
Effects of Valine and Urea on Carbon and Nitrogen Accumulation and Lignin Content in Peach Trees
Abstract
:1. Introduction
2. Results
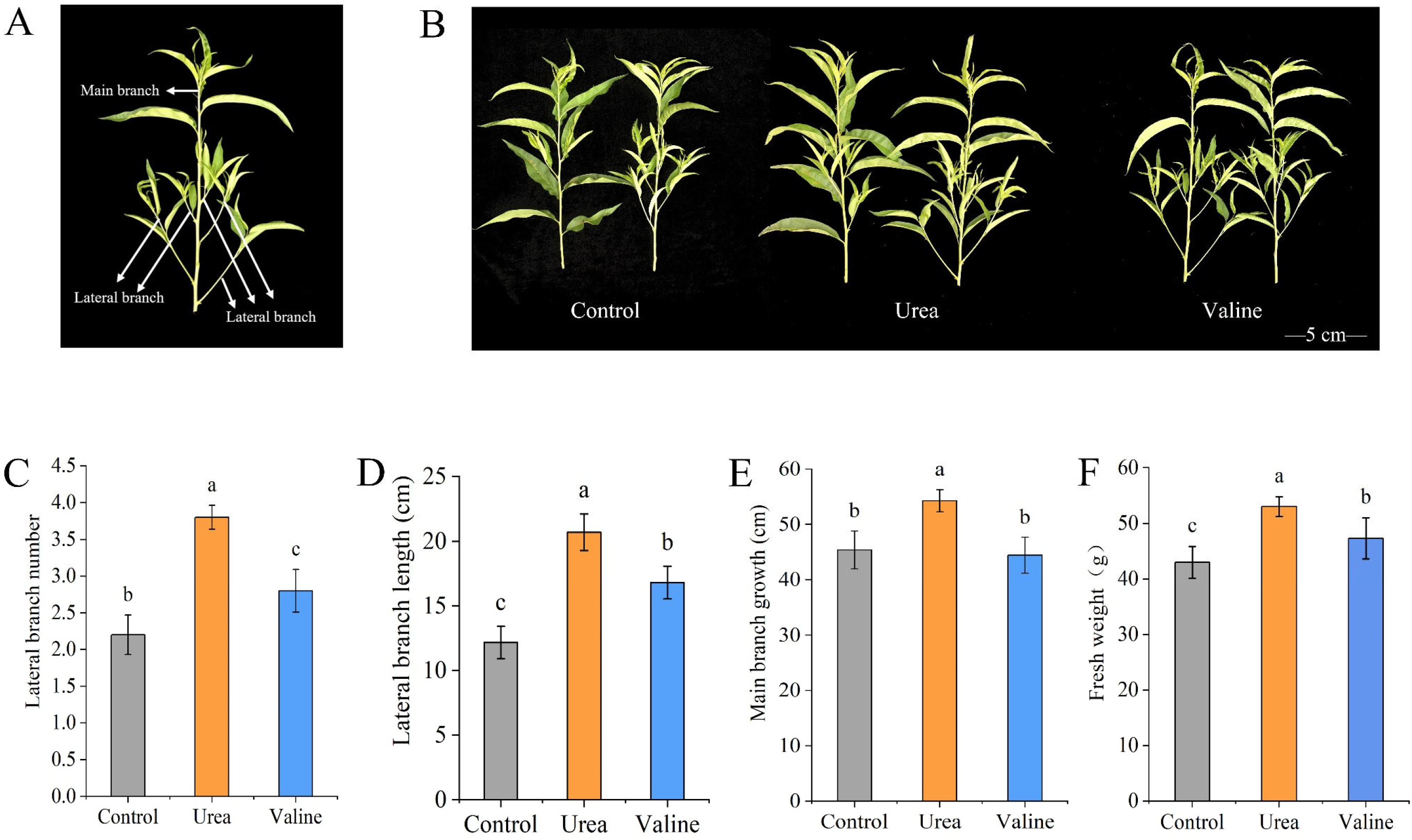
2.1. Effects of Valine and Urea on the Growth of New Shoots of Peach Trees
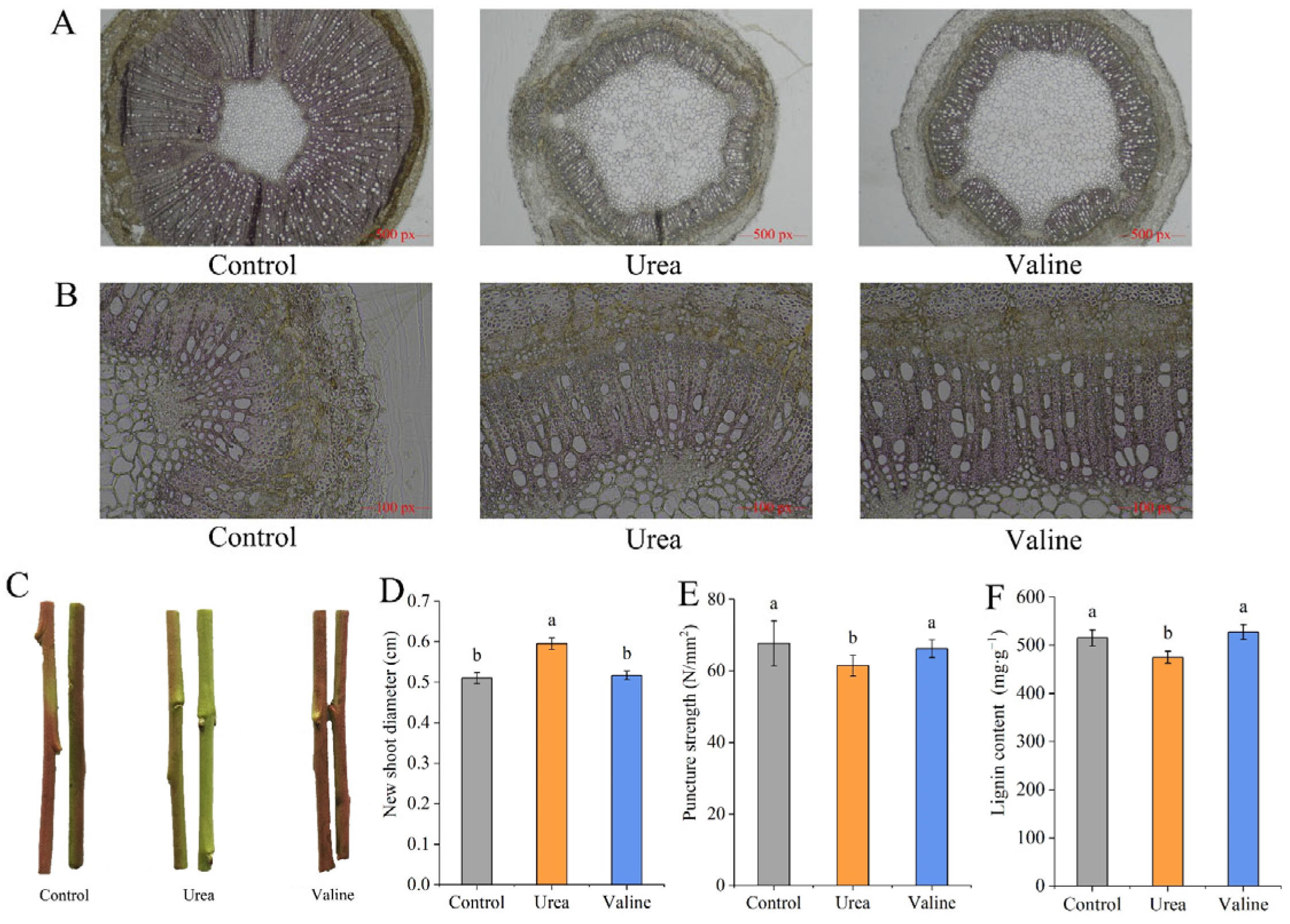
2.2. Effects of Valine and Urea on Lignin Content in New Shoots of Peach Trees
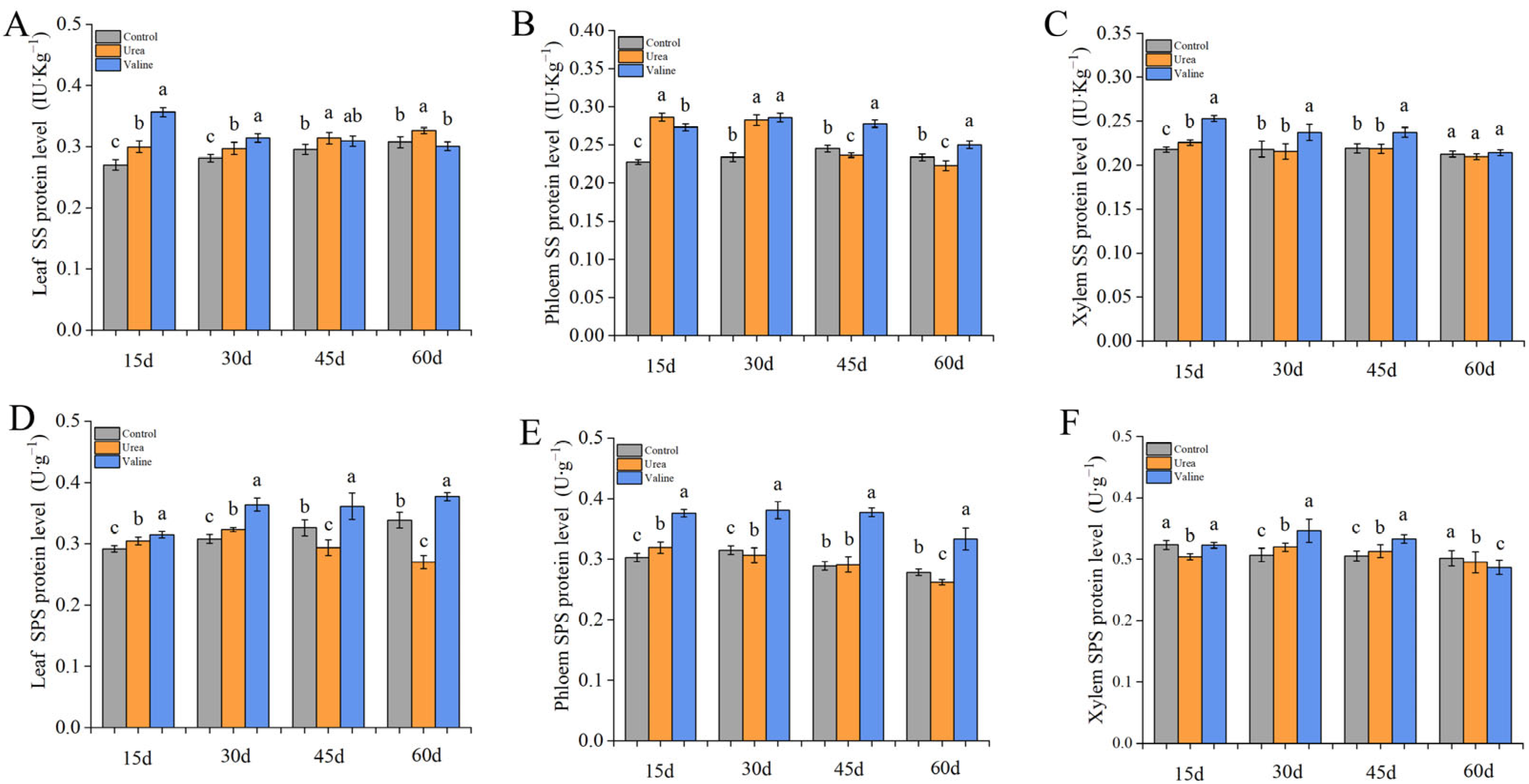
2.3. The Effect of Valine and Urea on Carbon Metabolism Enzymes in Peach Tree
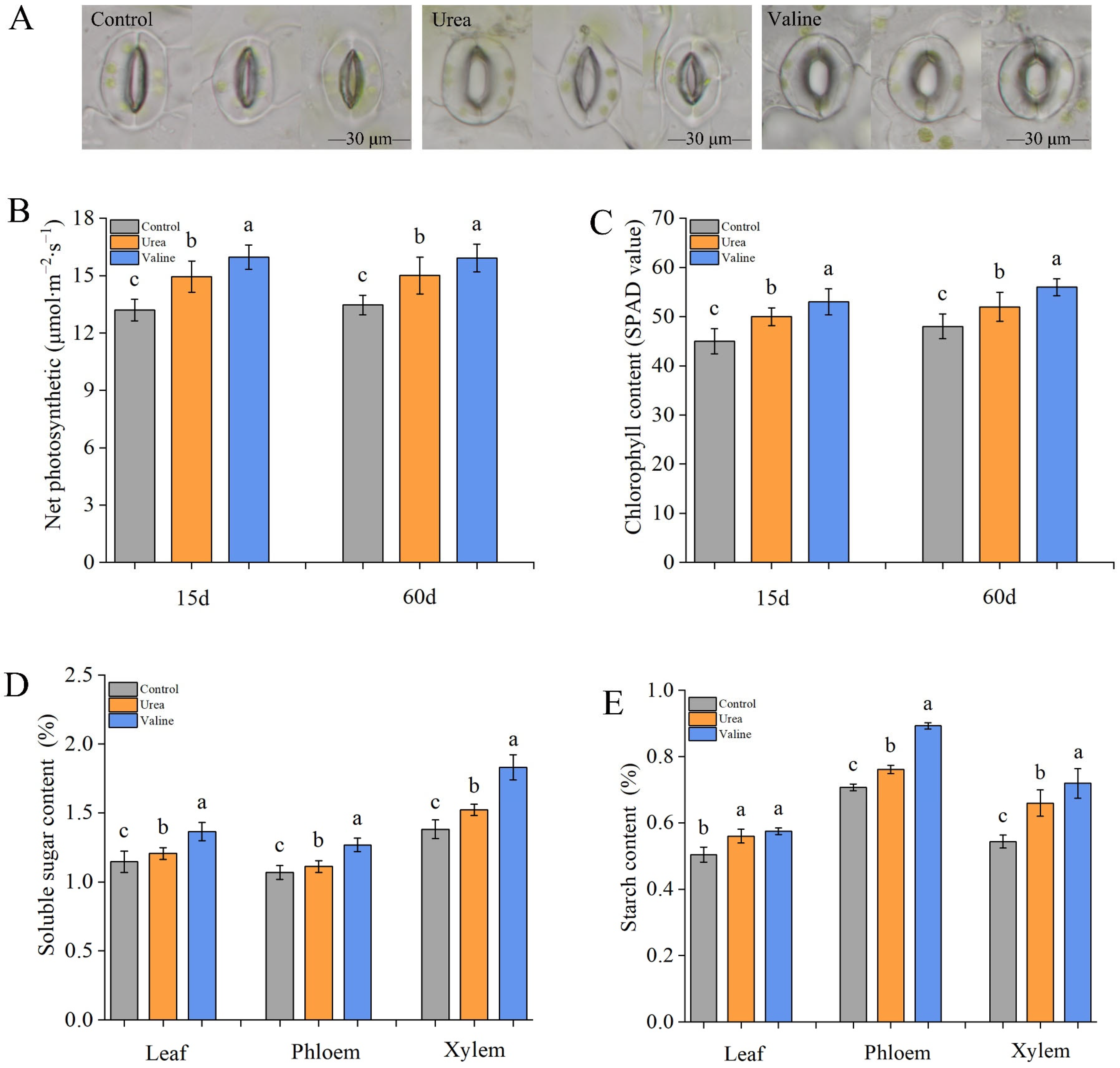
2.4. Effects of Valine and Urea on Carbon Metabolites in Peach Tree
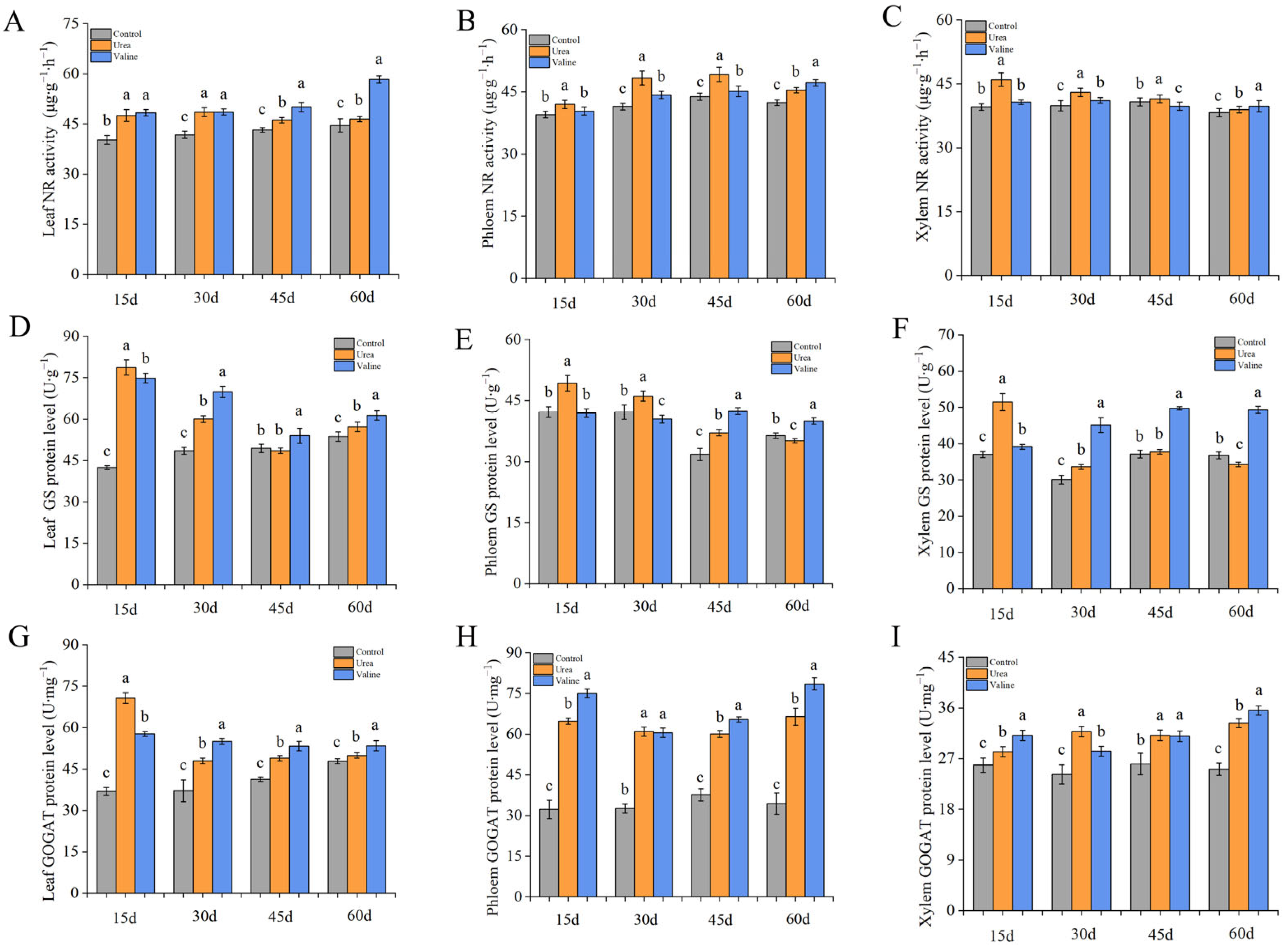
2.5. Effects of Valine and Urea on Enzymes of Nitrogen Metabolism in Peach Tree
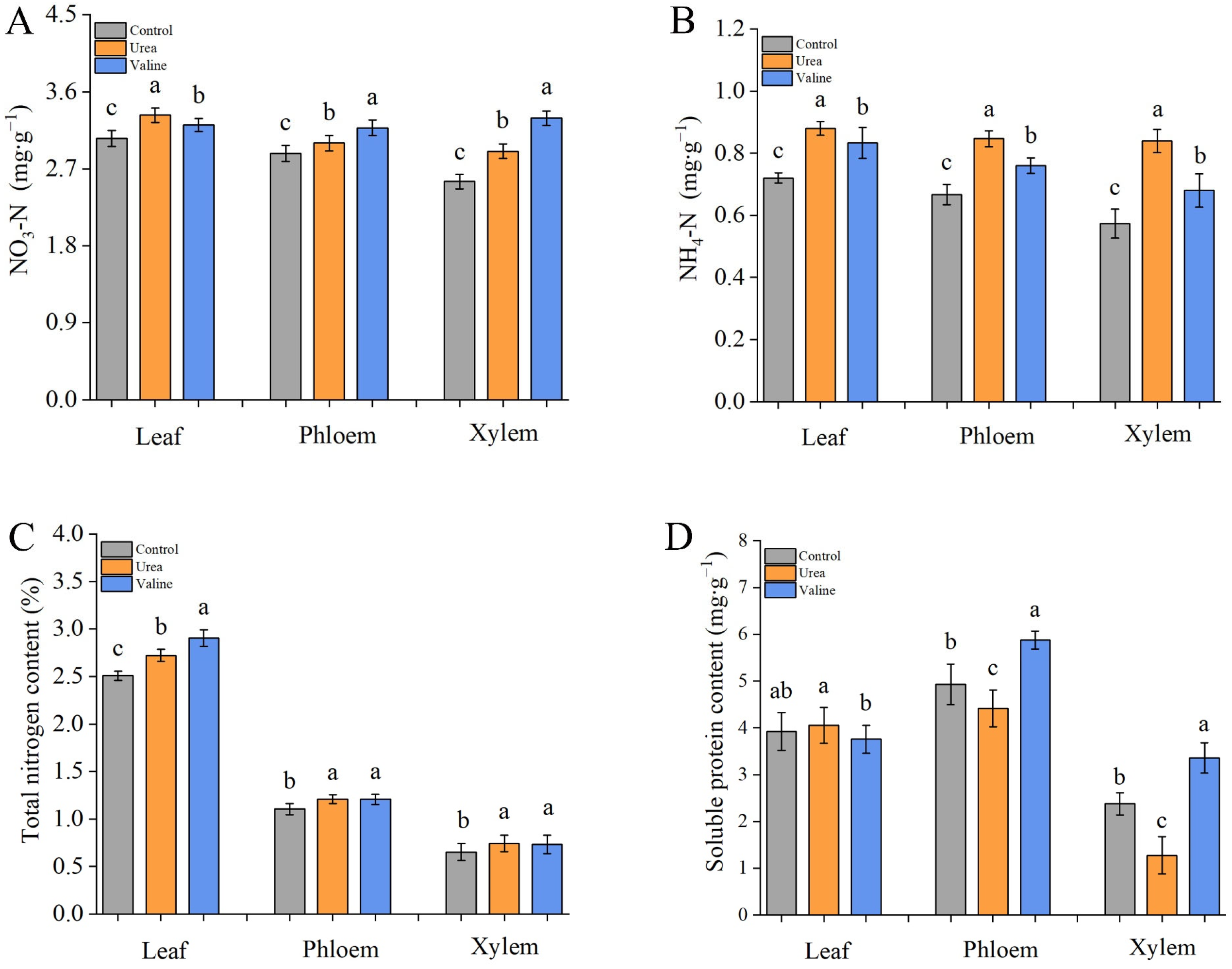
2.6. Effects of Valine and Urea on Nitrogen Metabolites in Peach Tree
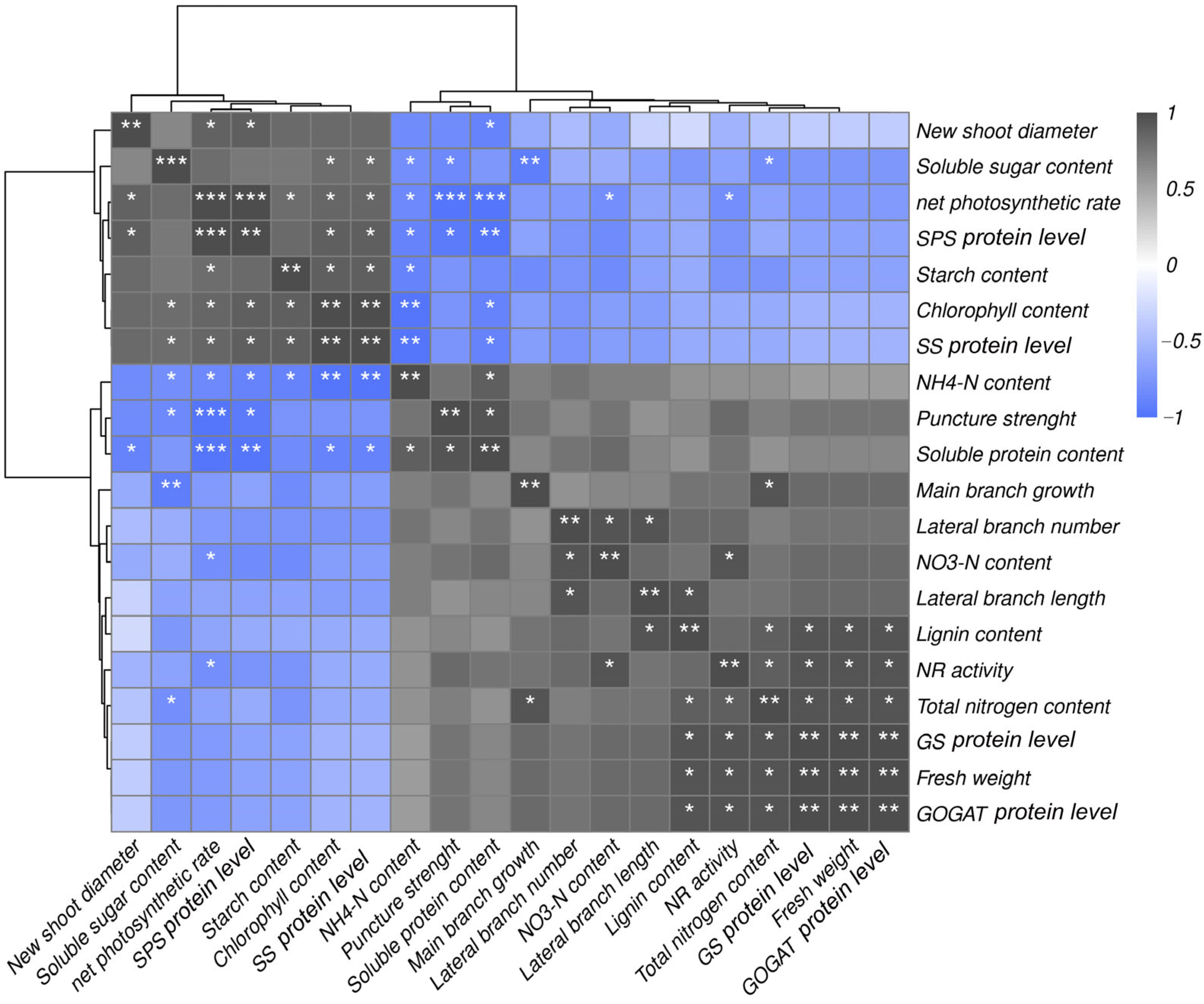
2.7. Correlation Effects of Different Treatments on Key Enzymes, Carbon and Nitrogen Metabolism and Accumulation
3. Discussion
4. Materials and Methods
4.1. Experimental Materials and Experimental Design
4.2. Determination of Shoot Length, Thickness, and Puncture Strength
4.3. Leaf Photosynthetic Rate Determination, Chlorophyll Content, and Stomata
4.4. Leaf Photosynthetic Rate Determination, Chlorophyll Content, and Stomata
4.5. Soluble Sugar, Starch, Soluble Protein, Nitrate Nitrogen, and Ammonium Nitrogen Determinations
4.6. Determination of Enzyme Activity Related to Carbon and Nitrogen Metabolism
4.7. Membership Function Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jin, M.; Mao, S.; Peng, L. Analysis and countermeasures of China’s peach industry export trade. Jiangsu Agric. Sci. 2019, 47, 334–338. [Google Scholar]
- Engindeniz, S.; Cukur, F.; Engindeniz, D.Y. Alternative opportunities for small farms: A case study on technical and economic analysis of peach growing in Turkey. J. Agric. Food Inf. 2003, 5, 47–58. [Google Scholar] [CrossRef]
- Witte, C.P. Urea metabolism in plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Zhang, R.H.; Shu, H.R.; Li, W.W.; Huang, H.C. Study on the nitrogen nutrition of apple tree: The variation of nitrogen nutrition within the plant in a year’s cycle. Acta Hortic. Sin. 1981, 8, 21–28. [Google Scholar]
- Beatty, P.H.; Klein, M.S.; Fischer, J.J.; Lewis, I.A.; Muench, D.G.; Good, A.G. Understanding plant nitrogen metabolism through metabolomics and computational approaches. Plants 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Lv, X.; Yang, J.; Chen, B.; Zhao, W.; Meng, Y.; Oosterhuis, D.M. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crops Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Ning, Y.; Ai, X.Z.; Li, Q.M.; Bi, H.G. Effects of light quality on carbon-nitrogen metabolism, growth, and quality of Chinese chives. J. Appl. Ecol. 2019, 30, 251–258. [Google Scholar]
- Sun, Z.; Chen, Y.F.; Du, J. Elevated CO2 improves lipid accumulation by increasing carbon metabolism in Chlorella sorokiniana. Plant Biotechnol. J. 2016, 14, 557–566. [Google Scholar] [CrossRef]
- Hu, W.; Coomer, T.D.; Loka, D.A.; Oosterhuis, D.M.; Zhou, Z. Potassium deficiency affects the carbon-nitrogen balance in cotton leaves. Plant Physiol. Biochem. 2017, 115, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Tung, S.A.; Huang, Y.; Ali, S.; Hafeez, A.; Shah, A.N.; Song, X.; Ma, X.; Luo, D.; Yang, G. Mepiquat chloride application does not favor leaf photosynthesis and carbohydrate metabolism as well as lint yield in late-planted cotton at high plant density. Field Crops Res. 2018, 221, 108–118. [Google Scholar] [CrossRef]
- Kishorekumar, R.; Bulle, M.; Wany, A.; Gupta, K.J. An overview of important enzymes involved in nitrogen assimilation of plants. Nitrogen Metab. Plants 2022, 2057, 1–13. [Google Scholar] [CrossRef]
- McCarthy, J.L.; Islam, A. Lignin Chemistry, Technology, and Utilization: A Brief History; ACS Publications: Washington, DC, USA, 2000. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.; Hott, C.; Tworkoski, T. Shade effects on growth, flowering and fruit of apple. J. Appl. Hortic. 2015, 17, 101–105. [Google Scholar] [CrossRef]
- Pan, R.Z. Pay attention to the residual toxicity of plant growth regulators. Bull. Biol. 2002, 4, 4–7. [Google Scholar]
- Jie, W.; Hang, Y.; Hua, R.; Qiang, D.; Sheng, X. Analysis of Prohexadione Calcium Residues in Rice by HPLC. Agrochemicals 2015, 54, 528–529. [Google Scholar]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino acid transporters in plant cells: A brief review. Plants 2020, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Meng, X.; Feng, C.; Ran, W.; Yu, G.; Zhang, Y.; Shen, Q. Hydrolytic amino acids employed as a novel organic nitrogen source for the preparation of PGPF-containing bio-organic fertilizer for plant growth promotion and characterization of substance transformation during BOF production. PLoS ONE 2016, 11, e0149447. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Qi, J.; Cai, Y.; Su, N.; Ma, G.; Zhao, X.; Zhang, X. Effects of organic nutrient fertilizer on growth of continuous cropping tomato and soil fertility improvement. China Veg. 2017, 7, 49–53. [Google Scholar]
- He, X.; Zhang, J.; Ren, Y.; Sun, C.; Deng, X.; Qian, M.; Xia, Y. Polyaspartate and liquid amino acid fertilizer are appropriate alternatives for promoting the phytoextraction of cadmium and lead in Solanum nigrum L. Chemosphere 2019, 237, 124483. [Google Scholar] [CrossRef]
- Wang, D.; Deng, X.; Wang, B.; Zhang, N.; Zhu, C.; Jiao, Z.; Li, R.; Shen, Q. Effects of foliar application of amino acid liquid fertilizers, with or without Bacillus amyloliquefaciens SQR9, on cowpea yield and leaf microbiota. PLoS ONE 2019, 14, e0222048. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Ren, G.Z.; Ynag, L.J.; Gong, Z.X.; Zhao, J.J.; Huang, H.T.; Wei, Z.Z.; Xu, Z.C. Effects of different small molecular organics on physiological characteristics, carbon and nitrogen metabolism and quality of flue-cured tobacco. Plant Physiol. J. 2017, 53, 1225–1233. [Google Scholar]
- Cao, X.; Wu, L.; Chen, X.; Han, K. Effects of partial replacement of NO3–N with amino acid on the yield, quality and root secretion of pakchoi (Brassica chinensis L.). Plant Nutr. Fertil. Sci. 2012, 18, 699–705. [Google Scholar]
- Li, S.; Peng, F.; Xiao, Y.; Gong, Q.; Bao, Z.; Li, Y.; Wu, X. Mechanisms of high concentration valine-mediated inhibition of peach tree shoot growth. Front. Plant Sci. 2020, 11, 603067. [Google Scholar] [CrossRef]
- Donaldson, L. Seasonal changes in lignin distribution during tracheid development in Pinus radiata D. Don. Wood Sci. Technol. 1991, 25, 15–24. [Google Scholar] [CrossRef]
- Donaldson, L. Lignin distribution during latewood formation in Pinus radiata D. Don. IAWA J. 1992, 13, 381–387. [Google Scholar] [CrossRef]
- Li, K.Y.; Xing, Y.; Huai, F. Effects of foliar application of urea on sugar content in leaves and fruits for ‘Red Globe’ grape by delay cultivation in greenhouse. China Fruits 2018, 3, 42–46. [Google Scholar]
- Jia, Y.; Yang, L.; Zhou, D.T.; Qu, S.J.; Wang, J.G.; Liu, H.L.; Wang, J.; Zhao, H.W. Effects of Exogenous Materials on Nitrogen Photosynthetic Efficiency and Yield Formation of japonica Rice in Cold Region Under Cold Water Stress at Booting Stage. Chin. J. Rice Sci. 2020, 34, 443–456. [Google Scholar]
- Kang, X.; Chang, C.; Sun, X.; Zhang, X.; Xie, Y.; Ma, F.; Zou, Y. How nitrogen and drought stress affect growth and nitrogen use efficiency for Fuji and Qinguan apple seedlings. Plant Nutr. Fert. Sci. 2014, 4, 965–973. [Google Scholar]
- Guo, L.W. Coupled Effects of Urea Fertilization Techniques and Water Retention Agent on Carbon and Nitrogen Metabolism and Water Use of Maize. Ph.D. Dissertation, Shandong Agricultural University, Tai’an, China, 2014. [Google Scholar]
- Hu, X.H.; Zhou, J.C.; Chen, L.X.; Wang, Q.H. Effects of combined application of ammonium and amino acid nitrogen on growth characteristics and carbon metabolism of sugar beet. Plant Res. 2015, 35, 370–377. [Google Scholar]
- Cui, Z.; Wang, K.; Qiu, J.; Zhu, M.; Zhang, P. Effect of spraying micronutrient and amino acids on growth and quality of Lonicera japonica Thunb. Acta Bot. Boreali-Occident. Sin. 2014, 34, 523–529. [Google Scholar]
- Zeng, X.M. The Influence of Nitrogen Application on Enzyme Activity and Its Output in Fluecured Tobacco Carbon and Nitrogen Metabolism Pathway. Ph.D. Dissertation, Sichuan Agricultural University, Chengdu, China, 2017. [Google Scholar]
- Hu, W.; Zhao, W.; Yang, J.; Oosterhuis, D.M.; Loka, D.A.; Zhou, Z. Relationship between potassium fertilization and nitrogen metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll during the boll development stage. Plant Physiol. Biochem. 2016, 101, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.Z.; Yu, Z.W. Nitrogen metabolism in flag leaf and grain of wheat in response to irrigation regimes. J. Plant Nutr. Soil Sci. 2006, 169, 118–126. [Google Scholar] [CrossRef]
- Zhang, W.Z. Effect of N Management on C and N Metoblisms and Yield of Soybean. Ph.D. Dissertation, Northeast Agricultural University, Ha’erbin, China, 2008. [Google Scholar]
- Jiao, Y.; Chen, Y.; Ma, C.; Qin, J.; Nguyen TH, N.; Liu, D.; Gan, H.; Ding, S.; Luo, Z.B. Phenylalanine as a nitrogen source induces root growth and nitrogen-use efficiency in Populus × canescens. Tree Physiol. 2018, 38, 66–82. [Google Scholar] [CrossRef] [Green Version]
- Trinh, C.S.; Lee, H.; Lee, W.J.; Lee, S.J.; Chung, N.; Han, J.; Lee, H. Evaluation of the plant growth-promoting activity of Pseudomonas nitroreducens in Arabidopsis thaliana and Lactuca sativa. Plant Cell Rep. 2018, 37, 873–885. [Google Scholar] [CrossRef]
- Zhao, S.; Cang, J. Experimental Guidance of Plant Physiology; China Agriculture Press: Beijing, China, 2016. [Google Scholar]
- Jia, X.J.; Dong, L.H.; Ding, C.B.; Li, X.; Yuan, M. Effects of drought stress on reactive oxygen species and their scavenging systems in Chlorophytum capense var. medio-pictum leaf. Acta Prataculturae Sin. 2013, 22, 248. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prunus persica (L.) Batsch. Stomatal Measurements | |||
|---|---|---|---|
| Length (μm) | Width (μm) | Area (μm2) | |
| Control | 27.4 ± 3.2 a | 16.4 ± 1.1 c | 319.1 ± 5.42 c |
| Urea | 26.8 ± 2.4 b | 18.3 ± 1.2 b | 343.5 ± 6.71 b |
| Valine | 26.3 ± 2.7 b | 21.8 ± 1.5 a | 415.3 ± 6.24 a |
| Treatments | Time | SS | SPS | NR | GS | GOGAT | Average |
|---|---|---|---|---|---|---|---|
| Leaf | Val | 0.63 | 1.00 | 1.00 | 1.00 | 0.70 | 0.87 |
| Urea | 0.45 | 0.00 | 0.51 | 0.77 | 0.67 | 0.48 | |
| Control | 0.00 | 0.32 | 0.00 | 0.00 | 0.00 | 0.06 | |
| Phloem | Val | 1.00 | 1.00 | 0.33 | 0.81 | 1.00 | 0.83 |
| Urea | 0.60 | 0.00 | 1.00 | 1.00 | 0.81 | 0.68 | |
| Control | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Xylem | Val | 1.00 | 1.00 | 0.26 | 1.00 | 0.82 | 0.82 |
| Urea | 0.03 | 0.06 | 1.00 | 0.38 | 0.83 | 0.46 | |
| Control | 0.00 | 0.15 | 0.00 | 0.00 | 0.00 | 0.03 |
| Treatments | Soluble Sugar | Starch | NO3-N | NH4-N | Soluble Protein | Average |
|---|---|---|---|---|---|---|
| Val | 0.71 | 0.21 | 0.81 | 0.52 | 1.00 | 0.65 |
| Urea | 0.20 | 0.32 | 1.00 | 1.00 | 0.00 | 0.50 |
| Control | 0.00 | 0.00 | 0.00 | 0.00 | 0.94 | 0.19 |
| Treatments | Concentration (g∙L−1) | Soluble Sugar Content (%) | Starch Content (%) | Total Nitrogen Content (%) | Lignin Content (mg∙g−1) |
|---|---|---|---|---|---|
| Val 1 g∙L−1 | 1 | 0.84 ± 0.09 e | 0.45 ± 0.05 d | 1.55 ± 0.09 e | 473.2 ± 39.7 c |
| Val 2 g∙L−1 | 2 | 1.54 ± 0.12 c | 0.66 ± 0.07 c | 1.96 ± 0.15 d | 507.4 ± 29.5 b |
| Val 4 g∙L−1 | 4 | 1.78 ± 0.11 a | 0.85 ± 0.09 a | 2.93 ± 0.16 a | 532.7 ± 57.3 a |
| Val 8 g∙L−1 | 8 | 1.65 ± 0.07 b | 0.76± 0.05 b | 2.56 ± 0.13 b | 470.5 ± 52.5 c |
| Val 16 g∙L−1 | 16 | 1.32 ± 0.07 d | 0.43 ± 0.04 d | 2.12 ± 0.17 c | 455.3 ± 47.6 d |
| Name | Manufacturer | Model |
|---|---|---|
| Hard tissue slicer | Shanghai Leica Instrument Co., Ltd., Shanghai, China | HistoCore |
| Tissue Spreader | KEDEE, Guangzhou, China | Category 1 |
| Upright Optical Microscope | Nikon Japan, Tokyo, Japan | Nikon Eclipse E100 |
| imaging system | Nikon Japan, Tokyo, Japan | Nikon DS-U3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Li, S.; Yu, H.; Gong, Q.; Zhang, B.; Liu, G.; Xiao, Y.; Peng, F. Effects of Valine and Urea on Carbon and Nitrogen Accumulation and Lignin Content in Peach Trees. Plants 2023, 12, 1596. https://doi.org/10.3390/plants12081596
Sun M, Li S, Yu H, Gong Q, Zhang B, Liu G, Xiao Y, Peng F. Effects of Valine and Urea on Carbon and Nitrogen Accumulation and Lignin Content in Peach Trees. Plants. 2023; 12(8):1596. https://doi.org/10.3390/plants12081596
Chicago/Turabian StyleSun, Maoxiang, Suhong Li, Haixiang Yu, Qingtao Gong, Binbin Zhang, Guangyuan Liu, Yuansong Xiao, and Futian Peng. 2023. "Effects of Valine and Urea on Carbon and Nitrogen Accumulation and Lignin Content in Peach Trees" Plants 12, no. 8: 1596. https://doi.org/10.3390/plants12081596