0.01 | |
| MDA | 82.765 | <0.01 | 188.203 | <0.01 | 10.8 | <0.01 | |
| HN65 | NO | 127.699 | <0.01 | 297.073 | <0.01 | 10.057 | <0.01 |
| NiR | 39.499 | <0.01 | 154.797 | <0.01 | 55.242 | <0.01 | |
| NR | 56.685 | <0.01 | 19.408 | <0.01 | 1.245 | 0.273 | |
| POD | 8.804 | <0.01 | 27.147 | <0.01 | 1.041 | 0.433 | |
| CAT | 7.782 | <0.01 | 16.526 | <0.01 | 0.942 | 0.527 | |
| SOD | 22.682 | <0.01 | 183.575 | <0.01 | 3.462 | 0.001 | |
| APX | 12.8 | <0.01 | 23.472 | <0.01 | 0.768 | 0.704 | |
| Pro | 257.096 | <0.01 | 107.024 | <0.01 | 7.367 | <0.01 | |
| SP | 750.399 | <0.01 | 1102.429 | <0.01 | 50.863 | <0.01 | |
| SS | 130.832 | <0.01 | 129.369 | <0.01 | 5.773 | <0.01 | |
| MDA | 194.069 | <0.01 | 459.815 | <0.01 | 18.867 | <0.01 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, Z.; Tian, Y.; Zhou, X.; Li, X.; Zhou, Q.; Wang, X.; Dong, S. Effects of Exogenous Sodium Nitroprusside Spraying on Physiological Characteristics of Soybean Leaves at the Flowering Stage under Drought Stress. Plants 2023, 12, 1598. https://doi.org/10.3390/plants12081598
Qu Z, Tian Y, Zhou X, Li X, Zhou Q, Wang X, Dong S. Effects of Exogenous Sodium Nitroprusside Spraying on Physiological Characteristics of Soybean Leaves at the Flowering Stage under Drought Stress. Plants. 2023; 12(8):1598. https://doi.org/10.3390/plants12081598
Chicago/Turabian StyleQu, Zhipeng, Yumei Tian, Xinyu Zhou, Xiaomei Li, Qi Zhou, Xiyue Wang, and Shoukun Dong. 2023. "Effects of Exogenous Sodium Nitroprusside Spraying on Physiological Characteristics of Soybean Leaves at the Flowering Stage under Drought Stress" Plants 12, no. 8: 1598. https://doi.org/10.3390/plants12081598