

Earlywood Anatomy Highlights the Prevalent Role of Winter Conditions on Radial Growth of Oak at Its Distribution Boundary in NW Iberia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
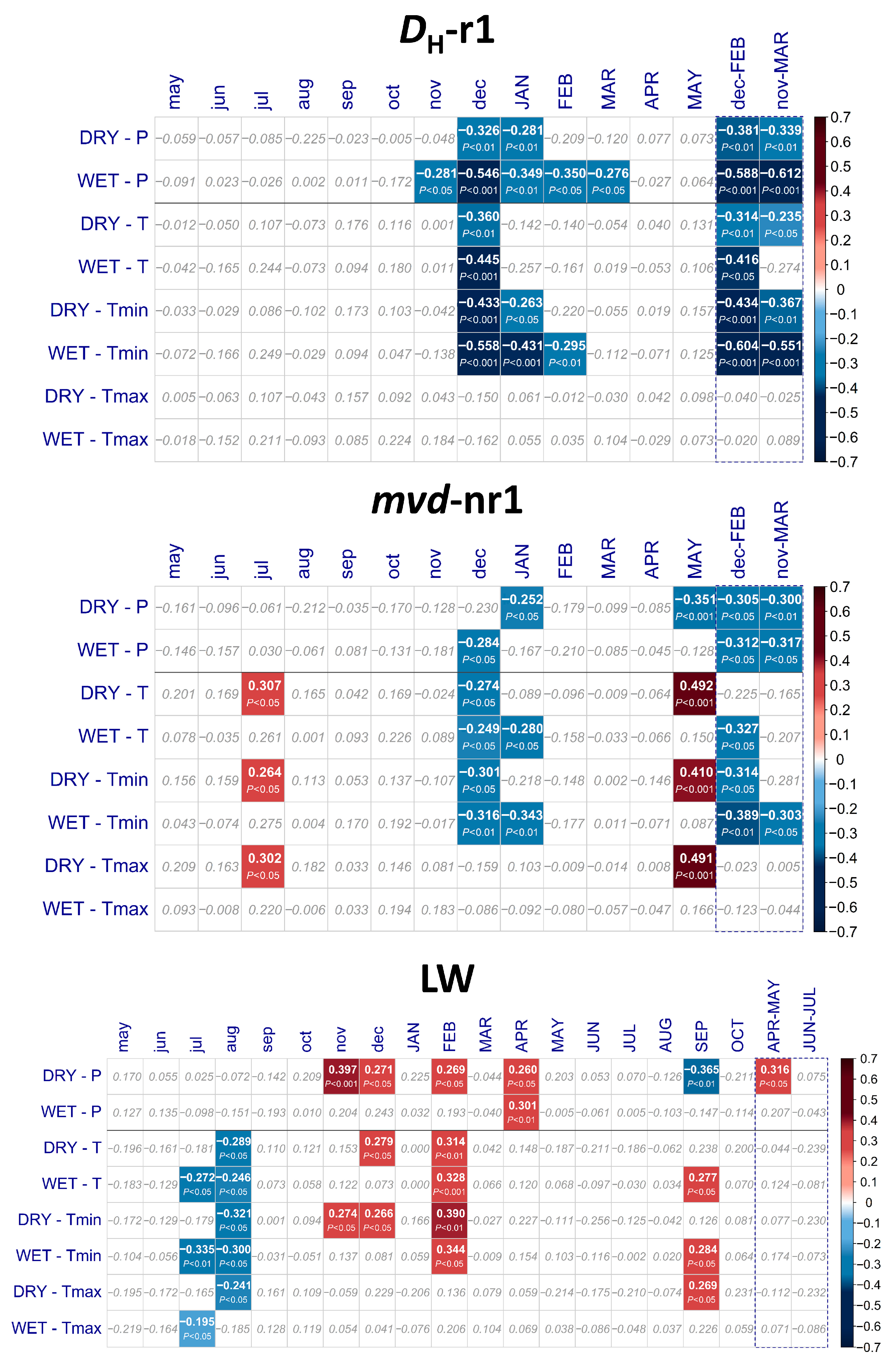
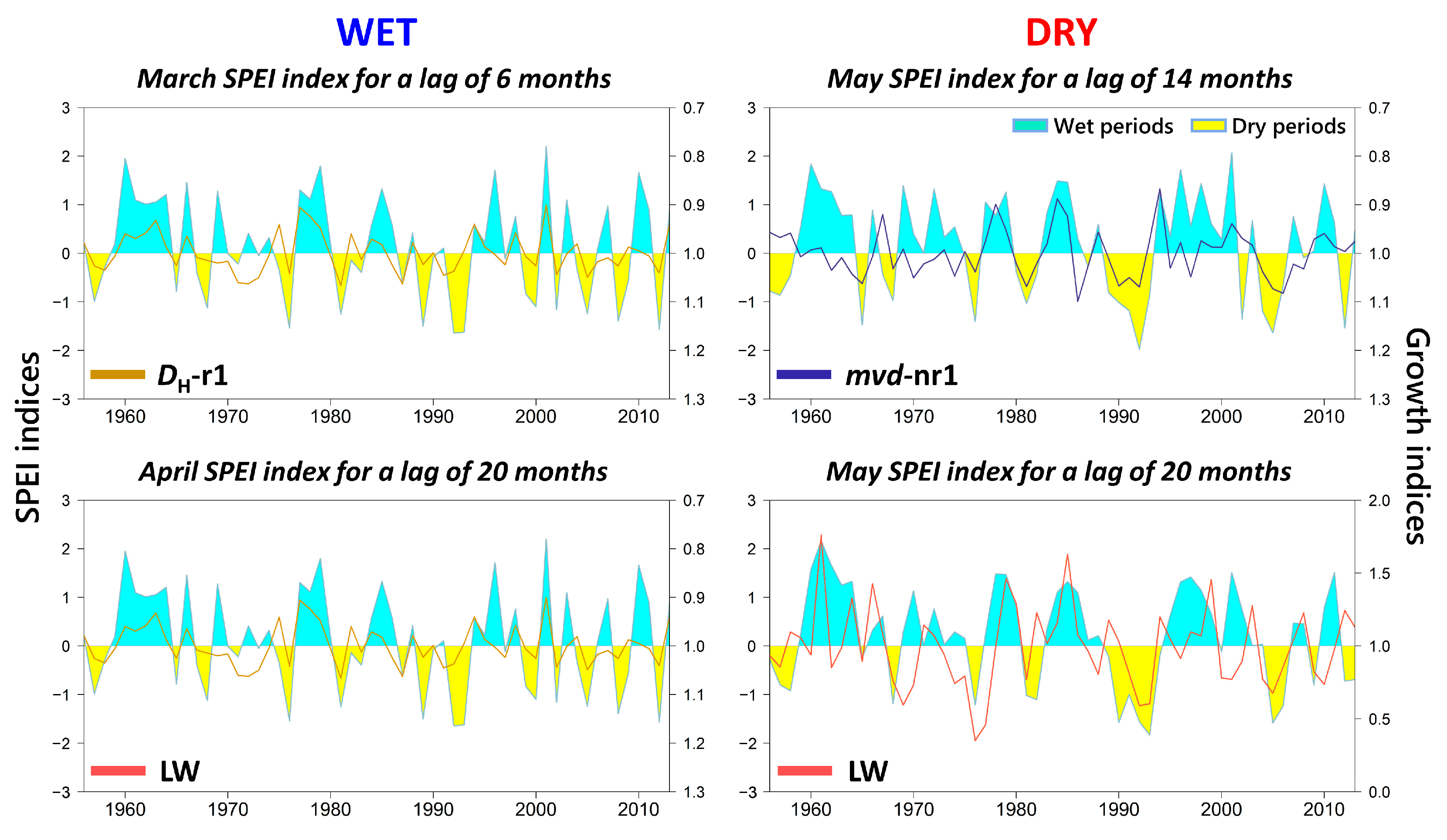
2. Results
3. Discussion
3.1. Considerations on the Selection of Variables and Site Characteristics
3.2. Effects of Climate on Wood Anatomy and Radial Increment
4. Materials and Methods
4.1. Study Area and Sites
4.2. Processing and Measurement of Tree Rings and Earlywood Vessels
4.3. Chronology Building and Assessment
4.4. Climate–Growth Relationships
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sánchez-de-Dios, R.; Benito-Garzón, M.; Sainz-Ollero, H. Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant Ecol. 2009, 204, 189–205. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis; IPCC: Cambridge, UK, 2021. [Google Scholar]
- Benito Garzón, M.; Sánchez de Dios, R.; Sainz Ollero, H. Effects of climate change on the distribution of Iberian tree species. Appl. Veg. Sci. 2008, 11, 169–178. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Franquesa, M.; Sanguesa-Barreda, G. Timing of Drought Triggers Distinct Growth Responses in Holm Oak: Implications to Predict Warming-Induced Forest Defoliation and Growth Decline. Forests 2015, 6, 1576–1597. [Google Scholar] [CrossRef] [Green Version]
- Rozas, V.; García-González, I. Too wet for oaks? Inter-tree competition and recent persistent wetness predispose oaks to rainfall-induced dieback in Atlantic rainy forest. Glob. Planet. Chang. 2012, 94–95, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Tyree, M.T. Xylem dysfunction in Quercus: Vessel sizes, tyloses, cavitation and seasonal changes in embolism. Tree Physiol. 1990, 6, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Aloni, R. Ecophysiological implications of vascular differentiation and plant evolution. Trees 2015, 29, 1–16. [Google Scholar] [CrossRef]
- Takahashi, S.; Okada, N.; Nobuchi, T. Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species. Ecol. Res. 2013, 28, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Pérez-de-Lis, G.; Rossi, S.; Vázquez-Ruiz, R.A.; Rozas, V.; García-González, I. Do changes in spring phenology affect earlywood vessels? Perspective from the xylogenesis monitoring of two sympatric ring-porous oaks. New Phytol. 2016, 209, 521–530. [Google Scholar] [CrossRef]
- Gallé, A.; Haldimann, P.; Feller, U. Photosynthetic performance and water relations in young pubescent oak (Quercus pubescens) trees during drought stress and recovery. New Phytol. 2007, 174, 799–810. [Google Scholar] [CrossRef]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef]
- González-González, B.D.; Rozas, V.; García-González, I. Earlywood vessels of the sub-Mediterranean oak Quercus pyrenaica have greater plasticity and sensitivity than those of the temperate Q. petraea at the Atlantic-Mediterranean boundary. Trees 2014, 28, 237–252. [Google Scholar] [CrossRef]
- Castagneri, D.; Regev, L.; Boaretto, E.; Carrer, M. Xylem anatomical traits reveal different strategies of two Mediterranean oaks to cope with drought and warming. Environ. Exp. Bot. 2017, 133, 128–138. [Google Scholar] [CrossRef]
- Souto-Herrero, M.; Rozas, V.; García-González, I. Earlywood vessels and latewood width explain the role of climate on wood formation of Quercus pyrenaica Willd. across the Atlantic-Mediterranean boundary in NW Iberia. For. Ecol. Manag. 2018, 425, 126–137. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Fonti, P.; Cherubini, P.; Martín-Benito, D.; Chaar, H.; Cañellas, I. Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. Tree Physiol. 2012, 32, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on growth and wood anatomical properties of Quercus faginea. IAWA J. 2004, 25, 185–204. [Google Scholar] [CrossRef]
- Fonti, P.; García-González, I. Earlywood vessel size of oak as a potential proxy for spring precipitation in mesic sites. J. Biogeogr. 2008, 35, 2249–2257. [Google Scholar] [CrossRef]
- García-González, I.; Eckstein, D. Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiol. 2003, 23, 497–504. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Camarero, J.J.; Azorin-Molina, C. Diverse responses of forest growth to drought time-scales in the Northern Hemisphere. Glob. Ecol. Biogeogr. 2014, 23, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- González-González, B.D.; Vázquez-Ruiz, R.A.; García-González, I. Effects of climate on earlywood vessel formation of Quercus robur and Q. pyrenaica at a site in the northwestern Iberian Peninsula. Can. J. For. Res. 2015, 45, 698–709. [Google Scholar] [CrossRef]
- Sperry, J.S.; Nichols, K.L.; Sullivan, J.E.M.; Eastlack, S.E. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of Northern Utah and Interior Alaska. Ecology 1994, 75, 1736–1752. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time-series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Hroš, M.; Vavrčík, H. Comparison of earlywood vessel variables in the wood of Quercus robur L. and Quercus petraea (Mattuschka) Liebl. growing at the same site. Dendrochronologia 2014, 32, 284–289. [Google Scholar] [CrossRef]
- Fonti, P.; García-González, I. Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol. 2004, 163, 77–86. [Google Scholar] [CrossRef]
- Kniesel, B.M.; Günther, B.; Roloff, A.; von Arx, G. Defining ecologically relevant vessel parameters in Quercus robur L. for use in dendroecology: A pointer year and recovery time case study in Central Germany. Trees 2015, 29, 1041–1051. [Google Scholar] [CrossRef]
- Souto-Herrero, M.; Rozas, V.; García-González, I. Chronologies of earlywood vessels and latewood width disentangle climate drivers of oak growth in a mild oceanic region. Dendrochronologia 2018, 51, 40–53. [Google Scholar] [CrossRef]
- Campelo, F.; Nabais, C.; Gutiérrez, E.; Freitas, H.; García-González, I. Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 2010, 24, 463–470. [Google Scholar] [CrossRef]
- Fonti, P.; Solomonoff, N.; García-González, I. Earlywood vessels of Castanea sativa record temperature before their formation. New Phytol. 2007, 173, 562–570. [Google Scholar] [CrossRef]
- García-González, I.; Souto-Herrero, M.; Campelo, F. Ring-porosity and earlywood vessels: A review on extracting environmental information through time. IAWA J. 2016, 37, 295–314. [Google Scholar] [CrossRef]
- Akhmetzyanov, L.; Buras, A.; Sass-Klaassen, U.; Den Ouden, J.; Mohren, F.; Groenendijk, P.; García-González, I. Multi-variable approach pinpoints origin of oak wood with higher precision. J. Biogeogr. 2019, 46, 1163–1177. [Google Scholar] [CrossRef] [Green Version]
- García-González, I.; Fonti, P. Selecting earlywood vessels to maximize their environmental signal. Tree Physiol. 2006, 26, 1289–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guada, G.; Vázquez-Ruiz, R.A.; García-González, I. Response patterns of xylem and leaf phenology to temperature at the southwestern distribution boundary of Quercus robur: A multi-spatial study. Agric. For. Meteorol. 2019, 269–270, 46–56. [Google Scholar] [CrossRef]
- Guada, G.; Vázquez-ruiz, R.A.; García-gonzález, I. Agricultural and Forest Meteorology Meteorological conditions control the cessation rather than the beginning of wood formation in a sub-Mediterranean ring-porous oak. Agric. For. Meteorol. 2020, 281, 107833. [Google Scholar] [CrossRef]
- Pérez-de-Lis, G.; Olano, J.M.; Rozas, V.; Rossi, S.; Vázquez-Ruiz, R.A.; García-González, I. Environmental conditions and vascular cambium regulate carbon allocation to xylem growth in deciduous oaks. Funct. Ecol. 2017, 31, 592–603. [Google Scholar] [CrossRef] [Green Version]
- Rozas, V.; Lamas, S.; García-González, I. Differential tree-growth responses to local and large-scale climatic variation in two Pinus and two Quercus species in northwest Spain. Ecoscience 2009, 16, 299–310. [Google Scholar] [CrossRef]
- Rozas, V.; García-González, I. Non-stationary influence of El Niño-Southern Oscillation and winter temperature on oak latewood growth in NW Iberian Peninsula. Int. J. Biometeorol. 2012, 56, 787–800. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Yoda, K.; Suzuki, H. Phenological comparison of the onset of vessel formation between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest. IAWA J. 1996, 17, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Cochard, H. Summer and winter embolism in oak: Impact on water relations. Ann. For. Sci. 1996, 53, 173–180. [Google Scholar] [CrossRef] [Green Version]
- El Zein, R.; Maillard, P.; Bréda, N.; Marchand, J.; Montpied, P.; Gérant, D. Seasonal changes of C and N non-structural compounds in the stem sapwood of adult sessile oak and beech trees. Tree Physiol. 2011, 31, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Barbaroux, C.; Bréda, N. Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiol. 2002, 22, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; White, D.A.; Ottenschlaeger, M.L.; Pinkard, E.A. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytol. 2013, 197, 862–872. [Google Scholar] [CrossRef]
- Sevanto, S.; McDowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Kreuzwieser, J.; Papadopoulou, E.; Rennenberg, H. Interaction of flooding with carbon metabolism of forest trees. Plant Biol. 2004, 6, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Vartapetian, B.B.; Jackson, M.B. Plant adaptations to anaerobic stress. Ann. Bot. 1997, 79, 3–20. [Google Scholar] [CrossRef]
- Visser, E.J.W.; Voensek, L.A.C.J.; Vartapetian, B.B.; Jackson, M.B. Flooding and Plant Growth. Ann. Bot. 2003, 91, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Pérez-de-Lis, G.; García-González, I.; Rozas, V.; Olano, J.M. Feedbacks between earlywood anatomy and non-structural carbohydrates affect spring phenology and wood production in ring-porous oaks. Biogeosciences 2016, 13, 5499–5510. [Google Scholar] [CrossRef] [Green Version]
- Amigo, J.; Izco, J.; Guitián, J.; Romero, M.I. Reinterpretación del robledal termófilo galaico-portugués: Rusco aculeati-Quercetum roboris. Lazaroa 1998, 19, 85–98. [Google Scholar]
- Gärtner, H.; Nievergelt, D. The core-microtome. A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 2010, 28, 85–92. [Google Scholar] [CrossRef]
- Fonti, P.; Broker, O.U.; Giudici, F. Relationship between ring shake incidence and earlywood vessel characteristics in chestnut wood. IAWA J. Suppl. 2002, 23, 287–298. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D. Research report evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- García-González, I.; Fonti, P. Ensuring a representative sample of earlywood vessels for dendroecological studies: An example from two ring-porous species. Trees 2008, 22, 237–244. [Google Scholar] [CrossRef]
- Cook, E.; Briffa, K.; Shiyatov, S.; Mazepa, V. Estimation of the Mean Chronology. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; pp. 123–132. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK; New York, NY, USA; San Francisco, CA, USA, 1976; p. 567. [Google Scholar]
- Mosteller, F.; Tukey, J.W. Data Analysis and Regression; Addison-Wesley: Reading, MA, USA, 1977. [Google Scholar]
- Rozas, V. Individual-based approach as a useful tool to disentangle the relative importance of tree age, size and inter-tree competition in dendroclimatic studies. iForest Biogeosci. For. 2015, 8, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2012, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-González, I.; Souto-Herrero, M. Earlywood Anatomy Highlights the Prevalent Role of Winter Conditions on Radial Growth of Oak at Its Distribution Boundary in NW Iberia. Plants 2023, 12, 1185. https://doi.org/10.3390/plants12051185
García-González I, Souto-Herrero M. Earlywood Anatomy Highlights the Prevalent Role of Winter Conditions on Radial Growth of Oak at Its Distribution Boundary in NW Iberia. Plants. 2023; 12(5):1185. https://doi.org/10.3390/plants12051185
Chicago/Turabian StyleGarcía-González, Ignacio, and Manuel Souto-Herrero. 2023. "Earlywood Anatomy Highlights the Prevalent Role of Winter Conditions on Radial Growth of Oak at Its Distribution Boundary in NW Iberia" Plants 12, no. 5: 1185. https://doi.org/10.3390/plants12051185