

De Novo Transcriptome Assembly and Comparative Analysis of Differentially Expressed Genes Involved in Cold Acclimation and Freezing Tolerance of the Arctic Moss Aulacomnium turgidum (Wahlenb.) Schwaegr

Abstract
:1. Introduction
2. Results
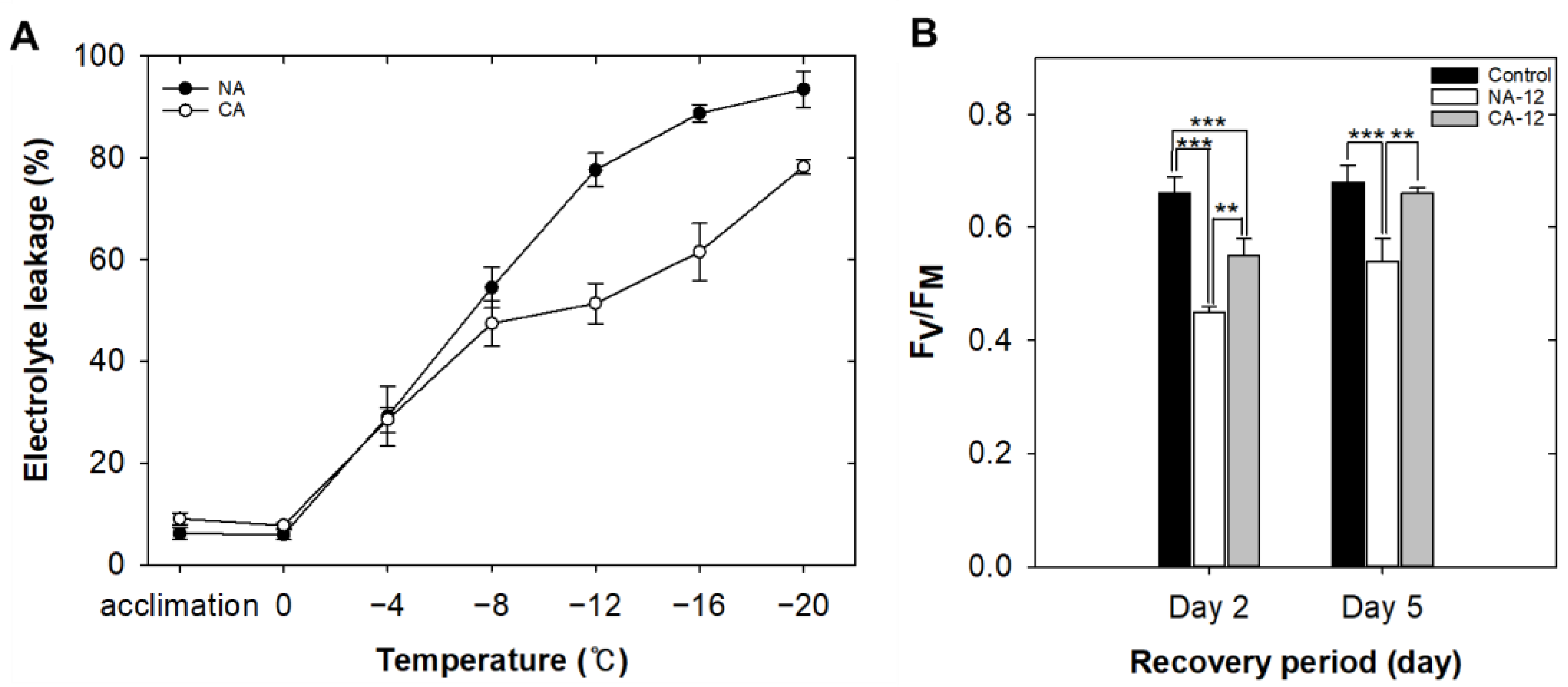
2.1. Physiological Changes Due to Cold Acclimation Treatment under Freezing Stress
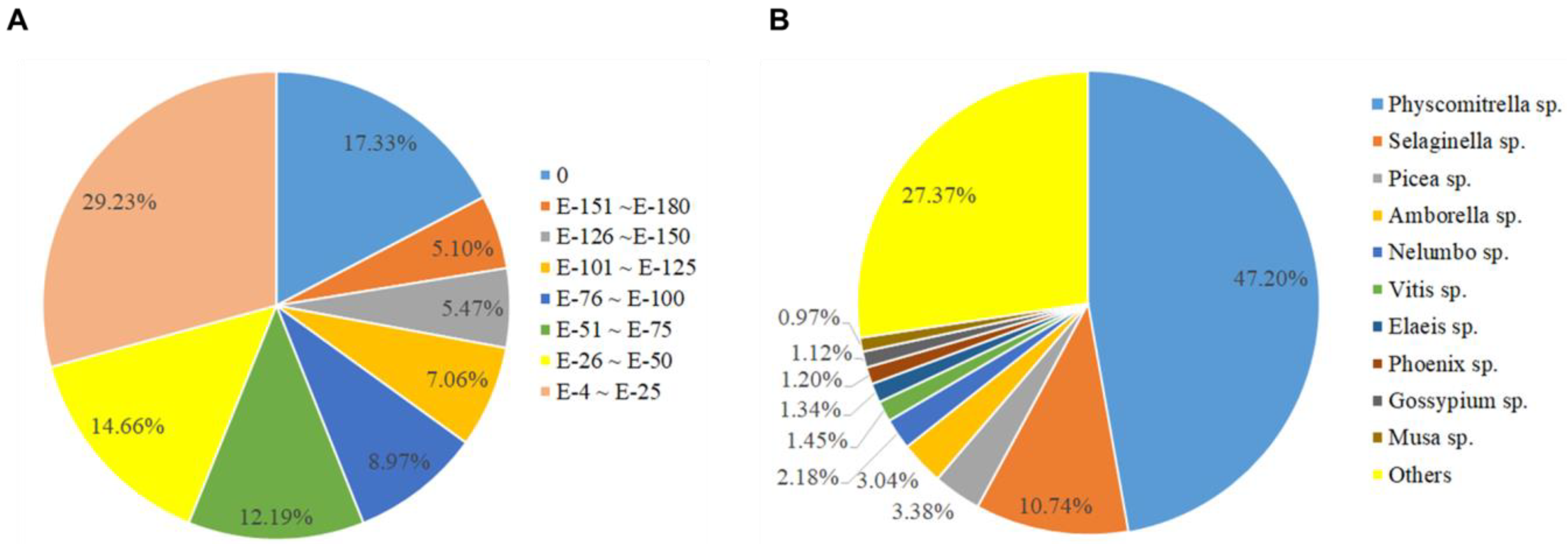
2.2. De Novo Assembly and Annotation of Assembled A. turgidum Protonema Transcripts
2.3. Gene Ontology (GO) Distribution and KEGG Pathway Analysis Using Whole Unigenes
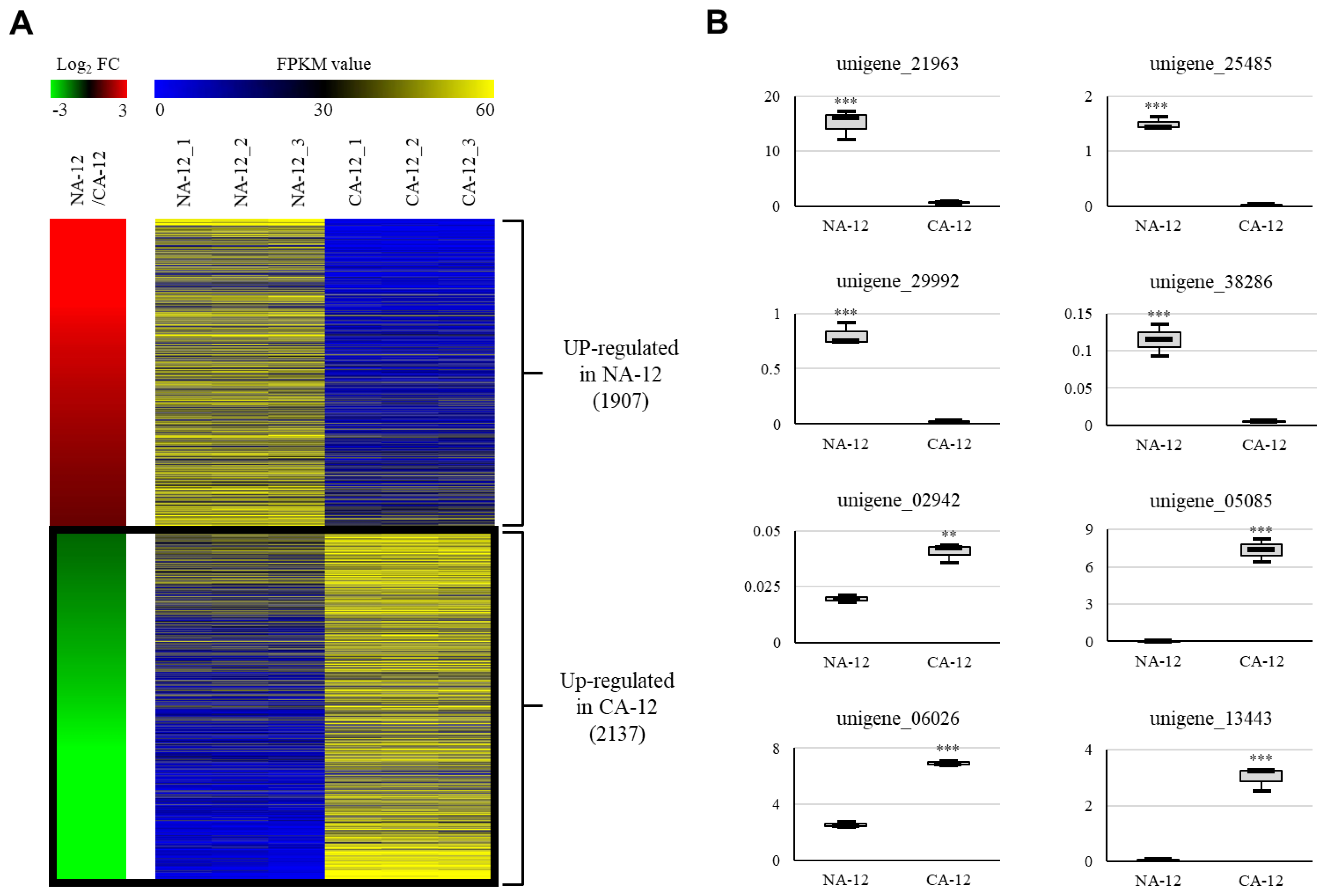
2.4. RNA-Seq Analysis Identified Candidate Unigenes Associated with Cold Acclimation and Freezing Stress
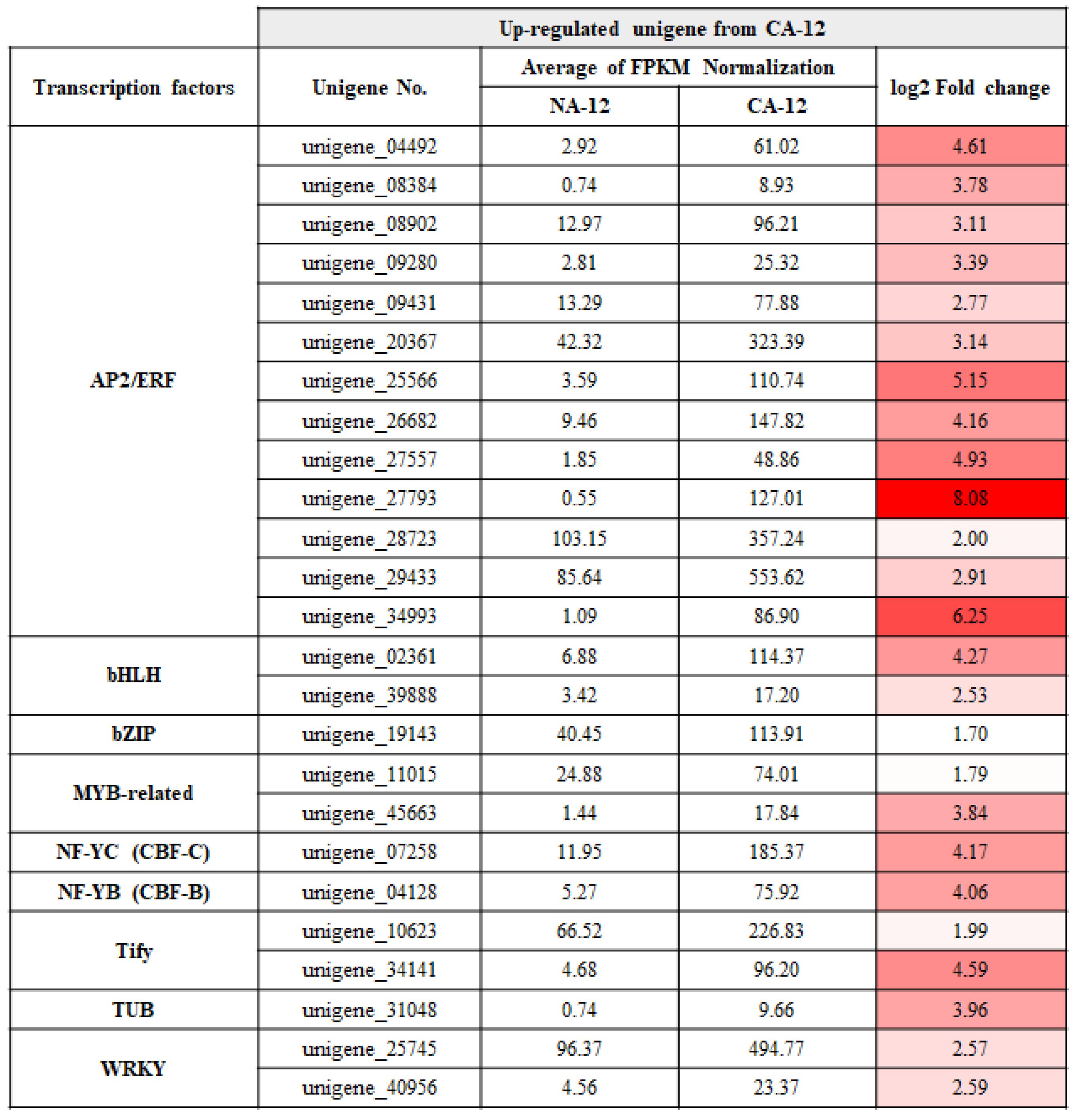
2.5. Transcription Factors Analysis of the Upregulated Group in the CA-12 Treatment
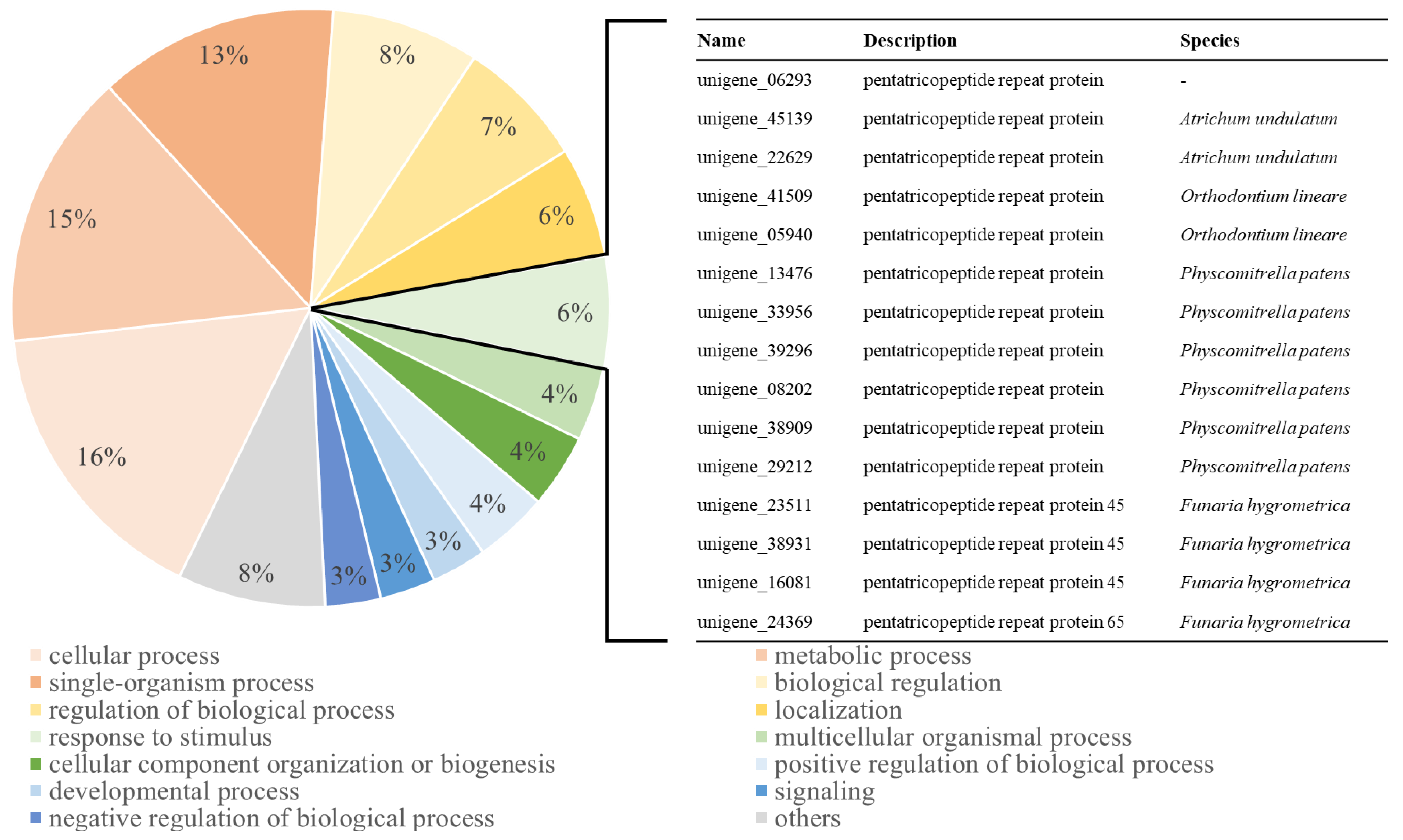
2.6. GO Distribution and KEGG Pathway Analysis of the Upregulated Group in CA-12
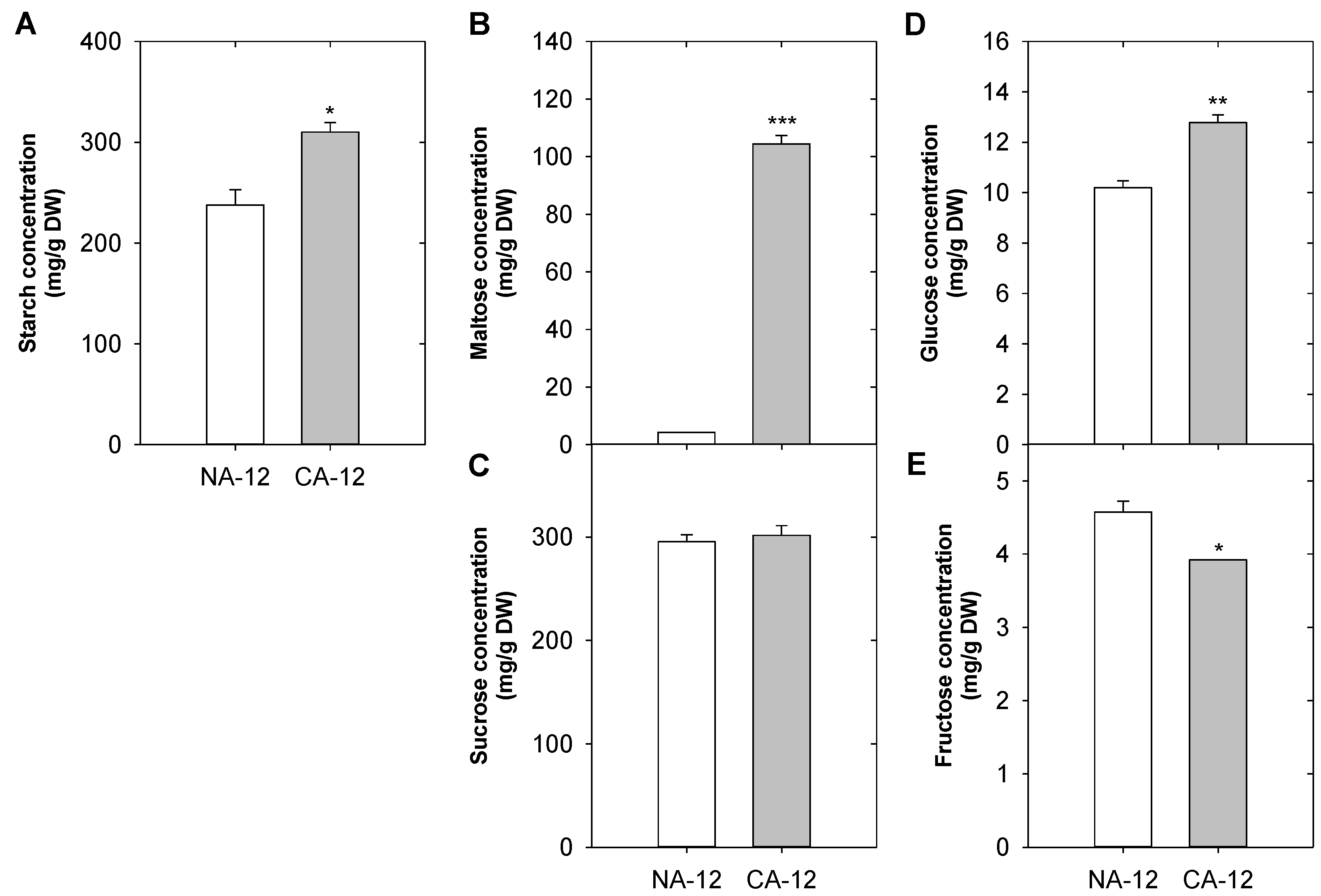
2.7. Differences in Starch and Sugar Concentrations According to Cold Acclimation and Freezing Stress
3. Discussion
3.1. Enhanced Freezing Tolerance by Cold Acclimation
3.2. AP2/ERF Family Transcription Factors Are Predicted to Contribute to Freezing Stress Tolerance of A. turgidum
3.3. AP2/ERF Family TFs Are Associated with Pentatricopeptide Repeat (PPR) Proteins and Carbohydrate Metabolism
3.4. Starch and Soluble Sugars Play an Important Role in Freezing Stress Tolerance
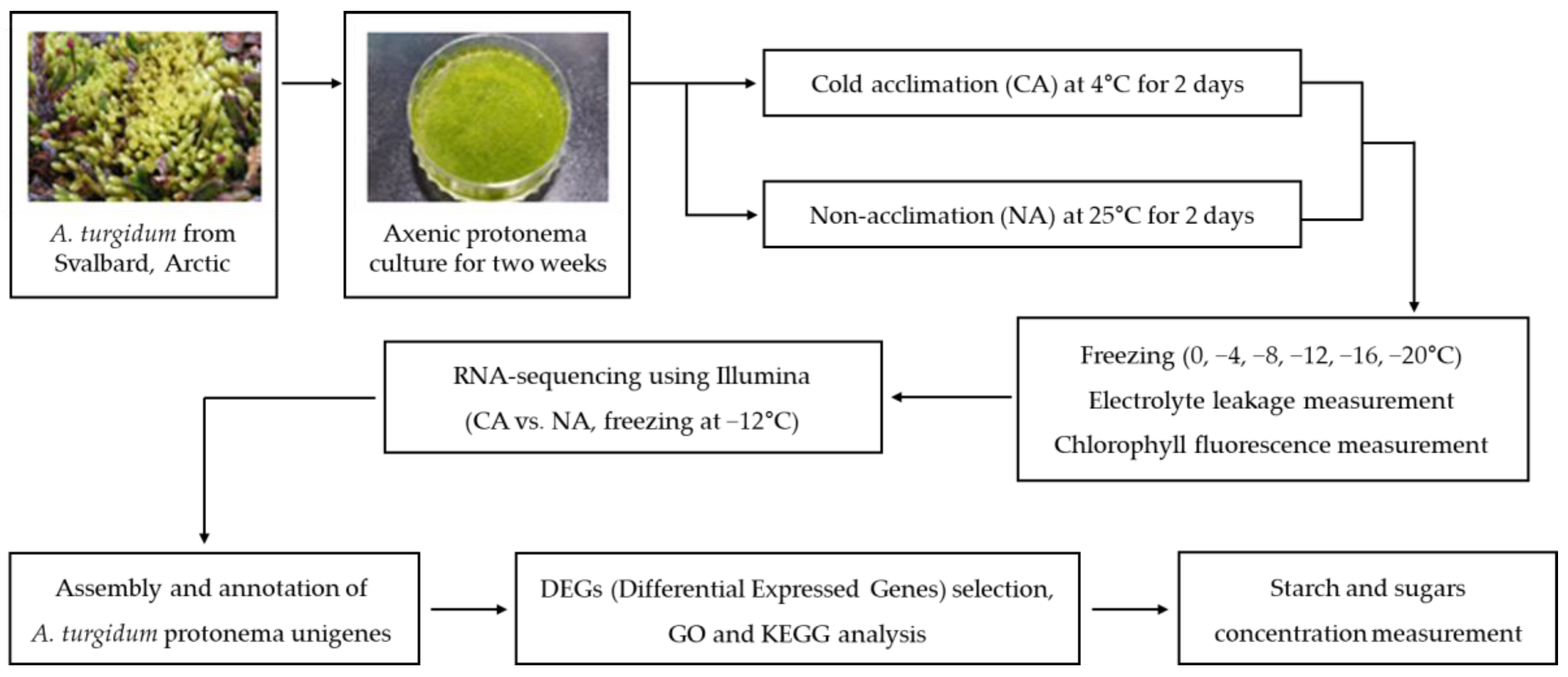
4. Materials and Methods
4.1. Sample Preparation
4.2. Cold Acclimation and Freezing Stress Treatment
4.3. Electrolyte Leakage Measurement
4.4. Chlorophyll Fluorescence Measurement
4.5. RNA Extraction and Library Preparation and Sequencing
4.6. De Novo Transcriptome Sequence Assembly and Functional Annotation
4.7. RNA-Seq Analysis
4.8. Starch and Sugars Concentration Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [Green Version]
- Gornall, J.L.; Jónsdóttir, I.S.; Woodin, S.J.; Van der Wal, R. Arctic mosses govern below-ground environment and ecosystem processes. Oecologia 2007, 153, 931–941. [Google Scholar] [CrossRef]
- GBIF Secretariat. GBIF Backbone Taxonomy. Available online: https://www.gbif.org/species/2675975 (accessed on 13 December 2022).
- La Farge, C.; Williams, K.H.; England, J.H. Regeneration of Little Ice Age bryophytes emerging from a polar glacier with implications of totipotency in extreme environments. Proc. Natl. Acad. Sci. USA 2013, 110, 9839–9844. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Lee, H.; Kang, P.S.; Huang, X.; Yim, J.H.; Lee, H.K.; Kim, I.C. Complementary DNA library construction and expressed sequence tag analysis of an Arctic moss, Aulacomnium turgidum . Polar Biol. 2010, 33, 617–626. [Google Scholar] [CrossRef]
- Kang, P.; Cho, S.M.; Lee, J.; Yim, J.H.; Lee, J.H.; Lee, H. The complete mitogenome of the Arctic moss Aulacomnium turgidum (Wahlenb.) Schwaegr. Mitochondrial DNA Part B 2019, 4, 3446–3447. [Google Scholar] [CrossRef]
- Levitt, J. Chilling, Freezing, and High Temperature Stresses. In Responses of Plants to Environmental Stress, 2nd ed.; Academic Press: New York, NY, USA, 1980; Volume 1, p. 497. [Google Scholar]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Choi, C. Mitochondrial network determines intracellular ROS dynamics and sensitivity to oxidative stress through switching inter-mitochondrial messengers. PLoS ONE 2011, 6, e23211. [Google Scholar] [CrossRef]
- Parvanova, D.; Ivanov, S.; Konstantinova, T.; Karanov, E.; Atanassov, A.; Tsvetkov, T.; Alexieva, V.; Djilianov, D. Transgenic tobacco plants accumulating osmolytes show reduced oxidative damage under freezing stress. Plant Physiol. Biochem. 2004, 42, 57–63. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Mitochondrial oxidative stress: Implications for cell death. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 143–183. [Google Scholar] [CrossRef]
- Hashempour, A.; Ghasemnezhad, M.; Sohani, M.M.; Ghazvini, R.F.; Abedi, A. Effects of freezing stress on the expression of fatty acid desaturase (FAD2, FAD6 and FAD7) and beta-glucosidase (BGLC) genes in tolerant and sensitive olive cultivars. Russ. J. Plant Physiol. 2019, 66, 214–222. [Google Scholar] [CrossRef]
- Artus, N.N.; Uemura, M.; Steponkus, P.L.; Gilmour, S.J.; Lin, C.; Thomashow, M.F. Constitutive expression of the cold-regulated Arabidopsis thaliana COR15a gene affects both chloroplast and protoplast freezing tolerance. Proc. Natl. Acad. Sci. USA 1996, 93, 13404–13409. [Google Scholar] [CrossRef] [Green Version]
- Sowemimo, O.; Borcherds, W.; Knox-Brown, P.; Rindfleisch, T.; Thalhammer, A.; Daughdrill, G. Evolution of transient helicity and disorder in late embryogenesis abundant protein COR15A. Biophys. J. 2019, 116, 473a. [Google Scholar] [CrossRef] [Green Version]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [Green Version]
- Konstantinova, T.; Parvanova, D.; Atanassov, A.; Djilianov, D. Freezing tolerant tobacco, transformed to accumulate osmoprotectants. Plant Sci. 2002, 163, 157–164. [Google Scholar] [CrossRef]
- Hughes, M.A.; Dunn, M.A. The molecular biology of plant acclimation to low temperature. J. Exp. Bot. 1996, 47, 291–305. [Google Scholar] [CrossRef] [Green Version]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Goffinet, B.; Buck, W.R.; Shaw, A.J. Morphology, anatomy, and classification of the Bryophyta. In Bryophyte Biology; Shaw, A.J., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 55–138. [Google Scholar]
- Fernandez-Pozo, N.; Haas, F.B.; Gould, S.B.; Rensing, S.A. An overview of bioinformatics, genomics, and transcriptomics resources for bryophytes. J. Exp. Bot. 2022, 73, 4291–4305. [Google Scholar] [CrossRef]
- Liu, S.; Fang, S.; Cong, B.; Li, T.; Yi, D.; Zhang, Z.; Zhao, L.; Zhang, P. The Antarctic moss Pohlia nutans genome provides insights into the evolution of bryophytes and the adaptation to extreme terrestrial habitats. Front. Plant Sci. 2022, 13, 920138. [Google Scholar] [CrossRef]
- Meleshko, O.; Martin, M.D.; Korneliussen, T.S.; Schröck, C.; Lamkowski, P.; Schmutz, J.; Healey, A.; Piatkowski, B.T.; Shaw, A.J.; Weston, D.J.; et al. Extensive genome-wide phylogenetic discordance is due to incomplete lineage sorting and not ongoing introgression in a rapidly radiated bryophyte genus. Mol. Biol. Evol. 2021, 38, 2750–2766. [Google Scholar] [CrossRef]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Szövényi, P.; Rensing, S.A.; Lang, D.; Wray, G.A.; Shaw, A.J. Generation-biased gene expression in a bryophyte model system. Mol. Biol. Evol. 2011, 28, 803–812. [Google Scholar] [CrossRef]
- Gao, B.; Zhang, D.; Li, X.; Yang, H.; Zhang, Y.; Wood, A.J. De novo transcriptome characterization and gene expression profiling of the desiccation tolerant moss Bryum argenteum following rehydration. BMC Genom. 2015, 16, 416. [Google Scholar] [CrossRef] [Green Version]
- Szövényi, P.; Perroud, P.F.; Symeonidi, A.; Stevenson, S.; Quatrano, R.S.; Rensing, S.A.; Cuming, A.C.; McDaniel, S.F. De novo assembly and comparative analysis of the Ceratodon purpureus transcriptome. Mol. Ecol. Resour. 2015, 15, 203–215. [Google Scholar] [CrossRef]
- Liu, S.; Fang, S.; Liu, C.; Zhao, L.; Cong, B.; Zhang, Z. Transcriptomics integrated with metabolomics reveal the effects of ultraviolet-B radiation on flavonoid biosynthesis in Antarctic moss. Front. Plant Sci. 2021, 12, 788377. [Google Scholar] [CrossRef]
- Chen, K.H.; Liao, H.L.; Arnold, A.E.; Korotkin, H.B.; Wu, S.H.; Matheny, P.B.; Lutzoni, F. Comparative transcriptomics of fungal endophytes in co-culture with their moss host Dicranum scoparium reveals fungal trophic lability and moss unchanged to slightly increased growth rates. New Phytol. 2022, 234, 1832–1847. [Google Scholar] [CrossRef]
- Carpenter, E.J.; Matasci, N.; Ayyampalayam, S.; Wu, S.; Sun, J.; Yu, J.; Vieira, F.R.J.; Bowler, C.; Dorrell, R.G.; Gitzendanner, M.A.; et al. Access to RNA-Sequencing Data from 1173 Plant Species: The 1000 Plant Transcriptomes Initiative (1KP). GigaScience 2019, 8, giz126. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Zhu, J.K. Response to the Abiotic Environment. In Arabidopsis: A Laboratory Manual; Weigel, D., Glazebrook, J., Eds.; Cold Spring Harbor Laboratory Press: Laurel Hollow, NY, USA, 2002; pp. 71–75. [Google Scholar]
- Kennedy, A.D. Photosynthetic response of the Antarctic moss Polytrichum alpestre Hoppe to low temperatures and freeze-thaw stress. Polar Biol. 1993, 13, 271–279. [Google Scholar] [CrossRef]
- Alberdi, M.; Bravo, L.A.; Gutiérrez, A.; Gidekel, M.; Corcuera, L.J. Ecophysiology of Antarctic vascular plants. Physiol. Plant. 2002, 115, 479–486. [Google Scholar] [CrossRef]
- Bokhorst, S.; Huiskes, A.; Convey, P.; Aerts, R. The effect of environmental change on vascular plant and cryptogam communities from the Falkland Islands and the Maritime Antarctic. BMC Ecol. 2007, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Fracheboud, Y.; Leipner, J. The application of chlorophyll fluorescence to study light, temperature, and drought stress. In Practical Applications of Chlorophyll Fluorescence in Plant Biology; De Ell, J.R., Toivonen, P.M.A., Eds.; Springer: Boston, MA, USA, 2013; pp. 125–150. [Google Scholar]
- Rapacz, M. Chlorophyll a fluorescence transient during freezing and recovery in winter wheat. Photosynthetica 2007, 45, 409–418. [Google Scholar] [CrossRef]
- Vosnjak, M.; Sircelj, H.; Vodnik, D.; Usenik, V. Physio-biochemical responses of sweet cherry leaf to natural cold conditions. Plants 2022, 11, 3507. [Google Scholar] [CrossRef]
- Gonzalez, D.H. Introduction to transcription factor structure and function. In Plant Transcription Factors; Gonzalez, D.H., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 3–11. [Google Scholar]
- Inukai, S.; Kock, K.H.; Bulyk, M.L. Transcription factor–DNA binding: Beyond binding site motifs. Curr. Opin. Genet. Dev. 2017, 43, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef]
- Liu, J.; Shi, Y.; Yang, S. Insights into the regulation of C-repeat binding factors in plant cold signaling. J. Integr. Plant Biol. 2018, 60, 780–795. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis . Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.G.; Zhang, W.K.; He, S.J.; Zhang, J.S.; Liu, Q.; Chen, S.Y. An EREBP/AP2-type protein in Triticum aestivum was a DRE-binding transcription factor induced by cold, dehydration and ABA stress. Theor. Appl. Genet. 2003, 106, 923–930. [Google Scholar] [CrossRef]
- Lv, K.; Li, J.; Zhao, K.; Chen, S.; Nie, J.; Zhang, W.; Liu, G.; Wei, H. Overexpression of an AP2/ERF family gene, BpERF13, in birch enhances cold tolerance through upregulating CBF genes and mitigating reactive oxygen species. Plant Sci. 2020, 292, 110375. [Google Scholar] [CrossRef]
- Moon, S.J.; Min, M.K.; Kim, J.; Kim, D.Y.; Yoon, I.S.; Kwon, T.R.; Byun, M.O.; Kim, B.G. Ectopic expression of OsDREB1G, a member of the OsDREB1 subfamily, confers cold stress tolerance in rice. Front. Plant Sci. 2019, 10, 297. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought-and low-temperature-responsive gene expression, respectively, in Arabidopsis . Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [Green Version]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt-and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA 2/Ethylene Responsive Factor (AP 2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis . Front. Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Lurin, C.; Andrés, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyère, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B.; et al. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089–2103. [Google Scholar] [CrossRef] [Green Version]
- Okuda, K.; Myouga, F.; Motohashi, R.; Shinozaki, K.; Shikanai, T. Conserved domain structure of pentatricopeptide repeat proteins involved in chloroplast RNA editing. Proc. Natl. Acad. Sci. USA 2007, 104, 8178–8183. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, N.; Hattori, M.; Andres, C.; Iida, K.; Lurin, C.; Schmitz-Linneweber, C.; Sugita, M.; Small, I. On the expansion of the pentatricopeptide repeat gene family in plants. Mol. Biol. Evol. 2008, 25, 1120–1128. [Google Scholar] [CrossRef]
- Sugita, M.; Ichinose, M.; Ide, M.; Sugita, C. Architecture of the PPR gene family in the moss Physcomitrella patens . RNA Biol. 2013, 10, 1439–1445. [Google Scholar] [CrossRef] [Green Version]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from chloroplasts converge to regulate nuclear gene expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- Jiang, S.C.; Mei, C.; Liang, S.; Yu, Y.T.; Lu, K.; Wu, Z.; Wang, X.F.; Zhang, D.P. Crucial roles of the pentatricopeptide repeat protein SOAR1 in Arabidopsis response to drought, salt and cold stresses. Plant Mol. Biol. 2015, 88, 369–385. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.H.; Byun, M.Y.; Oh, H.G.; Kim, S.J.; Lee, J.; Park, H.; Lee, H.; Kim, W.T. Poaceae Type II galactinol synthase 2 from Antarctic flowering plant Deschampsia antarctica and rice improves cold and drought tolerance by accumulation of raffinose family oligosaccharides in transgenic rice plants. Plant Cell Physiol. 2020, 61, 88–104. [Google Scholar] [CrossRef]
- Park, S.; Lee, C.M.; Doherty, C.J.; Gilmour, S.J.; Kim, Y.; Thomashow, M.F. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Plant J. 2015, 82, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Huijser, C.; Kortstee, A.; Pego, J.; Weisbeek, P.; Wisman, E.; Smeekens, S. The Arabidopsis SUCROSE UNCOUPLED-6 gene is identical to ABSCISIC ACID INSENSITIVE-4: Involvement of abscisic acid in sugar responses. Plant J. 2000, 23, 577–585. [Google Scholar] [CrossRef]
- Najafi, S.; Sorkheh, K.; Nasernakhaei, F. Characterization of the APETALA2/Ethylene-responsive factor (AP2/ERF) transcription factor family in sunflower. Sci. Rep. 2018, 8, 11576. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, F.; Guy, C.L. β-Amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar] [CrossRef] [Green Version]
- Tarkowski, Ł.P.; Van den Ende, W. Cold tolerance triggered by soluble sugars: A multifaceted countermeasure. Front. Plant Sci. 2015, 6, 203. [Google Scholar] [CrossRef] [Green Version]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [Green Version]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, F.; Guy, C.L. RNA interference of Arabidopsis beta-amylase8 prevents maltose accumulation upon cold shock and increases sensitivity of PSII photochemical efficiency to freezing stress. Plant J. 2005, 44, 730–743. [Google Scholar] [CrossRef]
- Nagao, M.; Minami, A.; Arakawa, K.; Fujikawa, S.; Takezawa, D. Rapid degradation of starch in chloroplasts and concomitant accumulation of soluble sugars associated with ABA-induced freezing tolerance in the moss Physcomitrella patens . J. Plant Physiol. 2005, 162, 169–180. [Google Scholar] [CrossRef]
- Fürtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of plant metabolism during cold acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef] [Green Version]
- Santarius, K.A. The protective effect of sugars on chloroplast membranes during temperature and water stress and its relationship to frost, desiccation and heat resistance. Planta 1973, 113, 105–114. [Google Scholar] [CrossRef]
- Sauter, J.J.; Van Cleve, B. Occurrence of a maltose pool and of maltase in poplar wood (Populus x canadensis robusta) during fall. J. Plant Physiol. 1993, 141, 248–250. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Laporte, M.; Lu, Y.; Weise, S.; Weber, A.P.M. Engineering plants for elevated CO2: A relationship between starch degradation and sugar sensing. Plant Biol. 2004, 6, 280–288. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, J.; Fan, Z.H.; Xiao, P.; Liu, S.N.; Li, R.P.; Zhu, W.B.; Huang, L. MAL62 overexpression enhances freezing tolerance of baker’s yeast in lean dough by enhancing Tps1 activity and maltose metabolism. J. Agric. Food Chem. 2019, 67, 8986–8993. [Google Scholar] [CrossRef]
- Adam, S.; Murthy, S.D.S. Effect of cold stress on photosynthesis of plants and possible protection mechanisms. In Approaches to Plant Stress and Their Management; Gaur, R., Sharma, P., Eds.; Springer: New Delhi, India, 2014; pp. 219–226. [Google Scholar]
- Taïbi, K.; Del Campo, A.D.; Vilagrosa, A.; Bellés, J.M.; López-Gresa, M.P.; López-Nicolás, J.M.; Mulet, J.M. Distinctive physiological and molecular responses to cold stress among cold-tolerant and cold-sensitive Pinus halepensis seed sources. BMC Plant Biol. 2018, 18, 236. [Google Scholar] [CrossRef] [Green Version]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; García-Agustín, P.; González-Bosch, C. Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U. Priming of induced plant defense responses. Adv. Bot. Res. 2009, 51, 361–395. [Google Scholar]
- NDong, C.; Danyluk, J.; Wilson, K.E.; Pocock, T.; Huner, N.P.; Sarhan, F. Cold-regulated cereal chloroplast late embryogenesis abundant-like proteins. Molecular characterization and functional analyses. Plant Physiol. 2002, 129, 1368–1381. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Nagao, M.; Arakawa, K.; Fujikawa, S.; Takezawa, D. Abscisic acid-induced freezing tolerance in the moss Physcomitrella patens is accompanied by increased expression of stress-related genes. J. Plant Physiol. 2003, 160, 475–483. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. Mev: Multiexperiment viewer. In Biomedical Informatics for Cancer Research; Ochs, M.F., Casagrande, J.T., Davuluri, R.V., Eds.; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar]
- Kobylińska, A.; Borek, S.; Posmyk, M.M. Melatonin redirects carbohydrates metabolism during sugar starvation in plant cells. J. Pineal Res. 2018, 64, e12466. [Google Scholar] [CrossRef]
- Menezes, F.O.; Carvalho, V.; Moreira, V.A.; Rigui, A.P.; Gaspar, M.; Nievola, C.C. Juvenile plants of an epiphytic bromeliad exhibit rapid changes in carbohydrate content and antioxidant capacity to cope with water withholding. Theor. Exp. Plant Physiol. 2020, 32, 89–98. [Google Scholar] [CrossRef]
- Maturilli, M.; Herber, A.; König-Langlo, G. Climatology and time series of surface meteorology in Ny-Ålesund, Svalbard. Earth Syst. Sci. Data 2013, 5, 155–163. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | No. Raw Reads | No. Trimmed Reads | % of Reads after Trimming | Size of Trimmed Reads (bp) | GC (%) | Q30 (%) |
|---|---|---|---|---|---|---|
| NA-12_1 | 3,529,180 | 3,305,320 | 93.66 | 814,851,894 | 49.13 | 93.77 |
| NA-12_2 | 3,759,002 | 3,536,726 | 94.09 | 879,302,307 | 49.63 | 93.84 |
| NA-12_3 | 3,446,114 | 3,202,404 | 92.93 | 789,298,951 | 49.5 | 93.61 |
| CA-12_1 | 3,636,552 | 3,421,010 | 94.07 | 848,381,538 | 49.24 | 93.8 |
| CA-12_2 | 3,241,544 | 3,006,906 | 92.76 | 732,304,413 | 49.22 | 93.52 |
| CA-12_3 | 3,727,552 | 3,518,610 | 94.39 | 874,412,690 | 49.66 | 93.86 |
| Contig | Unigene | |
|---|---|---|
| Total Length (bp) | 157,581,303 | 24,753,852 |
| No. Sequence | 116,979 | 45,796 |
| Max. Length (bp) | 19,624 | 17,139 |
| Mean Length (bp) | 1347 | 541 |
| N50 (bp) | 2481 | 1017 |
| N90 (bp) | 507 | 207 |
| N90 Sequence No. | 73,457 | 31,863 |
| GC% | 46.5 | 49.2 |
| Class | Pathway | ID | DEG | All Genes | p-Value | Corrected p-Value |
|---|---|---|---|---|---|---|
| Category/Metabolism | ||||||
| Carbohydrate metabolism | Starch and sucrose metabolism | ppp00500 | 32 | 198 | 1.46 × 10−11 | 4.57 × 10−10 |
| Glycolysis/gluconeogenesis | ppp00010 | 26 | 172 | 4.06 × 10−9 | 7.63 × 10−8 | |
| Amino sugar and nucleotide sugar metabolism | ppp00520 | 23 | 137 | 5.49 × 10−9 | 8.61 × 10−8 | |
| Galactose metabolism | ppp00052 | 13 | 52 | 2.32 × 10−7 | 3.12 × 10−6 | |
| Pentose phosphate pathway | ppp00030 | 16 | 86 | 3.36 × 10−7 | 3.91 × 10−6 | |
| Butanoate metabolism | ppp00650 | 9 | 24 | 1.06 × 10−6 | 9.93 × 10−6 | |
| Fructose and mannose metabolism | ppp00051 | 12 | 88 | 1.46 × 10−4 | 7.64 × 10−4 | |
| Ascorbate and aldarate metabolism | ppp00053 | 8 | 52 | 9.09 × 10−4 | 3.88 × 10−3 | |
| Pyruvate metabolism | ppp00620 | 12 | 111 | 9.86 × 10−4 | 4.03 × 10−3 | |
| Glyoxylate and dicarboxylate metabolism | ppp00630 | 11 | 107 | 2.27 × 10−3 | 8.54 × 10−3 | |
| Amino acid metabolism | Phenylalanine metabolism | ppp00360 | 12 | 45 | 3.74 × 10−7 | 3.91 × 10−6 |
| Alanine, aspartate, and glutamate metabolism | ppp00250 | 12 | 59 | 4.38 × 10−6 | 3.74 × 10−5 | |
| Tyrosine metabolism | ppp00350 | 8 | 43 | 2.96 × 10−4 | 1.39 × 10−3 | |
| Lipid metabolism | Fatty acid biosynthesis | ppp00061 | 11 | 60 | 2.56 × 10−5 | 2.01 × 10−4 |
| Glycerophospholipid metabolism | ppp00564 | 13 | 87 | 3.37 × 10−5 | 2.43 × 10−4 | |
| Cutin, suberine, and wax biosynthesis | ppp00073 | 5 | 11 | 1.41 × 10−4 | 7.64 × 10−4 | |
| Biosynthesis of other secondary metabolites | Phenylpropanoid biosynthesis | ppp00940 | 15 | 117 | 4.34 × 10−5 | 2.84 × 10−4 |
| Metabolism of other amino acids | Taurine and hypotaurine metabolism | ppp00430 | 5 | 8 | 4.54 × 10−5 | 2.84 × 10−4 |
| beta-Alanine metabolism | ppp00410 | 6 | 36 | 2.76 × 10−3 | 9.97 × 10−3 | |
| Energy metabolism | Carbon fixation in photosynthetic organisms | ppp00710 | 15 | 125 | 8.56 × 10−5 | 5.03 × 10−4 |
| Metabolism of cofactors and vitamins | Ubiquinone and other terpenoid-quinone biosynthesis | ppp00130 | 8 | 40 | 1.92 × 10−4 | 9.48 × 10−4 |
| Category/Cellular Processes | ||||||
| Transport and catabolism | Peroxisome | ppp04146 | 11 | 96 | 1.03 × 10−3 | 4.02 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, P.; Yoo, Y.-H.; Kim, D.-I.; Yim, J.H.; Lee, H. De Novo Transcriptome Assembly and Comparative Analysis of Differentially Expressed Genes Involved in Cold Acclimation and Freezing Tolerance of the Arctic Moss Aulacomnium turgidum (Wahlenb.) Schwaegr. Plants 2023, 12, 1250. https://doi.org/10.3390/plants12061250
Kang P, Yoo Y-H, Kim D-I, Yim JH, Lee H. De Novo Transcriptome Assembly and Comparative Analysis of Differentially Expressed Genes Involved in Cold Acclimation and Freezing Tolerance of the Arctic Moss Aulacomnium turgidum (Wahlenb.) Schwaegr. Plants. 2023; 12(6):1250. https://doi.org/10.3390/plants12061250
Chicago/Turabian StyleKang, Pilsung, Yo-Han Yoo, Dong-Il Kim, Joung Han Yim, and Hyoungseok Lee. 2023. "De Novo Transcriptome Assembly and Comparative Analysis of Differentially Expressed Genes Involved in Cold Acclimation and Freezing Tolerance of the Arctic Moss Aulacomnium turgidum (Wahlenb.) Schwaegr" Plants 12, no. 6: 1250. https://doi.org/10.3390/plants12061250