
Cadmium Transport in Maize Root Segments Using a Classical Physiological Approach: Evidence of Influx Largely Exceeding Efflux in Subapical Regions
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
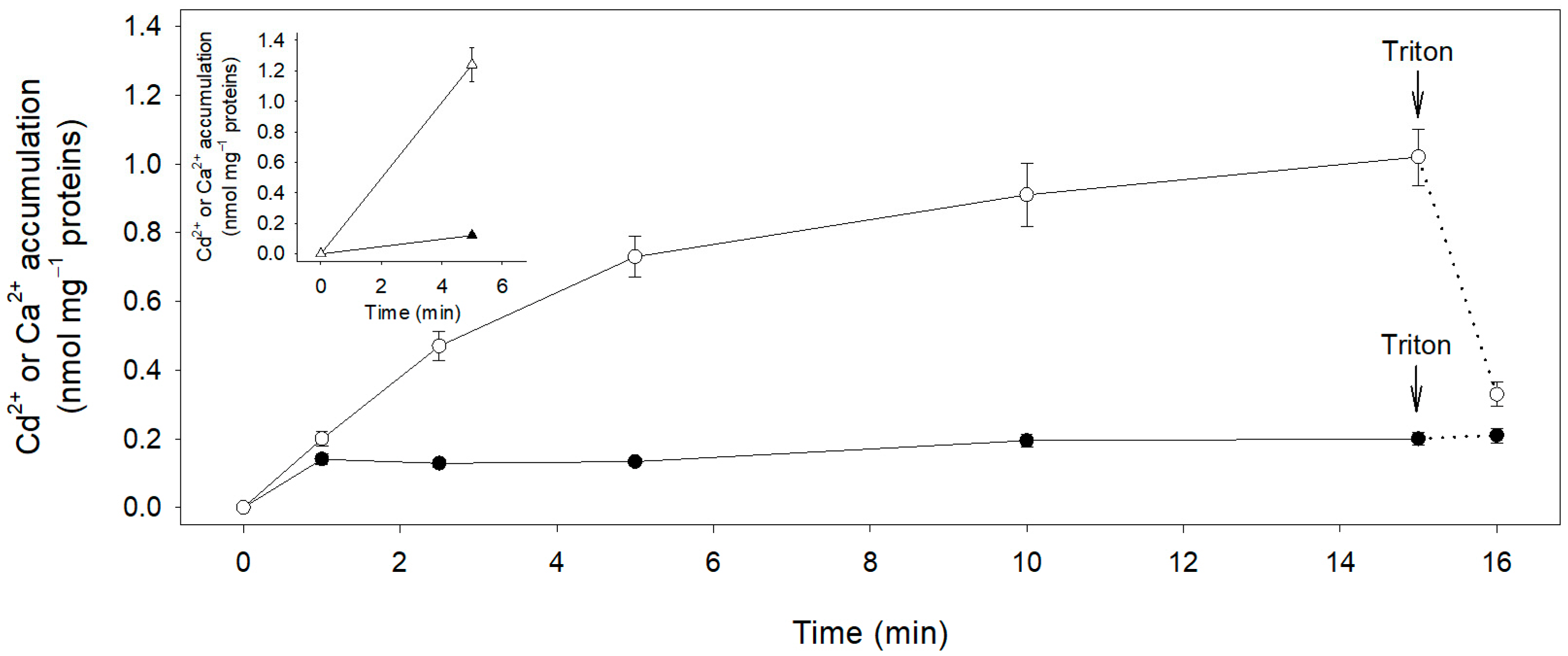
2.1. Short Term Cd2+ Influx in Maize Root Segments
2.2. Short Term Ca2+ Influx in Maize Root Segments
2.3. Effect of Ca2+ on Cd2+ Influx in Maize Root Segments
2.4. Cd2+ and Ca2+ Efflux from Subapical Maize Root Segments
2.5. Cd2+ Transport in Plasma Membrane Vesicles from Maize Root Cortical Cells
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Measurement of Cd2+ Influx
4.3. Measurement of Ca2+ Influx
4.4. Pulse and Chase Experiments with (109Cd2+)Cd2+ or (45Ca2+)Ca2+
4.5. Plasma Membrane Vesicle Isolation
4.6. Measurement of Ca2+ and Cd2+ Uptake in Plasma Membrane Vesicles
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nordberg, G.F. Historical perspectives on cadmium toxicology. Toxicol. Appl. Pharmacol. 2009, 238, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J.; Steinnes, E. Anthropogenic additions of cadmium to soils. In Cadmium in Soils and Plants; McLaughlin, M.J., Singh, B.R., Eds.; Developments in Plant and Soil Sciences; Springer Netherlands: Dordrecht, The Netherlands, 1999; pp. 97–123. ISBN 978-94-011-4473-5. [Google Scholar]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant Science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; Parker, D.R.; Clarke, J.M. Metals and micronutrients—Food safety issues. Field Crops Res. 1999, 60, 143–163. [Google Scholar] [CrossRef]
- Welch, R.M.; Norvell, W.A. Mechanisms of cadmium uptake, translocation and deposition in plants. In Cadmium in Soils and Plants; McLaughlin, M.J., Singh, B.R., Eds.; Developments in Plant and Soil Sciences; Springer Netherlands: Dordrecht, The Netherlands, 1999; pp. 125–150. ISBN 978-94-011-4473-5. [Google Scholar]
- Sterckeman, T.; Thomine, S. Mechanisms of cadmium accumulation in plants. Crit. Rev. Plant Sci. 2020, 39, 322–359. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Korshunova, Y.O.; Eide, D.; Clark, W.G.; Guerinot, M.L. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol. Biol. 1999, 40, 37–44. [Google Scholar] [CrossRef]
- Eide, D.; Broderius, M.; Fett, J.; Guerinot, M.L. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 5624–5628. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.E.; Eide, D.J.; Guerinot, M.L. Altered selectivity in an Arabidopsis metal transporter. Proc. Natl. Acad. Sci. USA 2000, 97, 12356–12360. [Google Scholar] [CrossRef] [Green Version]
- Ajeesh Krishna, T.P.; Maharajan, T.; Victor Roch, G.; Ignacimuthu, S.; Antony Ceasar, S. Structure, function, regulation and phylogenetic relationship of ZIP family transporters of plants. Front. Plant Sci. 2020, 11, 662. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Shin, R.; Eide, D.J.; Schachtman, D.P. Differential metal selectivity and gene expression of two zinc transporters from rice. Plant Physiol. 2003, 133, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake 478 in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation 482 mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in Rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Hinkle, P.; Kinsella, P.; Osterhoudt, K. Cadmium uptake and toxicity via voltage-sensitive calcium channels. J. Biol. Chem. 1987, 262, 16333–16337. [Google Scholar] [CrossRef]
- Usai, C.; Barberis, A.; Moccagatta, L.; Marchetti, C. Pathways of cadmium influx in mammalian neurons. J. Neurochem. 1999, 72, 2154–2156. [Google Scholar] [CrossRef] [PubMed]
- Souza, V.; Bucio, L.; Gutiérrez-Ruiz, M.C. Cadmium uptake by a human hepatic cell line (WRL-68 Cells). Toxicology 1997, 120, 215–220. [Google Scholar] [CrossRef]
- Jacobson, B.; Turner, J.E. The interaction of cadmium and certain other metal ions with proteins and nucleic acids. Toxicology 1980, 16, 1–37. [Google Scholar] [CrossRef]
- Choong, G.; Ying Liu, Y.; Templeton, D.M. Interplay of calcium and cadmium in mediating cadmium toxicity. Chem. Biol. Interact. 2014, 211, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Rivetta, A.; Negrini, N.; Cocucci, M. Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus L.) seed germination. Plant Cell Environ. 1997, 20, 600–608. [Google Scholar] [CrossRef]
- Jarvis, S.C.; Jones, L.H.P.; Hopper, M.J. Cadmium uptake from solution by plants and its transport from roots to shoots. Plant Soil 1976, 44, 179–191. [Google Scholar] [CrossRef]
- Zhao, F.; Hamon, R.E.; Lombi, E.; McLaughlin, M.J.; McGrath, S.P. Characteristics of cadmium uptake in two contrasting ecotypes of the hyperaccumulator Thlaspi caerulescens. J. Exp. Bot. 2002, 53, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ouyang, Y.; Fan, Y.; Qiu, B.; Zhang, G.; Zeng, F. The pathway of transmembrane cadmium influx via calcium-permeable channels and its spatial characteristics along rice root. J. Exp. Bot. 2018, 69, 5279–5291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grill, E.; Winnacker, E.-L.; Zenk, M.H. Phytochelatins, a class of heavy-metal-binding peptides from plants, are functionally analogous to metallothioneins. Proc. Natl. Acad. Sci. USA 1987, 84, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Zenk, M.H. Heavy metal detoxification in higher plants—A review. Gene 1996, 179, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.-F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.-H.; Pittman, J.K.; Barkla, B.J.; Shigaki, T.; Hirschi, K.D. The Arabidopsis Cax1 mutant exhibits impaired ion homeostasis, development, and hormonal responses and reveals interplay among vacuolar transporters. Plant Cell 2003, 15, 347–364. [Google Scholar] [CrossRef] [Green Version]
- Nocito, F.F.; Lancilli, C.; Dendena, B.; Lucchini, G.; Sacchi, G.A. Cadmium retention in rice roots is influenced by cadmium availability, chelation and translocation. Plant Cell Environ. 2011, 34, 994–1008. [Google Scholar] [CrossRef] [PubMed]
- Silver, S.; Phung, L.T. Bacterial heavy metal resistance: New surprises. Annu. Rev. Microbiol. 1996, 50, 753–789. [Google Scholar] [CrossRef] [PubMed]
- Maghrebi, M.; Baldoni, E.; Lucchini, G.; Vigani, G.; Valè, G.; Sacchi, G.A.; Nocito, F.F. Analysis of cadmium root retention for two contrasting rice accessions suggests an important role for OsHMA2. Plants 2021, 10, 806. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Morel, J.L. Cadmium uptake by Lupinus albus (L.): Cadmium excretion, a possible mechanism of cadmium tolerance. J. Plant Nutr. 1993, 16, 1921–1929. [Google Scholar] [CrossRef]
- Piñeros, M.A.; Shaff, J.E.; Kochian, L.V. Development, characterization, and application of a cadmium-selective microelectrode for the measurement of cadmium fluxes in roots of Thlaspi species and wheat. Plant Physiol. 1998, 116, 1393–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.-Z.; Tu, C.; Peijnenburg, W.J.G.M.; Luo, Y.-M. Characteristics of cadmium uptake and membrane transport in roots of intact wheat (Triticum aestivum L.) seedlings. Environ. Pollut. 2017, 221, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Ayub, A.; Hussain, S.; Waraich, E.A.; El-Esawi, M.A.; Ishfaq, M.; Ahmad, M.; Ali, N.; Maqsood, M.F. Cadmium toxicity in plants: Recent progress on morpho-physiological effects and remediation strategies. J. Soil Sci. Plant Nutr. 2022, 22, 212–269. [Google Scholar] [CrossRef]
- Deckert, J. Cadmium toxicity in plants: Is there any analogy to its carcinogenic effect in mammalian Cells? Biometals 2005, 18, 475–481. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Cadmium stress tolerance in crop plants. Plant Signal. Behav. 2011, 6, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Andresen, E.; Küpper, H. Cadmium toxicity in plants. In Cadmium: From Toxicity to Essentiality; Sigel, A., Sigel, H., Sigel, R.K., Eds.; Metal Ions in Life Sciences; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 395–413. [Google Scholar]
- Sanità di Toppi, L.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Ferri, A.; Lancilli, C.; Maghrebi, M.; Lucchini, G.; Sacchi, G.A.; Nocito, F.F. The sulfate supply maximizing Arabidopsis shoot growth is higher under long- than short-term exposure to cadmium. Front. Plant Sci. 2017, 8, 854. [Google Scholar] [CrossRef]
- Martelli, A.; Rousselet, E.; Dycke, C.; Bouron, A.; Moulis, J.-M. Cadmium toxicity in animal cells by interference with essential metals. Biochimie 2006, 88, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Cadmium Interaction with Animal Cells; Thévenod, F.; Petering, D.M.; Templeton, D.; Lee, W.-K.; Hartwig, A. (Eds.) Springer International Publishing: Cham, Switzerland, 2018; pp. 1–125. [Google Scholar]
- Sattelmacher, B. The Apoplast and its significance for plant mineral nutrition. New Phytol. 2001, 149, 167–192. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.J. Ion exchange properties of roots and ionic interactions within the root apoplasm: Their role in ion accumulation by plants. Bot. Rev. 1980, 46, 75–99. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Sanders, D.; Brownlee, C.; Harper, J.F. Communicating with calcium. Plant Cell 1999, 11, 691–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAinsh, M.R.; Pittman, J.K. Shaping the calcium signature. New Phytol. 2009, 181, 275–294. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Thor, K. Calcium—Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Askerlund, P.; Fredrikson, K.; Widell, S.; Sommarin, M.; Larsson, C. Sealed inside-out and right-side-out plasma membrane vesicles. Optimal conditions for formation and separation. Plant Physiol. 1990, 92, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Palmgren, M.; Morsomme, P. The plasma membrane H+-ATPase, a simple polypeptide with a long history. Yeast 2019, 36, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, J.; Pottosin, I.; Shabala, S.; Palmgren, M.; Shabala, S. Calcium efflux systems in stress signaling and adaptation in plants. Front. Plant Sci. 2011, 2, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, M.; Muto, S. Ca2+ pump and Ca2+ /H+ antiporter in plasma membrane vesicles isolated by aqueous two-phase partitioning from corn leaves. J. Membr. Biol. 1990, 114, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.Z.; Wang, H.W.; Huang, J.; Tian, A.G.; Wang, Y.J.; Zhang, J.S.; Chen, S.Y. A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Mol. Biol. 2005, 59, 809–820. [Google Scholar] [CrossRef]
- Clemens, S.; Antosiewicz, D.M.; Ward, J.M.; Schachtman, D.P.; Schroeder, J.I. The plant cDNA LCT1 mediates the uptake of calcium and cadmium in yeast. Pro. Natl. Acad. Sci. USA 1998, 95, 12043–12048. [Google Scholar] [CrossRef] [Green Version]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Q.; Nazir, M.M.; Ali, S.; Ouyang, Y.; Ye, S.; Zeng, F. Calcium plays a double-edged role in modulating cadmium uptake and translocation in rice. Int. J. Mol. Sci. 2020, 21, 8058. [Google Scholar] [CrossRef]
- Suzuki, N. Alleviation by calcium of cadmium-induced root growth inhibition in Arabidopsis seedlings. Plant Biotechnol. 2005, 22, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, N.; Saifullah; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Williams, L.E.; Mills, R.F. P1B-ATPases—An ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Kim, D.-Y.; Shim, D.; Song, W.-Y.; Lee, J.; Schroeder, J.I.; Kim, S.; Moran, N.; Lee, Y. Expression of the novel wheat gene TM20 confers enhanced cadmium tolerance to bakers’ yeast. J. Biol. Chem. 2008, 283, 15893–15902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Case, R.M.; Eisner, D.; Gurney, A.; Jones, O.; Muallem, S.; Verkhratsky, A. Evolution of calcium homeostasis: From birth of the first cell to an omnipresent signalling system. Cell Calcium 2007, 42, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [Green Version]
- Larsson, C.; Widell, S.; Kjellbom, P. Preparation of high-purity plasma membranes. Meth. Enzymol. 1987, 148, 558–568. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| [Ca2+] (µM) | [Cd2+] | |||||
|---|---|---|---|---|---|---|
| 10 µM | 50 µM | 100 µM | ||||
| Cd2+ influx (µmol h−1 g−1 FW) | Δ (%) | Cd2+ influx (µmol h−1 g−1 FW) | Δ (%) | Cd2+ influx (µmol h−1 g−1 FW) | Δ (%) | |
| 0 | 0.79 ± 0.04 a | 1.97 ± 0.03 a | 2.32 ± 0.12 a | |||
| 10 | 0.65 ± 0.03 b | −17.7 | 1.68 ± 0.02 b | −14.7 | 2.19 ± 0.08 a | −5.6 |
| 50 | 0.33 ± 0.02 c | −58.2 | 1.32 ± 0.09 c | −33.0 | 1.72 ± 0.04 b | −25.9 |
| 100 | 0.24 ± 0.01 d | −69.6 | 1.11 ± 0.02 d | −43.7 | 1.47 ± 0.04 c | −36.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivetta, A.; Pesenti, M.; Sacchi, G.A.; Nocito, F.F.; Cocucci, M. Cadmium Transport in Maize Root Segments Using a Classical Physiological Approach: Evidence of Influx Largely Exceeding Efflux in Subapical Regions. Plants 2023, 12, 992. https://doi.org/10.3390/plants12050992
Rivetta A, Pesenti M, Sacchi GA, Nocito FF, Cocucci M. Cadmium Transport in Maize Root Segments Using a Classical Physiological Approach: Evidence of Influx Largely Exceeding Efflux in Subapical Regions. Plants. 2023; 12(5):992. https://doi.org/10.3390/plants12050992
Chicago/Turabian StyleRivetta, Alberto, Michele Pesenti, Gian Attilio Sacchi, Fabio Francesco Nocito, and Maurizio Cocucci. 2023. "Cadmium Transport in Maize Root Segments Using a Classical Physiological Approach: Evidence of Influx Largely Exceeding Efflux in Subapical Regions" Plants 12, no. 5: 992. https://doi.org/10.3390/plants12050992