Production of Secondary Metabolites from Cell Cultures of Sageretia thea (Osbeck) M.C. Johnst. Using Balloon-Type Bubble Bioreactors

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
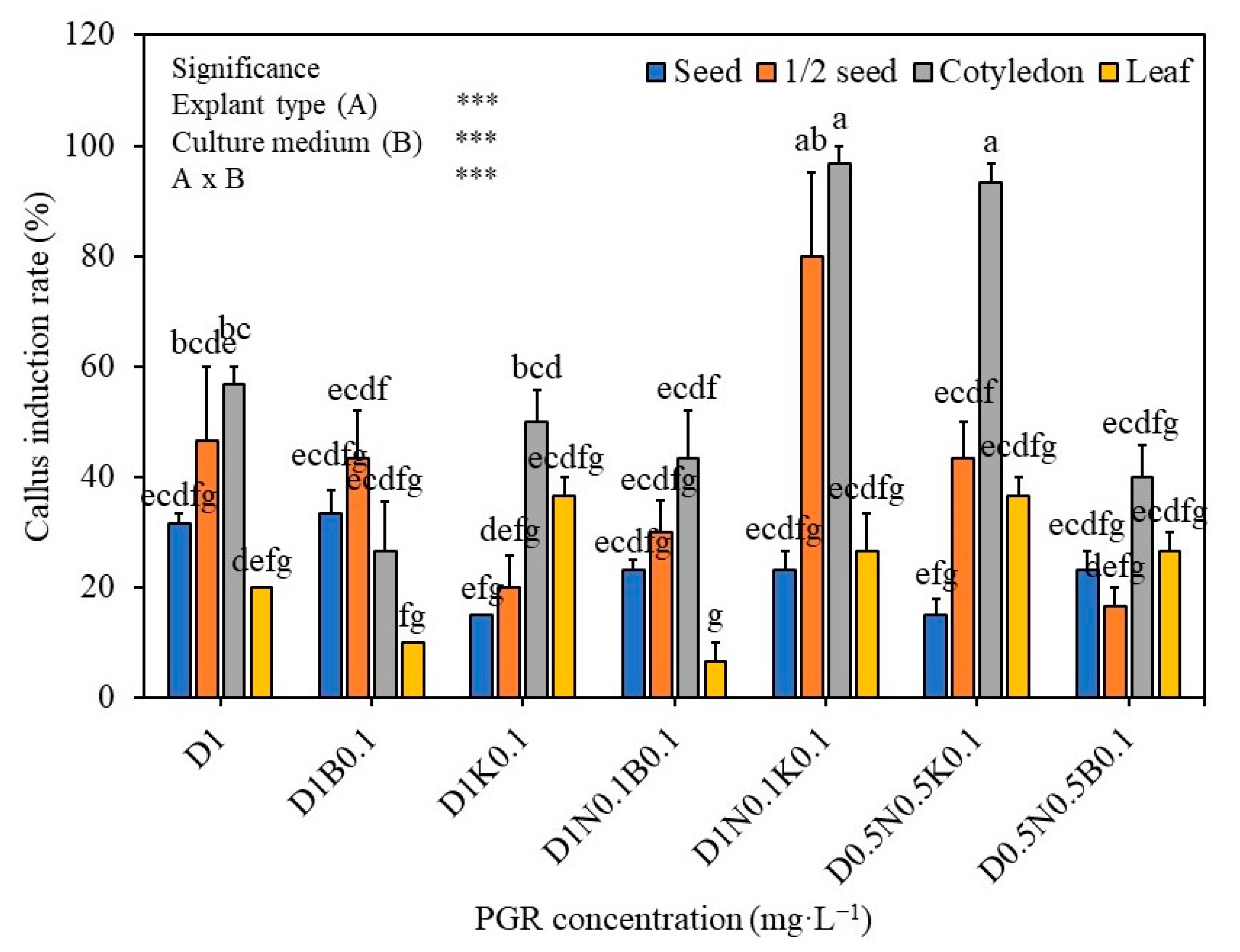
2.1. The Effect of Growth Regulators and Explant Type on the Callus Induction
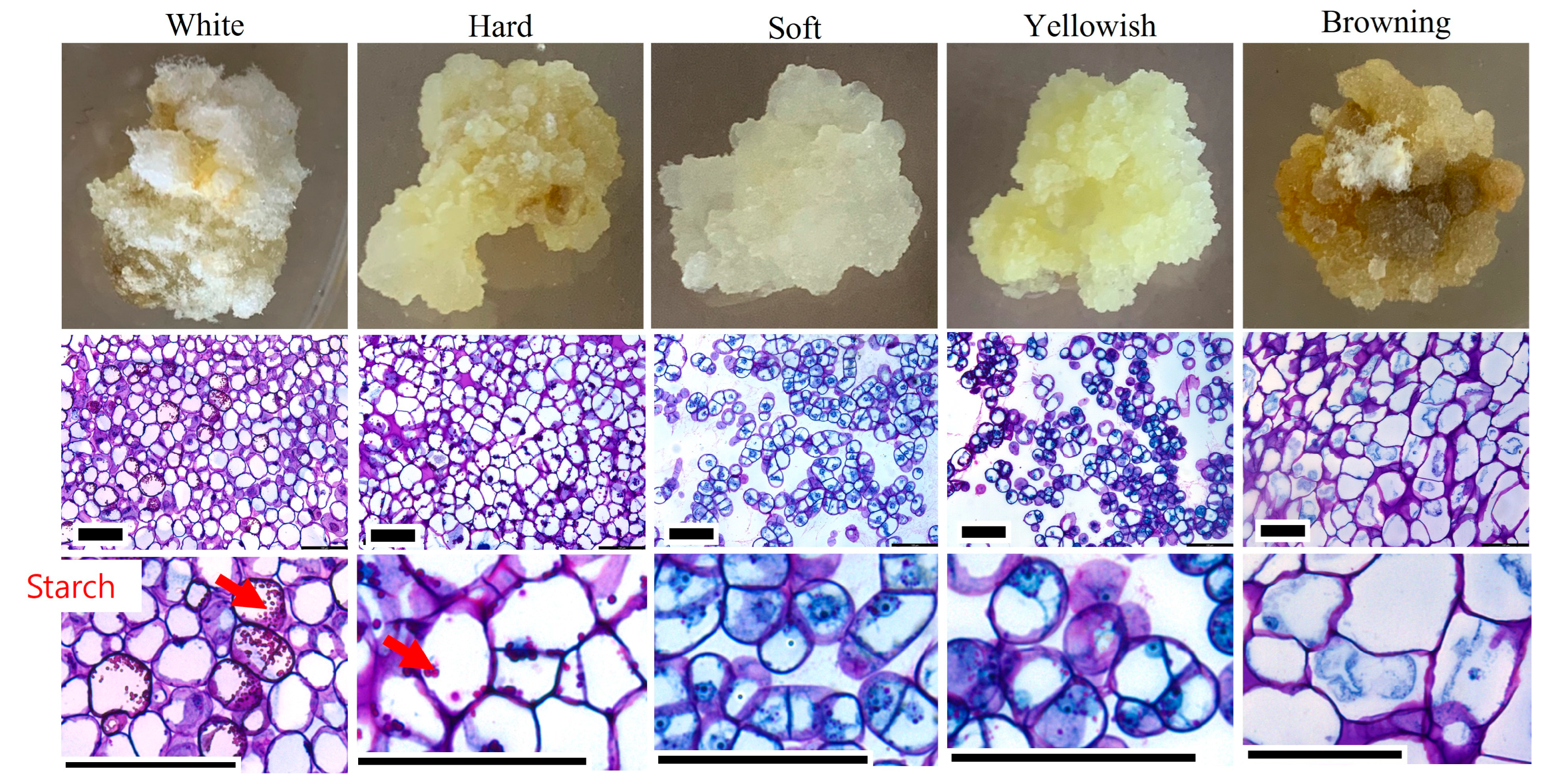
2.2. Selection of Friable Callus
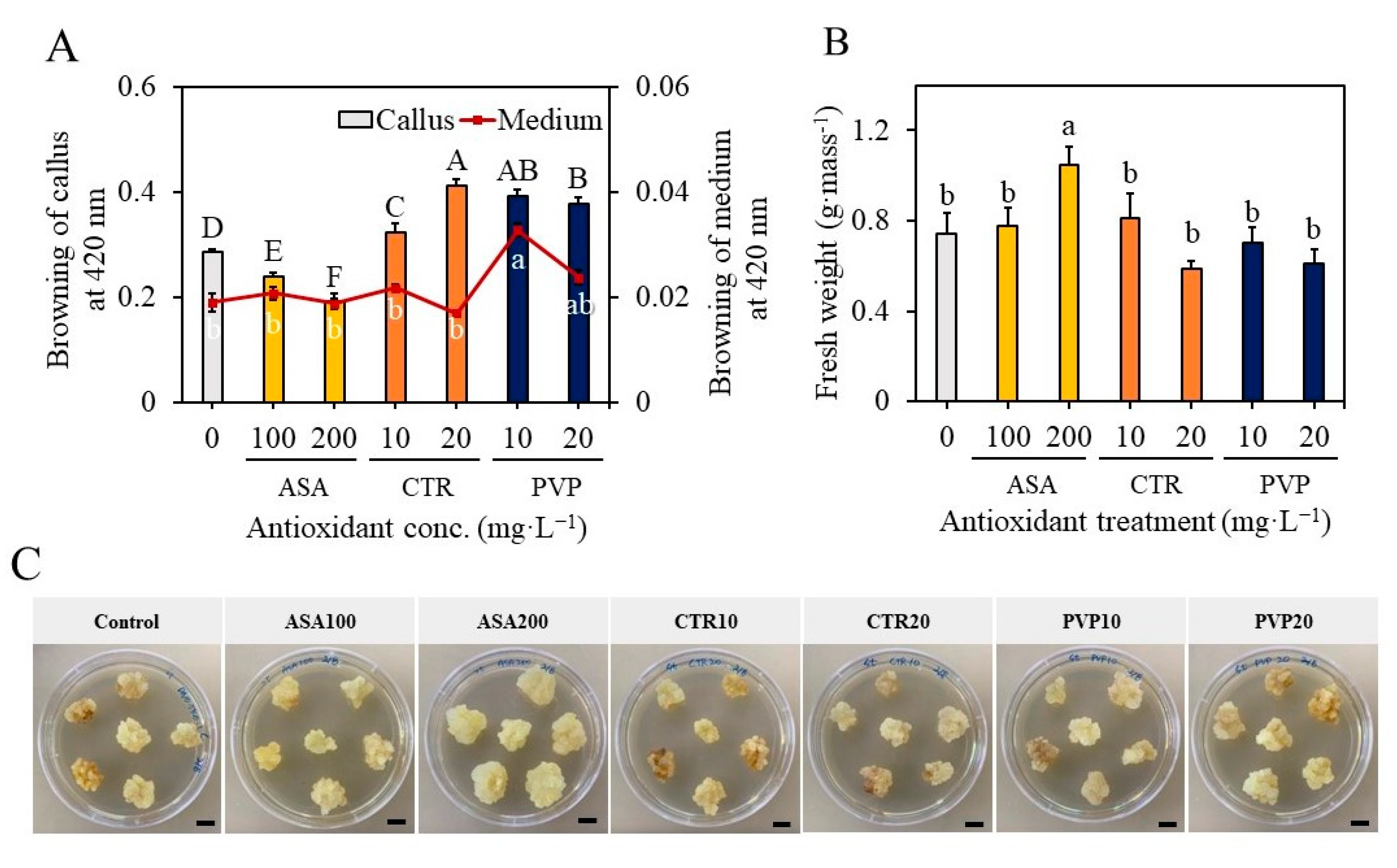
2.3. Effect of Antioxidants on Callus Proliferation and Overcoming the Problems of Browning of the Medium
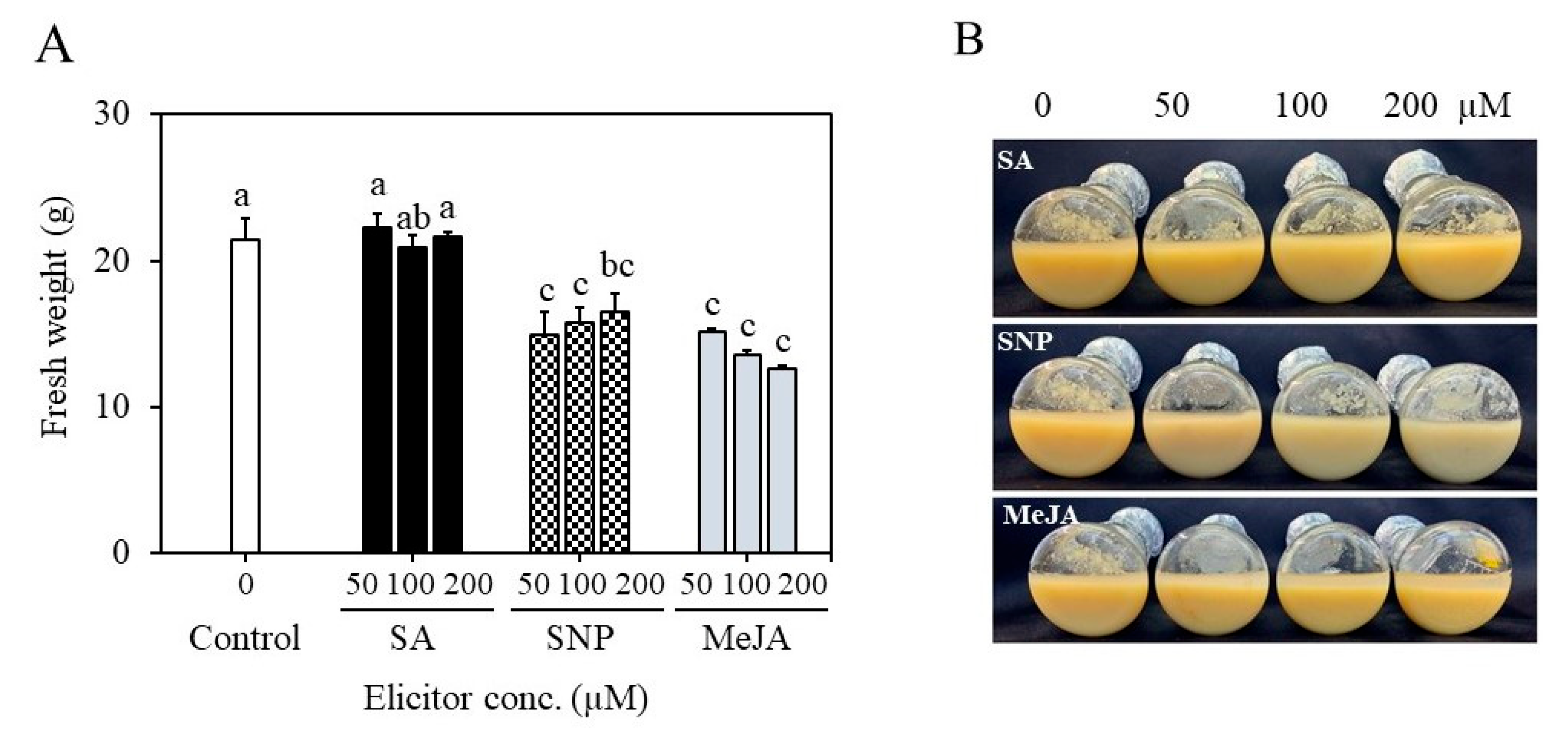
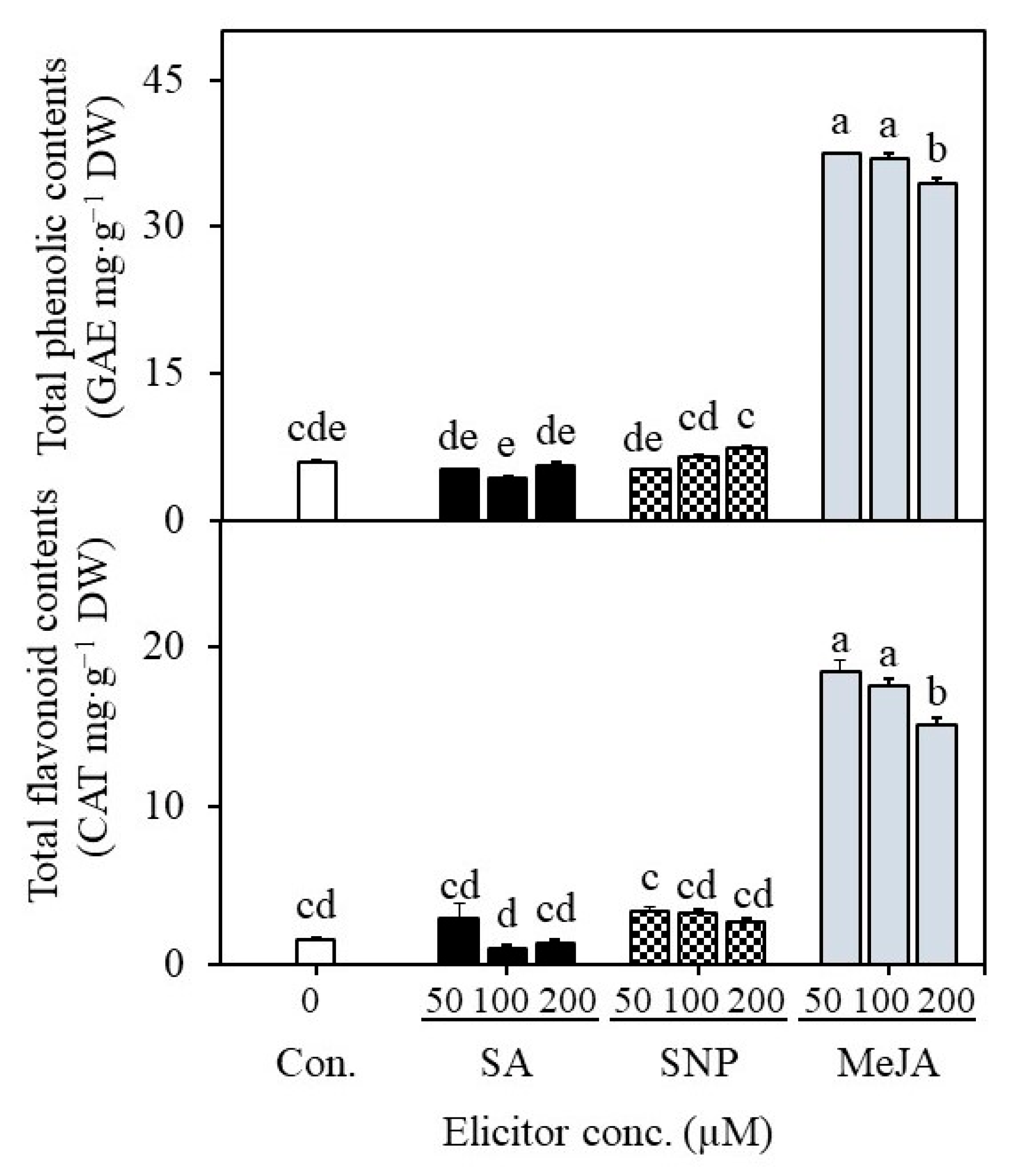
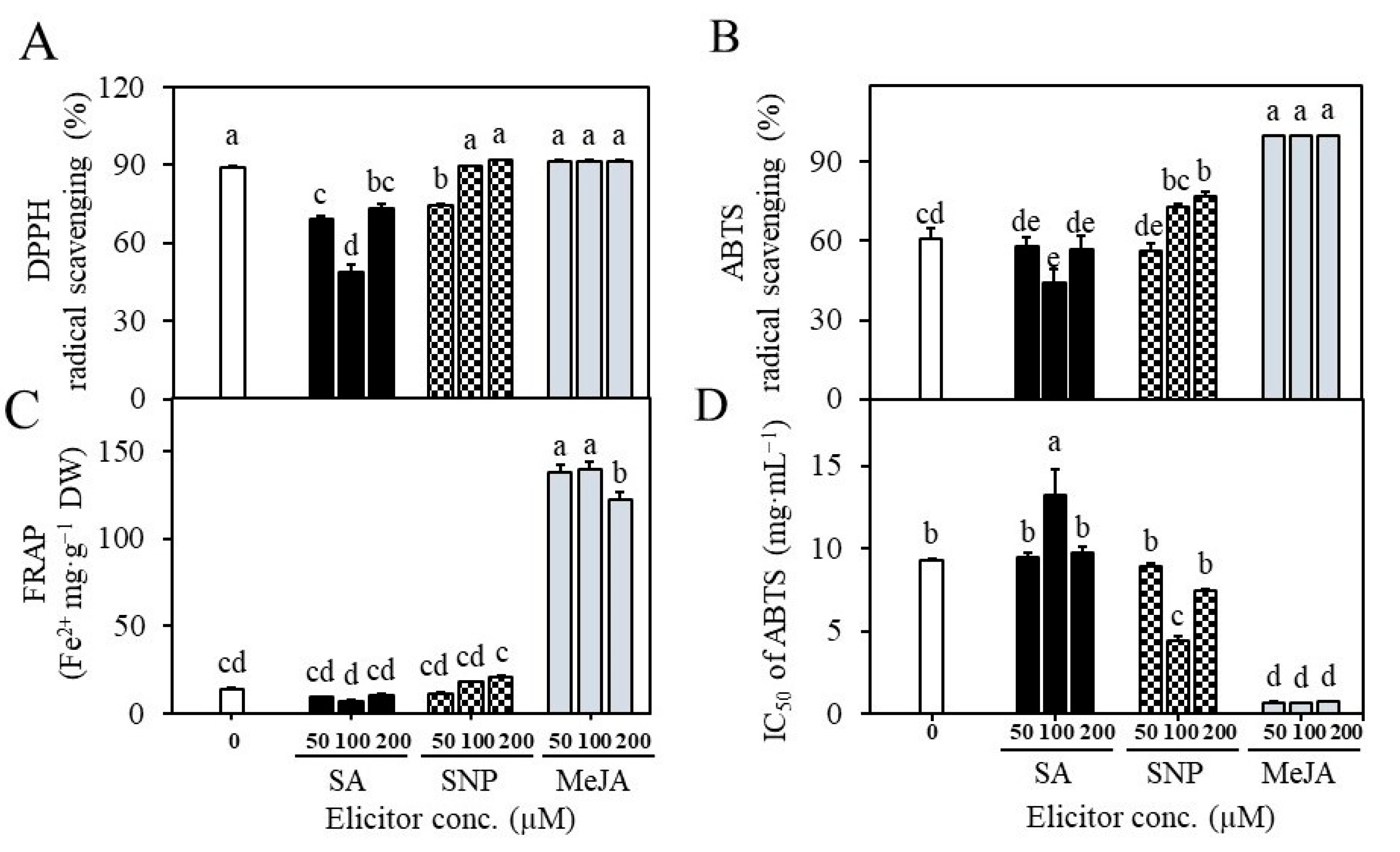
2.4. Establishment of Suspension Cultures in Erlenmeyer Flasks and Elicitation
2.5. Establishment of Bioreactor Cultures for the Production of Biomass and Phenolics
3. Discussion
4. Materials and Methods
4.1. Plant Material and Seed Germination
4.2. Chemicals and Reagents
4.3. Callus Induction
4.4. Histological Analysis of Callus
4.5. Raising of Callus Cultures with Supplementation of Antioxidants
4.6. Establishment of Cell Suspension Culture in Erlenmeyer’s Flasks and Elicitation of Cultures
4.7. Establishment of Cell Suspension Cultures in Balloon-Type Bubble Bioreactors
4.8. Preparation of Cell Extracts
4.9. Estimation of Total Phenolic Content
4.10. Estimation of Total Flavonoid Content
4.11. Analysis of Antioxidant Activities
4.11.1. 2,2 Diphenyl 1 picrylhydrazyl (DPPH) Radical Scavenging Assay
4.11.2. 2,2′-Azino-bis (3-ethybenzothiazoline-6-sulphonic Acid (ABTS) Assay
4.11.3. Ferric Reducing Antioxidant Power (FRAP) Assay
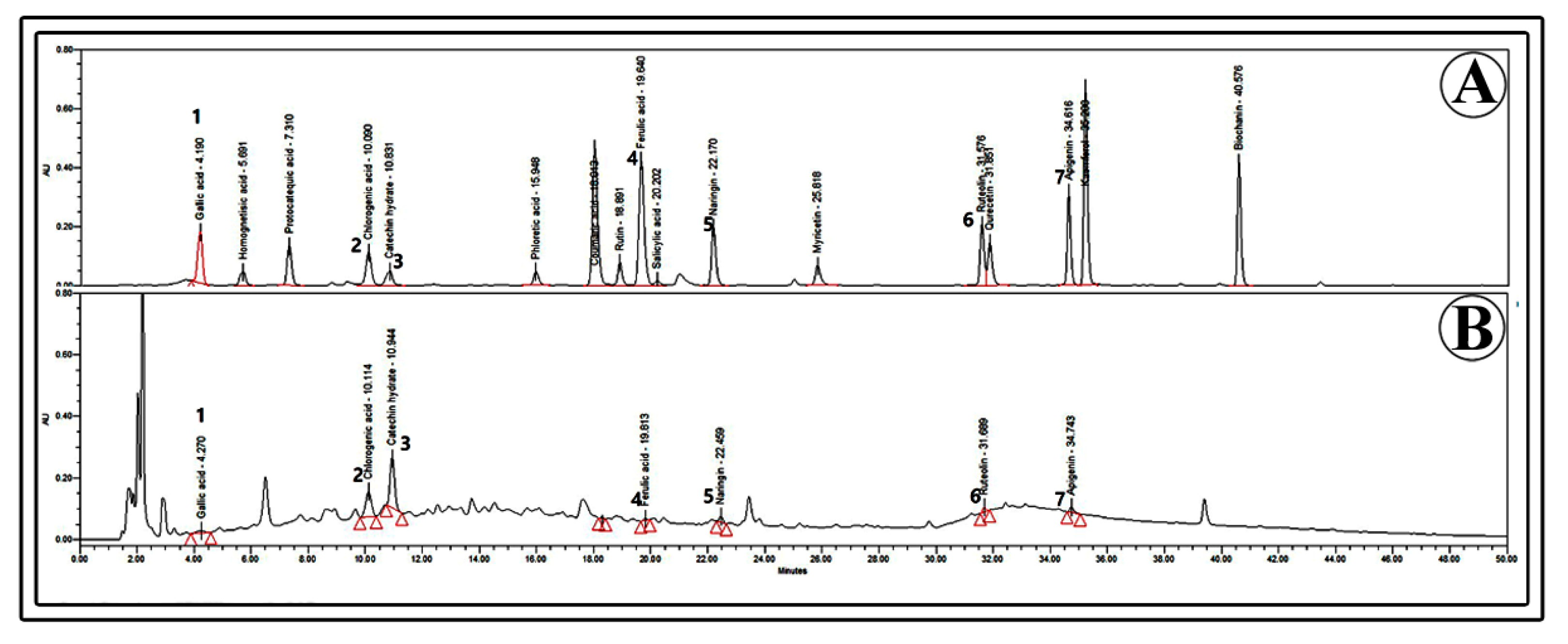
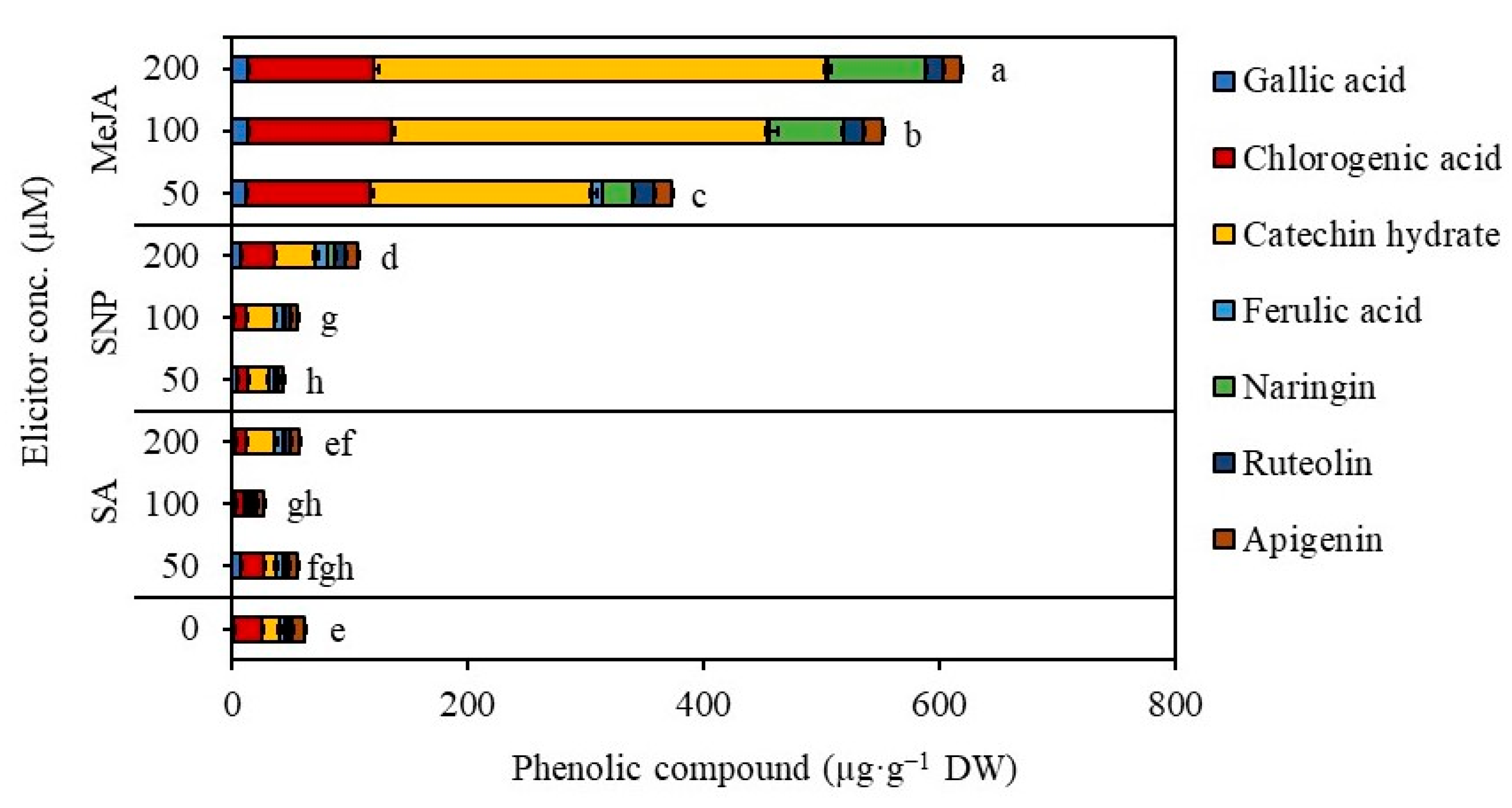
4.12. Quantification of Phenolic Compounds Using High-Performance Liquid Chromatography (HPLC)
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO (Plants of the World Online). Facilitated by the Royal Botanic Gardens, Kew. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 10 January 2023).
- Hyun, T.K.; Sang, S.C.; Sang, C.K.; Kim, J.S. Nutritional and nutraceutical characteristics of Sageretia theezans fruit. J. Food Drug Anal. 2015, 23, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.K.; Chen, C.Y.O.; Blumberg, J.B. Flavonoid-rich fraction from Sageretia theezans leaves scavenges reactive oxygen radical species and increases the resistance of low-density lipoprotein in oxidation. J. Med. Food 2009, 12, 1310–1315. [Google Scholar] [CrossRef] [PubMed]
- Song, S.C.; Song, C.K.; Kim, J.S. Vegetation and habitat environment of Sageretia thea in Jeju island. Korean J. Med. Crop Sci. 2014, 22, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Song, S.C.; Song, C.K.; Kim, J.S. Characteristics of seed germination and fruit for Segeretia thea in Jeju region. Korean J. Med. Crop Sci. 2015, 23, 8–12. [Google Scholar] [CrossRef]
- Xu, L.; Yang, X.; Li, B. Chemical constituents of Sageretia theezans Brongn. Zhongguo Zhong Yao Za Zhi 1994, 19, 675–676. [Google Scholar]
- Chung, S.K.; Kim, Y.C.; Takay, Y.; Terahima, K.; Niwa, M. Novel flavanol glycoside, 7,O-methyl mearnsitrin, from Sageretia theezans and its antioxidant effect. J. Agric. Food Chem. 2004, 52, 4664–4668. [Google Scholar] [CrossRef]
- Ko, G.A.; Shrestha, S.; Cho, S.K. Sageretia thea fruit extracts rich in methyl linoleate and methyl linolenate downregulate melanogenesis via the Akt/GSK3 β signalling pathway. Nutr. Res. Pract. 2018, 12, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.A.; Son, M.; Cho, S.K. Comparative evaluation of free radical scavenging activities and cytotoxicity of various solvent fractions of Sandoong Sageretia thea (Osbeck) M.C. Johnst. Branches. Food Sci. Bioethanol. 2016, 25, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.N.; Park, H.G.; Park, S.B.; Kim, J.D.; Eo, H.J.; Song, J.H.; Jeong, J.B. Extracts from Sageretia thea reduce cell viability inducing cyclin D1 proteasomal degradation and HO-1 expression in human colorectal cancer cells. BMC Complemen. Altern. Med. 2019, 19, 43. [Google Scholar] [CrossRef]
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Huhn, T.; Eibl, D. Plant cell culture technology in the cosmetics and food industries: Current state and future trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675. [Google Scholar] [CrossRef] [Green Version]
- Krasteva, G.; Georgiev, V.; Pavlov, A. Recent applications of plant cell culture technology in cosmetics and foods. Eng. Life Sci. 2021, 21, 68–76. [Google Scholar] [CrossRef]
- Barbulova, A.; Apone, F.; Colucci, G. Plant cell cultures as source of cosmetic active ingredients. Cosmetics 2014, 1, 94–104. [Google Scholar] [CrossRef]
- Murthy, H.N.; Lee, E.J.; Paek, K.Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dalawai, D.; Bhat, M.A.; Dandin, V.S.; Paek, K.Y.; Park, S.Y. Biotechnological Production of Useful Phytochemicals from Adventitious Root Cultures. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Fundamentals and Applications; Ramawat, K.G., Ekiert, H.M., Goyal, S., Eds.; Springer Nature: Geneva, Switzerland, 2021; pp. 469–486. [Google Scholar]
- Murthy, H.N.; Joseph, K.S.; Paek, K.Y.; Park, S.Y. Anthraquinone production from cell and organ cultures of Rubia species: An overview. Metabolites 2023, 13, 39. [Google Scholar] [CrossRef]
- Thanh, N.T.; Murthy, H.N.; Paek, K.Y. Optimization of ginseng cell culture in airlift bioreactors and developing the large-scale production system. Ind. Crops Prod. 2014, 60, 343–348. [Google Scholar] [CrossRef]
- Murthy, H.N.; Georgiev, M.I.; Kim, Y.S.; Jeong, C.C.; Kim, S.J.; Park, S.Y.; Paek, K.Y. Ginsenosides: Perspective for sustainable biotechnological production. Appl. Microbiol. Biotechnol. 2014, 98, 6243–6295. [Google Scholar] [CrossRef]
- Murthy, H.N.; Kim, Y.S.; Park, S.Y.; Paek, K.Y. Biotechnological production of caffeic acid derivatives from cell and organ cultures of Echinacea species. Appl. Microbiol. Biotechnol. 2014, 98, 7707–7717. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Kim, Y.S.; Georgiev, M.I.; Paek, K.Y. Biotechnological production of eleutherosides: Current state and perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 7319–7329. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Kim, Y.S.; Park, S.Y.; Paek, K.Y. Hypericins: Biotechnological production from cell and organ cultures. Appl. Microbiol. Biotechnol. 2014, 98, 9187–9198. [Google Scholar] [CrossRef]
- Cui, H.Y.; Murthy, H.N.; Moh, S.H.; Cui, Y.; Lee, E.J.; Paek, K.Y. Protocrom culture of Dendrobium candidum in balloon type bubble bioreactors. Biochem. Eng. J. 2014, 88, 26–29. [Google Scholar] [CrossRef]
- Cui, H.Y.; Murthy, H.N.; Moh, S.H.; Cui, Y.Y.; Lee, E.J.; Paek, K.Y. Production of biomass and bioactive compounds in protocorm cultures of Dendrobium candidum Wall ex Lindl. using balloon type bubble bioreactors. Ind. Crops Prod. 2014, 53, 28–33. [Google Scholar] [CrossRef]
- Thungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabi, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Ncube, B.; Finnie, J.F.; Van Staden, J. Quality from the field: The impact of environmental factors as quality determinants in medicinal plants. S. Afr. J. Bot. 2012, 82, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sehgal, H.; Joshi, M. The journey and new breakthroughs of plant growth regulators in tissue cultures. In Advances in Plant Tissue Culture, Current Development and Future Trends; Academic Press: Cambridge, MA, USA, 2022; pp. 85–108. [Google Scholar] [CrossRef]
- Machakova, I.; Zazimolova, E.; George, E.F. Plant growth regulators: Introduction; auxins, their analogues and inhibitors. In Plant Propagation by Tissue Culture, 3rd ed.; George, E.F., Hall, M.A., De Klerk, G.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar] [CrossRef]
- Nurazah, Z.; Radzali, M.; Syahida, A.; Maziah, M. Effects of plant growth regulators on callus induction from Cananga odorata flower petal explant. Afr. J. Biotechnol. 2009, 8, 2740–2743. [Google Scholar]
- Khanpour-Ardesteni, N.; Sharifi, M.; Behmanesh, M. Establishment of callus and cell suspension culture of Scrophularia striata Boiss.: An in vitro approach for acetoside production. Cytobiotechnology 2015, 67, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Yu, L.J.; Hu, Q.Y.; Chen, S.C.; Sun, Y.P. Establishment of callus and cell suspension cultures of Corydalis saxicola Bunting, a rare medicinal plant. Z. Naturforsch. 2006, 61, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Winson, K.W.S.; Chew, B.L.; Sathasivam, K.; Subramaniam, S. The establishment of callus and cell suspension cultures of Hylocerus costaricensis for the production of betalain pigments with antioxidant potential. Ind. Crops Prod. 2020, 155, 112750. [Google Scholar] [CrossRef]
- Legha, M.R.; Prasad, K.V.; Singh, S.K.; Kaur, C.; Arora, A.; Kumar, S. Induction of carotenoid pigments in callus cultures of Calendula officinals L. in response to nitrogen and sucrose levels. In Vitro Cell. Dev. Biol. Plant 2012, 28, 39–45. [Google Scholar] [CrossRef]
- Fahim, J.R.; Hegazi, G.A.E.M.; El-Fadi, R.E.S.A.; Alhady, M.R.A.A.A.; Desoukey, S.Y.; Rmadan, M.A.; Kamel, M.S. Production of rhoifolin and tiliroside from callus cultures of Chorisia chodatii and Chorisia spciosa. Phytochem. Lett. 2015, 13, 218–227. [Google Scholar] [CrossRef]
- Akaneme, F.I.; Eneobong, E.E. Tissue culture of Pinus carbaea Mor. var. hondurensis barr. And golf. II. Effects of two auxins and two cytokinins on callus growth habits and subsequent organogenesis. Afr. J. Biotechnol. 2008, 7, 757–765. [Google Scholar]
- Betekhtin, A.; Rojek, M.; Jaskowiak, J.; Milewaska-Handel, A.; Kwasniewaska, J.; Kostyukova, Y.; Hasterok, R. Nuclear genome stability in long-term cultivated callus lines of Fogopyrum tataricum (L.) Gaetn. PLoS ONE 2017, 12, e0173537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laukkanen, H.; Rautiainen, L.; Taulavuori, E.; Hohtola, A. Changes in cellular structures and enzymatic acuities during browning of Scots pine callus derived from mature buds. Tree Physiol. 2000, 20, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchendu, E.E.; Paliyath, G.; Brown, D.C.; Saxena, P.K. In vitro propagation of North American ginseng (Panax quinquefolius L.). In Vitro Cellular Dev. Biol. Plant 2011, 47, 710–718. [Google Scholar] [CrossRef]
- Tang, W.; Newton, R.J.; Outhavong, V. Exogenously added polyamines recover browning tissue into normal callus cultures and improve plant regeneration in pine. Physiol. Plant. 2004, 124, 386–395. [Google Scholar] [CrossRef]
- Habibi, N.; Sutar, R.K.; Purohit, S.D. Role of PGRs and inhibitors in induction and control of somatic embryogenesis in Themeda quadrivalvis. Indian J. Exp. Biol. 2009, 47, 198–203. [Google Scholar]
- Nagomuo, M.; Mnene, E.; Ndakidemi, P. Control of lethal browning by using ascorbic acid on shoot tip cultures of a local Musa spp. (banana) cv. Mzuzu in Tanzania. Afr. J. Biotechnol. 2014, 13, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Vijayalakshmi, U.; Shourie, A. Remedial effect of ascorbic acid and citric acid on oxidative browning of Glycyrrhiza glabra callus cultures. BioTechnolgia 2016, 97, 179–186. [Google Scholar] [CrossRef]
- Ho, T.T.; Murthy, H.N.; Park, S.Y. Methyl jasmonate induced odixative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [Green Version]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhancement of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [Green Version]
- Thanh, N.T.; Murthy, H.N.; Yu, K.Y.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate elicitation enhanced synthesis of ginsenoside by cell suspension cultures of Panax ginseng in 5-l balloon type bubble bioreactors. Appl. Microbiol. Biotechnol. 2005, 67, 197–201. [Google Scholar] [CrossRef]
- Kim, Y.S.; Hahn, E.J.; Murthy, H.N.; Paek, K.Y. Adventitious root growth and ginsenoside accumulation of Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 2004, 26, 1619–1622. [Google Scholar] [CrossRef]
- Wang, J.W.; Wu, J.Y. Nitric oxide is involved in methyl jasmonate-induced defense responses and secondary metabolism activities in Taxus cells. Plant Cell Physiol. 2005, 46, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zheng, P.; Wang, J.W. Nitric oxide elicitation for secondary metabolite production in cultured plant cell. Appl. Microbiol. Biotechnol. 2012, 93, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, D.; Cuaspud, O.; Arias, P.; Ruiz, O.; Arias, M. Effect of salicylic acid and methyl jasmonate in the production of phenolic compounds in plant cell suspension cultures of Thevetia peruviana. Biotechnol. Rep. 2018, 19, e00273. [Google Scholar] [CrossRef]
- Gadzovaska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Hagege, D.; Courtois, D.; Joseph, C. The influence of salicylic acid elicitation on shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant Cell Tissue Organ Cult. 2013, 113, 25–39. [Google Scholar] [CrossRef]
- Xu, M.J.; Dong, J.F.; Zhu, M.Y. Effect of nitric oxide on catharanthine production and growth of Catharanthus roseus suspension cells. Biotechnol. Bioeng. 2005, 89, 367–372. [Google Scholar] [CrossRef]
- Marchev, A.S.; Yordanov, Z.P.; Georgiev, M.I. Green (cell) factories for advanced production of plant secondary metabolites. Crit. Rev. Biotechnol. 2020, 40, 443–458. [Google Scholar] [CrossRef]
- Chen, G.; Yi, Z.; Chen, X.; Su, W.; Li, X. Polyphenol nonoparticals from commonly consumed tea for scavenging free radicals, stabilizing, pickering, emulsions, and inhibiting cancer cells. ACS Appl. Nano Mater. 2021, 4, 652–665. [Google Scholar] [CrossRef]
- Lu, H.; Tian, Z.; Cui, Y.; Liu, Z.; Ma, X. Chlorogenic acid: A comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanism of action, and future directions. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3130–3158. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Murasige, T.; Skoog, F. A revised medium for growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dewir, Y.H.; Dalawai, D.; Al-Suhaibani, N. Comparative physicochemical analysis of seed oils of wild cucumber (Cucumis sativus var. hardwickii (Royle) Alef.), cucumber (Cucumis sativus L. var. sativus), and gherkin (Cucumis aunguira L.). S. Afr. J. Bot. 2022, 145, 186–191. [Google Scholar] [CrossRef]
- Pekal, A.; Pyrzynska, K. Evaluation of aluminum complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef] [Green Version]
- Yadav, G.G.; Murthy, H.N.; Dewir, Y.H. Nutritional composition and in vitro antioxidant activities of seed kernel and seed oil of Balanites roxburghii: An underutilized species. Horticuturae 2022, 8, 798. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antidominant activity applying an improved ABTS radical cation decolourization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Han, J.-E.; Murthy, H.N.; Kim, J.-Y.; Kim, M.-J.; Jeong, T.-K.; Park, S.-Y. Production of Secondary Metabolites from Cell Cultures of Sageretia thea (Osbeck) M.C. Johnst. Using Balloon-Type Bubble Bioreactors. Plants 2023, 12, 1390. https://doi.org/10.3390/plants12061390
Kim J-H, Han J-E, Murthy HN, Kim J-Y, Kim M-J, Jeong T-K, Park S-Y. Production of Secondary Metabolites from Cell Cultures of Sageretia thea (Osbeck) M.C. Johnst. Using Balloon-Type Bubble Bioreactors. Plants. 2023; 12(6):1390. https://doi.org/10.3390/plants12061390
Chicago/Turabian StyleKim, Ji-Hye, Jong-Eun Han, Hosakatte Niranjana Murthy, Ja-Young Kim, Mi-Jin Kim, Taek-Kyu Jeong, and So-Young Park. 2023. "Production of Secondary Metabolites from Cell Cultures of Sageretia thea (Osbeck) M.C. Johnst. Using Balloon-Type Bubble Bioreactors" Plants 12, no. 6: 1390. https://doi.org/10.3390/plants12061390