
Antitumor and Antioxidant Activities of In Vitro Cultivated and Wild-Growing Clinopodium vulgare L. Plants
 , , , ,
, , , , 

Abstract
:1. Introduction
2. Results
2.1. In Vitro Culture
2.1.1. Shoot Induction and Multiplication
2.1.2. In Vitro Rooting and Acclimation
2.2. Phytochemical Analyses
2.2.1. Total Polyphenol and Total Flavonoid Contents
2.2.2. Antioxidant Activity
2.2.3. HPLC Analysis
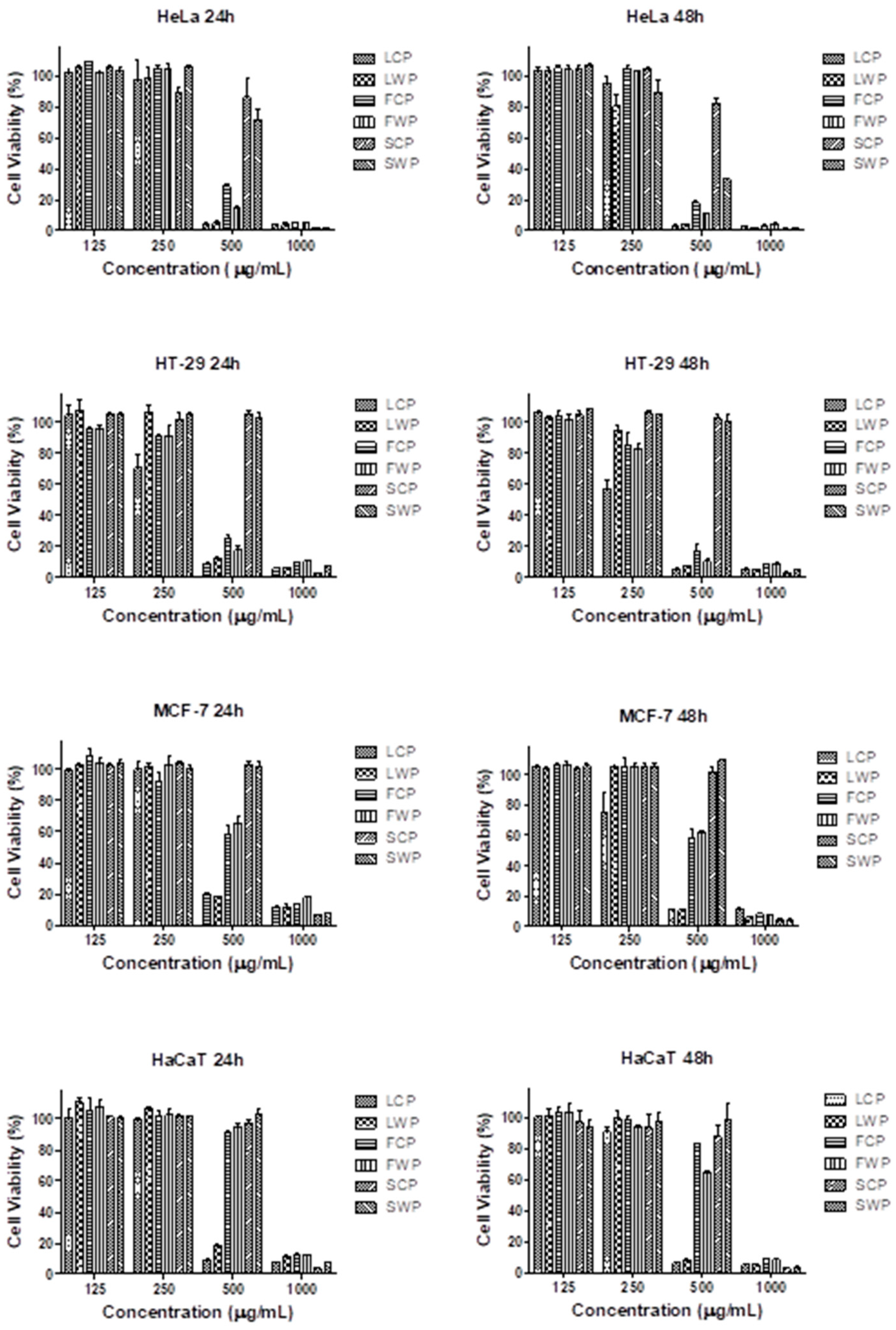
2.3. Antitumor Activity
2.3.1. MTT Cell Viability Assay
2.3.2. Fluorescence Microscopy
3. Discussion
4. Materials and Methods
4.1. In Vitro Culture
4.1.1. Initial Plant Material
4.1.2. Seed Sterilization
4.1.3. Media Composition for In Vitro Cultivation
4.1.4. In Vitro Rooting and Acclimatization of Obtained Plants
4.1.5. Conditions for In Vitro Cultures
4.2. Phytochemical Analysis
4.2.1. Plant Material
4.2.2. Preparation of Freeze-Dried Extracts
- FCP—flowers of in vitro cultivated plants;
- LCP—leaves of in vitro cultivated plants;
- SCP—stem of in vitro cultivated plants;
- FWP—flowers of wild plants;
- LWP—leaves of wild plants;
- SWP—stems of wild plants.
4.2.3. Total Polyphenol and Total Flavonoid Contents
4.2.4. Antioxidant Activity Assays
4.2.5. HPLC Determination of Phenolic and Flavonoid Compounds
4.3. Antitumor Activity
4.3.1. Cell Lines
4.3.2. Test Sample Preparation
4.3.3. Cell Viability Assay
4.3.4. Fluorescence Microscopy Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batsalova, T.; Bardarov, K.; Bardarov, V.; Moten, D.; Dzhambazov, B. Cytotoxic properties of Clinopodium vulgare L. extracts on selected human cell lines. Comptes Rendus L’acad. Bulg. Sci. 2017, 70, 645–650. [Google Scholar]
- Nassar-Eddin, G.; Zheleva-Dimitrova, D.; Danchev, N.; Vitanska-Simeonova, R. Antioxidant and enzyme-inhibiting activity of lyophilized extract from Clinopodium vulgare L. (Lamiaceae). Pharmacia 2021, 68, 259–263. [Google Scholar] [CrossRef]
- Medicinal Plant Act. Decree no. 29 of Ministry of Environment and Water, Sofia, Bulgaria accepted by the National Assembly on 23 March 2000; No 29/07.04.2000; State Gazette: Dyersburg, TN, USA, 2002; pp. 9–21. (In Bulgarian) [Google Scholar]
- Carović-Stanko, K.; Petek, M.; Grdiša, M.; Pintar, J.; Bedeković, D.; Ćustić, M.H.; Satovic, Z. Medicinal plants of the family Lamiaceae as functional foods—A review. Czech J. Food Sci. 2016, 34, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Miyase, T.; Matsushima, Y. Saikosaponin homologues from Clinopodium spp. The structures of clinoposaponins XII-XX. Chem. Pharm. Bull. 1997, 45, 1493–1497. [Google Scholar] [CrossRef] [Green Version]
- Obreshkova, D.; Tashkov, W.; Ilieva, I. Phenolcarboxylic acids in Clinopodium vulgare L. Comptes Rendus L’acad. Bulg. Sci. 2001, 54, 54–57. [Google Scholar]
- Tepe, B.; Sihoglu-Tepe, A.; Daferera, D.; Polissiou, M.; Sokmen, A. Chemical composition and antioxidant activity of the essential oil of Clinopodium vulgare L. Food Chem. 2007, 103, 766–770. [Google Scholar] [CrossRef]
- Bardarov, K.; Todorova, T.; Miteva, D.; Bardarov, V.; Atanassov, A.; Chankova, S. Preliminary screening for study of the chemical composition of Clinopodium vulgare L. water extract. Comptes Rendus L’acad. Bulg. Sci. 2016, 69, 717–725. [Google Scholar]
- Burk, D.R.; Senechal-Willis, P.; Lopez, L.C.; Hogue, B.G.; Daskalova, S.M. Suppression of lipopolysaccharide-induced inflammatory responses in RAW 264.7 murine macrophages by aqueous extract of Clinopodium vulgare L. (Lamiaceae). J. Ethnopharmacol. 2009, 126, 397–405. [Google Scholar] [CrossRef]
- Amirova, K.M.; Dimitrova, P.; Marchev, A.S.; Aneva, I.Y.; Georgiev, M.I. Clinopodium vulgare L. (wild basil) extract and its active constituents modulate cyclooxygenase-2 expression in neutrophils. Food Chem. Toxicol. 2019, 124, 1–9. [Google Scholar] [CrossRef]
- Canter, P.H.; Thomas, H.; Ernst, E. Bringing medicinal plants into cultivation: Opportunities and challenges for biotechnology. Trends Biotechnol. 2005, 23, 180–185. [Google Scholar] [CrossRef]
- Anand, S. Various approaches for secondary metabolite production through plant tissue culture. Pharmacia 2010, 1, 1–7. [Google Scholar]
- Diaz, M.S.; Palacio, L.; Figueroa, A.C.; Goleniowski, M.E. In vitro propagation of muna-muna (Clinopodium odorum (Griseb.) Harley). Biotechnol. Res. Int. 2012, 2012, 196583. [Google Scholar] [CrossRef] [Green Version]
- Engelmann-Sylvestre, I.; Engelmann, F. Cryopreservation of in vitro-grown shoot tips of Clinopodium odorum using aluminium cryo-plates. In Vitro Cell. Dev. Biol. Plant 2015, 51, 185–191. [Google Scholar] [CrossRef]
- Vázquez, A.M.; Goleniowski, M.E.; Aimar, M.L.; Diaz, M.S.; Demmel, G.I.; Decarlini, M.F.; Cantero, J.J. Profile Characterization of Volatile Organic Compounds on in vitro Propagated Plants of Clinopodium odorum and it’s Comparison with the Wild Plant. SAJ Pharm. Pharmacol. 2018, 5, 103. [Google Scholar]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. Studies on seed germination and micropropagation of Clinopodium nepeta: A medicinal and aromatic plant. HortScience 2019, 54, 1558–1564. [Google Scholar] [CrossRef]
- Mehandzhiyski, A.; Batovska, D.; Dimitrov, D.; Evstatieva, L.; Danova, K. Nitric oxide-scavenging activity of in vitro cultured Balkan medicinal and aromatic plants. Bulg. J. Agric. Sci. 2013, 19, 31–34. [Google Scholar]
- Dzhambazov, B.; Daskalova, S.; Monteva, A.; Popov, N. In vitro screening for antitumour activity of Clinopodium vulgare L. (Lamiaceae) extracts. Biol. Pharm. Bull. 2002, 25, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gerdakaneh, M.; Badakhshan, H.; Mohamadi, M.; Arji, I. Effect of different media and growth regulators on micropropagation of GF677. Plant Prod. 2020, 43, 241–254. [Google Scholar]
- Gaspar, T.H.; Kevers, C.; Faivre-Rampant, O.; Crevecoeur, M.; Penel, C.; Greppin, H.; Dommes, J. Changing concepts in plant hormone action. In Vitro Cell. Dev. Biol. Plant 2003, 39, 85–106. [Google Scholar] [CrossRef]
- Malik, S.K.; Chaudhury, R.; Kalia, R.K. Rapid in vitro multiplication and conservation of Garcinia indica: A tropical medicinal tree species. Sci. Hortic. 2005, 106, 539–553. [Google Scholar] [CrossRef]
- Lodha, D.; Patel, A.K.; Rai, M.K.; Shekhawat, N.S. In vitro plantlet regeneration and assessment of alkaloid contents from callus cultures of Ephedra foliata (Unth phog), a source of anti-asthmatic drugs. Acta Physiol. Plant. 2014, 36, 3071–3079. [Google Scholar] [CrossRef]
- Nordström, A.; Tarkowski, P.; Tarkowska, D.; Norbaek, R.; Åstot, C.; Dolezal, K.; Sandberg, G. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance for auxin–cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 2004, 101, 8039–8044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Máthé, Á.; Hassan, F.; Abdul Kader, A. In Vitro Micropropagation of Medicinal and Aromatic Plants. In Medicinal and Aromatic Plants of the World; Máthé, Á., Ed.; Springer: Dordrecht, The Netherlands, 2015; Volume 1, pp. 305–336. [Google Scholar]
- Sharma, S.; Shahzad, A.; Mahmood, S.; Saeed, T. High-frequency clonal propagation, encapsulation of nodal segments for short-term storage and germplasm exchange of Ficus carica L. Trees 2015, 29, 345–353. [Google Scholar] [CrossRef]
- Lakshmanan, P.; Lee, C.L.; Goh, C.J. An efficient in vitro method for mass propagation of a woody ornamental Ixora coccinea L. Plant Cell Rep. 1997, 16, 572–577. [Google Scholar] [CrossRef]
- Pazos-Navarro, M.; Del Río, J.A.; Ortuño, A.; Romero-Espinar, P.; Correal, E.; Dabauza, M. Micropropagation from apical and nodal segments of Bituminaria bituminosa and the furanocoumarin content of propagated plants. J. Hortic. Sci. Biotechnol. 2012, 87, 29–35. [Google Scholar] [CrossRef]
- Zayova, E.; Stancheva, I.; Geneva, M.; Petrova, M.; Dimitrova, L. Antioxidant activity of in vitro propagated Stevia rebaudiana Bertoni plants of different origins. Turk. J. Biol. 2013, 37, 106–113. [Google Scholar] [CrossRef]
- Saeedpour, A.; Godehkahriz, S.J.; Lohrasebi, T.; Esfahani, K.; Salmanian, A.H.; Razavi, K. The effect of endogenous hormones, total antioxidant and total phenol changes on regeneration of barley cultivars. Iran. J. Biotechnol. 2021, 19, e2838. [Google Scholar]
- Shiwani, K.; Sharma, D.; Kumar, A. Improvement of Plant Survival and Expediting Acclimatization Process. In Commercial Scale Tissue Culture for Horticulture and Plantation Crops; Gupta, S., Chaturvedi, P., Eds.; Springer: Singapore, 2022; pp. 277–291. [Google Scholar]
- Ruhland, C.T.; Xiong, F.S.; Clark, W.D.; Day, T.A. The Influence of Ultraviolet-B Radiation on Growth, Hydroxycinnamic Acids and Flavonoids of Deschampsia antarctica during Springtime Ozone Depletion in Antarctica. Photochem. Photobiol. 2005, 81, 1086–1093. [Google Scholar] [CrossRef]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef]
- Oberoi, H.S.; Sandhu, S. KTherapeutic and Nutraceutical Potential of Bioactive Compounds Extracted from Fruit Residues AU—Babbar, Neha. Crit. Rev. Food Sci. Nutr. 2015, 55, 319–337. [Google Scholar]
- Spiridon, I.; Bodirlau, R.; Teaca, C.A. Total phenolic content and antioxidant activity of plants used in traditional Romanian herbal medicine. Cent. Eur. J. Biol. 2011, 6, 388–396. [Google Scholar] [CrossRef]
- Amarowicz, R.; Pegg, R.B.; Rahimi-Moghaddam, P.; Barl, B.; Weil, J.A. Free-radical scavenging capacity and antioxidant activity of selected plant species from the Canadian prairies. Food Chem. 2004, 84, 551–562. [Google Scholar] [CrossRef]
- Kratchanova, M.; Denev, P.; Ciz, M.; Lojek, A.; Mihailov, A. Evaluation of antioxidant activity of medicinal plants containing polyphenol compounds. Comparison of two extraction systems. Acta Biochim. Pol. 2010, 57, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Stefanovic, O.; Stankovic, M.S.; Comic, L. In vitro antibacterial efficacy of Clinopodium vulgare L. extracts and their synergistic interaction with antibiotics. J. Med. Plant Res. 2011, 5, 4074–4079. [Google Scholar]
- Balkan, B.; Balkan, S.; Aydoğdu, H.; Güler, N.; Ersoy, H.; Aşkın, B. Evaluation of antioxidant activities and antifungal activity of different plants species against pink mold rot-causing Trichothecium roseum. Arab. J. Sci. Eng. 2017, 42, 2279–2289. [Google Scholar] [CrossRef]
- Bektašević, M.; Politeo, O.; Roje, M.; Jurin, M. Polyphenol Composition, Anticholinesterase and Antioxidant Potential of the Extracts of Clinopodium vulgare L. Chem. Biodiver. 2022, 19, e202101002. [Google Scholar] [CrossRef]
- Vladimir-Knežević, S.; Blažeković, B.; Kindl, M.; Vladić, J.; Lower-Nedza, A.D.; Brantner, A.H. Acetylcholinesterase inhibitory, antioxidant and phytochemical properties of selected medicinal plants of the Lamiaceae family. Molecules 2014, 19, 767–782. [Google Scholar] [CrossRef] [Green Version]
- Sarikurkcu, C.; Ozer, M.S.; Tepe, B.; Dilek, E.; Ceylan, O. Phenolic composition, antioxidant and enzyme inhibitory activities of acetone, methanol and water extracts of Clinopodium vulgare L. subsp. vulgare L. Ind. Crop. Prod. 2015, 76, 961–966. [Google Scholar] [CrossRef]
- Nunes, R.; Pasko, P.; Tyszka-Czochara, M.; Szewczyk, A.; Szlosarczyk, M.; Carvalho, I.S. Antibacterial, antioxidant and anti-proliferative properties and zinc content of five south Portugal herbs. Pharm. Biol. 2017, 55, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarino, M.; Sandin-Espana, P.; Melgarejo, P.; De Cal, A. High chlorogenic and neochlorogenic acid levels in immature peaches reduce Monilinia laxa infection by interfering with fungal melanin biosynthesis. J. Agric. Food Chem. 2011, 59, 3205–3213. [Google Scholar] [CrossRef] [PubMed]
- Kurita, S.; Kashiwagi, T.; Ebisu, T.; Shimamura, T.; Ukeda, H. Identification of neochlorogenic acid as the predominant antioxidant in Polygonum cuspidatum leaves. Ital. J. Food Sci. 2016, 28, 25–31. [Google Scholar]
- Park, S.Y.; Jin, M.L.; Yi, E.H.; Kim, Y.; Park, G. Neochlorogenic acid inhibits against LPS-activated inflammatory responses through up-regulation of Nrf2/HO-1 and involving AMPK pathway. Environ. Toxicol. Pharmacol. 2018, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mirza, F.; Lorenzo, J.; Drissi, H.; Lee, F.Y.; Soung, D.Y. Dried plum alleviates symptoms of inflammatory arthritis in TNF transgenic mice. J. Nutr. Biochem. 2018, 52, 54–61. [Google Scholar] [CrossRef]
- Kozyra, M.; Komsta, Ł.; Wojtanowski, K. Analysis of phenolic compounds and antioxidant activity of methanolic extracts from inflorescences of Carduus sp. Phytochem. Lett. 2019, 31, 256–262. [Google Scholar] [CrossRef]
- Gao, X.H.; Zhang, S.D.; Wang, L.T.; Yu, L.; Zhao, X.L.; Ni, H.Y.; Wang, Y.Q.; Wang, J.D.; Shan, C.H.; Fu, Y.J. Anti-inflammatory effects of neochlorogenic acid extract from mulberry leaf (Morus alba L.) against LPS-stimulated inflammatory response through mediating the AMPK/Nrf2 signaling pathway in A549 cells. Molecules 2020, 25, 1385. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Orcajada, S.; Matencio, A.; Vicente-Herrero, C.; García-Carmona, F.; López-Nicolás, J.M. Study of the fluorescence and interaction between cyclodextrins and neochlorogenic acid, in comparison with chlorogenic acid. Sci. Rep. 2021, 11, 3275. [Google Scholar] [CrossRef]
- Fang, W.; Ma, Y.; Wang, J.; Yang, X.; Gu, Y.; Li, Y. In vitro and in vivo antitumor activity of neochlorogenic acid in human gastric carcinoma cells are complemented with ROS generation, loss of mitochondrial membrane potential and apoptosis induction. J. BUON 2019, 24, 221–226. [Google Scholar]
- Sitarek, P.; Kowalczyk, T.; Santangelo, S.; Białas, A.J.; Toma, M.; Wieczfinska, J.; Sliwinski, T.; Skała, E. The extract of Leonurus sibiricus transgenic roots with AtPAP1 transcriptional factor induces apoptosis via DNA damage and down regulation of selected epigenetic factors in human cancer cells. Neurochem. Res. 2018, 43, 1363–1370. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.Y.; Sang, L.X.; Jiang, M. Catechins and their therapeutic benefits to inflammatory bowel disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef] [Green Version]
- Li, X.X.; Liu, C.; Dong, S.L.; Ou, C.S.; Lu, J.L.; Ye, J.H.; Yue-Rong, L.; Zheng, X.Q. Anticarcinogenic potentials of tea catechins. Front. Nutr. 2022, 9, 3024. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Denev, P.; Ciz, M.; Ambrozova, G.; Lojek, A.; Yanakieva, I.; Kratchanova, M. Solid-phase extraction of berries’ anthocyanins and evaluation of their antioxidative properties. Food Chem. 2010, 123, 1055–1061. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Medium | Shoot Tips | Nodal Segments | ||||
|---|---|---|---|---|---|---|
| Shoot Formation (%) | Mean Number of Shoots Per Explant | Mean Height of Shoots (cm) | Shoot Formation (%) | Mean Number of Shoots Per Explants | Mean Height of Shoots (cm) | |
| MS | 10 | 1.15 ± 0.36 a | 7.55 ± 1.24 e | 10 | 1.2 ± 0.41 a | 8.3 ± 1.54 d |
| B1 | 60 | 2.05 ± 0.68 cd | 3.85 ± 1.00 c | 90 | 4.6 ± 1.69 e | 4.78 ± 0.70 b |
| iP1 | 15 | 1.35 ± 0.48 ab | 3.95 ± 1.02 c | 30 | 2.15 ± 0.67 b | 3.45 ± 1.05 a |
| Z1 | 40 | 1.95 ± 0.60 cd | 3.1 ± 0.80 ab | 70 | 3.4 ± 1.09 cd | 5.35 ± 1.59 bc |
| K1 | 30 | 1.75 ± 0.55 bc | 5.85 ± 1.14 d | 80 | 2.65 ± 0.81 bc | 6.0 ± 1.12 c |
| B1I0.1 | 80 | 3.0 ± 0.72 e | 3.6 ± 1.04 bc | 100 | 6.9 ± 2.57 f | 4.65 ± 0.86 b |
| iP1I0.1 | 20 | 1.4 ± 0.59 ab | 2.9 ± 0.92 a | 100 | 2.6 ± 0.82 bc | 3.7 ± 0.76 a |
| Z1I0.1 | 60 | 2.3 ± 0.57 d | 2.86 ± 0.78 a | 100 | 4.0 ± 1.55 de | 5.23 ± 2.36 bc |
| K1I0.1 | 40 | 1.9 ± 0.64 c | 4.1 ± 0.89 c | 100 | 3.6 ± 1.18 d | 4.8 ± 1.42 b |
| LSD | 0.37 | 0.62 | 0.84 | 0.85 | ||
| Samples | Total Polyphenols, mg GAE/100 g DW | Total Flavonoids, mg QE/100 g DW | ORAC, µmol TE/g DW | HORAC, µmol GAE/g DW |
|---|---|---|---|---|
| FCP | 29,927.6 ± 592.1 d | 763.2 ± 37.2 a | 7281.3 ± 82.9 c | 2251.0 ± 21.2 c |
| LCP | 28,016.9 ± 447.3 c | 1100.9 ± 81.5 b | 7342.5 ± 54.4 c | 2326.9 ± 56.0 d |
| SCP | 16,733.7 ± 390.0 a | 824.6 ± 109.1 a | 3393.1 ± 40.2 a | 1278.8 ± 32.4 b |
| FWP | 27,292.8 ± 85.3 b | 1460.5 ± 63.1 c | 7246.3 ± 62.4 c | 2534.0 ± 45.8 e |
| LWP | 30,510.9 ± 85.3 d | 1815.8 ± 91.2 d | 7258.2 ± 105.9 c | 2572.7 ± 41.8 e |
| SWP | 16,713.6 ± 142.2 a | 1614.0 ± 154.3 c | 3849.9 ± 74.0 b | 1179.4 ± 18.3 a |
| LSD | 623.72 | 171.92 | 129.98 | 68.18 |
| Samples | Neochlorogenic Acid | Chlorogenic Acid | Catechin | Rosmarinic Acid | Quercetin | Apigenin | Caffeic Acid | Sum |
|---|---|---|---|---|---|---|---|---|
| FCP | 790.6 ± 1.0 f | 52.7 ± 0.4 a | 222.9 ± 5.3 b | 369.3 ± 0.3 c | - | - | 65.9 ± 0.9 c | 1501.4 |
| LCP | 595.6 ± 1.7 e | 183.0 ± 0.8 d | 136.6 ± 6.1 a | 1286.6 ± 1.8 e | - | - | 23.0 ± 0.6 a | 2224.8 |
| SCP | 244.8 ± 0.2 b | 175.0 ± 1.3 c | 114.8 ± 0.8 b | 565.8 ± 5.6 b | 125.3 ± 0.2 b | 23.0 ± 0.9 a | 1248.7 | |
| FWP | 526.8 ± 0.2 c | 81.1 ± 1.6 b | - | 851.8 ± 0.2 d | - | - | 69.3 ± 0.9 d | 1529.0 |
| LWP | 575.0 ± 0.1 d | 205.6 ± 0.3 e | - | 1734.6 ± 4.2 f | - | - | 31.1 ± 0.4 b | 2546.2 |
| SCP | 193.0 ± 1.3 a | 259.5 ± 0.2 f | - | 87.5 ± 1.6 a | 319.7 ± 0.5 a | 39.3 ± 0.3 a | 31.1 ± 0.1 b | 930.1 |
| LSD | 1.73 | 1.65 | 12.95 | 3.57 | 9.01 | 0.57 | 1.25 |
| IC50 | HeLa | HeLa | HT-29 | HT-29 | MCF-7 | MCF-7 | HaCaT | HaCaT |
|---|---|---|---|---|---|---|---|---|
| (µg/mL) | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h |
| LCP | 361.6 | 343.9 | 304.4 | 266.5 | 429.0 | 319.1 | 400.0 | 345.1 |
| LWP | 380.8 | 309.0 | 426.8 | 363.0 | 427.8 | 426.3 | 427.6 | 398.9 |
| FCP | 479.6 | 467.8 | 398.9 | 362.0 | 557.0 | 537.8 | 729.1 | 659.6 |
| FWP | 464.2 | 457.9 | 380.9 | 336.5 | 620.3 | 556.9 | 753.9 | 557.0 |
| SCP | 621.2 | 597.1 | 882.5 | 880.1 | 933.7 | 924.7 | 721.7 | 639.7 |
| SWP | 560.0 | 423.6 | 934.1 | 928.8 | 938.2 | 924.7 | 906.4 | 742.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, M.; Dimitrova, L.; Dimitrova, M.; Denev, P.; Teneva, D.; Georgieva, A.; Petkova-Kirova, P.; Lazarova, M.; Tasheva, K. Antitumor and Antioxidant Activities of In Vitro Cultivated and Wild-Growing Clinopodium vulgare L. Plants. Plants 2023, 12, 1591. https://doi.org/10.3390/plants12081591
Petrova M, Dimitrova L, Dimitrova M, Denev P, Teneva D, Georgieva A, Petkova-Kirova P, Lazarova M, Tasheva K. Antitumor and Antioxidant Activities of In Vitro Cultivated and Wild-Growing Clinopodium vulgare L. Plants. Plants. 2023; 12(8):1591. https://doi.org/10.3390/plants12081591
Chicago/Turabian StylePetrova, Maria, Lyudmila Dimitrova, Margarita Dimitrova, Petko Denev, Desislava Teneva, Ani Georgieva, Polina Petkova-Kirova, Maria Lazarova, and Krasimira Tasheva. 2023. "Antitumor and Antioxidant Activities of In Vitro Cultivated and Wild-Growing Clinopodium vulgare L. Plants" Plants 12, no. 8: 1591. https://doi.org/10.3390/plants12081591