
Methoxyflavones from Black Ginger (Kaempferia parviflora Wall. ex Baker) and their Inhibitory Effect on Melanogenesis in B16F10 Mouse Melanoma Cells
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Structural Identification of Compounds
2.2. Effects of Methoxyflavones 1–6 on Viability of B16F10 Mouse Melanoma Cells
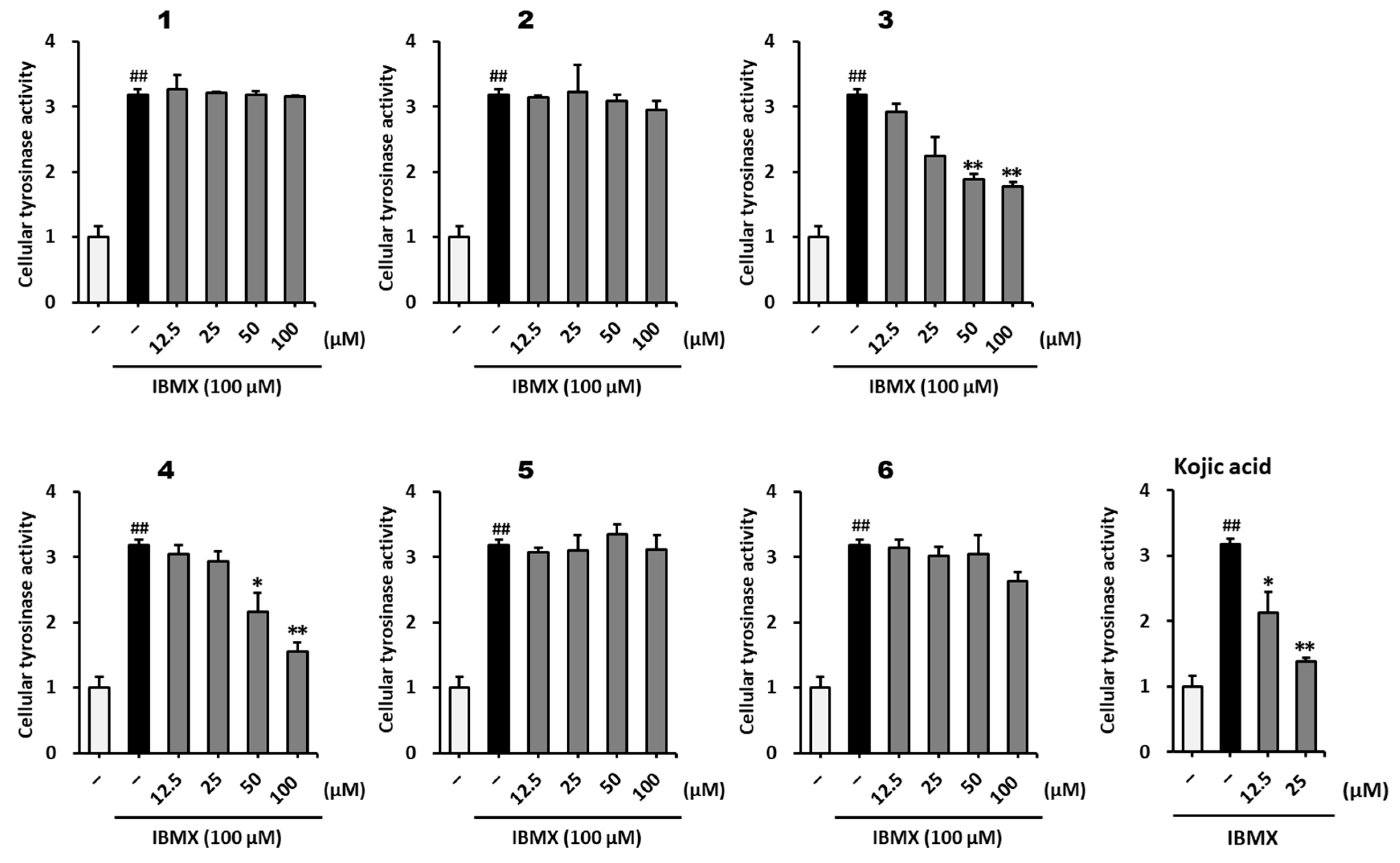
2.3. Inhibitory Effect of Methoxyflavones 1–6 on Melanogenesis in B16F10 Mouse Melanoma Cells
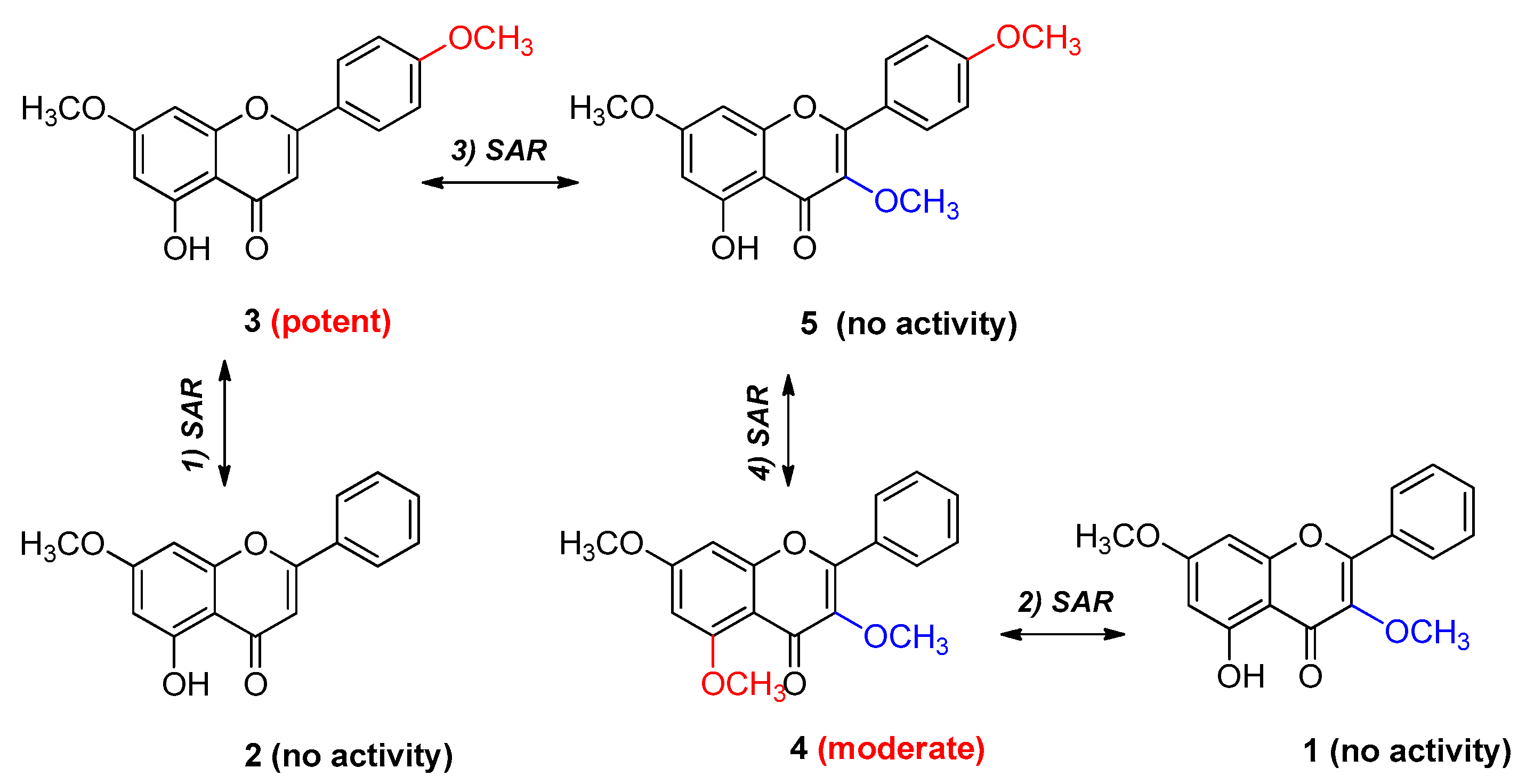
2.4. SAR Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Extraction and Separation of Methoxyflavones
3.3. Cell Culture
3.4. Cell Viability
3.5. Measurement of Cellular Tyrosinase Activity
3.6. Measurement of Cellular Melanin Content
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tucker, M.A. Melanoma epidemiology. Hematol. Oncol. Clin. N. Am. 2009, 23, 383–395. [Google Scholar] [CrossRef]
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Leutou, A.S.; Jeong, H.; Kim, D.; Seong, C.N.; Nam, S.J.; Lim, K.M. Anti-pigmentary effect of (-)-4-hydroxysattabacin from the marine-derived bacterium Bacillus sp. Mar. Drugs 2017, 15, 138. [Google Scholar]
- Kondo, T.; Hearing, V.J. Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Tief, K.; Hahne, M.; Schmidt, A.; Beermann, F. Tyrosinase, the key enzyme in melanin synthesis, is expressed in murine brain. Eur. J. Biochem. 1996, 241, 12–16. [Google Scholar] [CrossRef]
- Sim, M.O.; Ham, J.R.; Lee, M.K. Young leaves of reed (Phragmites communis) suppress melanogenesis and oxidative stress in B16F10 melanoma cells. Biomed. Pharmacother. 2017, 93, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 131, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Vachtenheim, J.; Borovanský, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef]
- Jimbow, K.; Obata, H.; Pathak, M.A.; Fitzpatrick, T.B. Mechanism of depigmentation by hydroquinone. J. Investig. Dermatol. 1974, 62, 436–449. [Google Scholar] [CrossRef] [Green Version]
- Pathak, M.A.; Fitzpatrick, T.B.; Kraus, E.W. Usefulness of retinoic acid in the treatment of melasma. J. Am. Acad. Dermatol. 1986, 15, 894–899. [Google Scholar] [CrossRef]
- Parvez, S.; Kang, M.; Chung, H.S.; Bae, H.S. Naturally occurring tyrosinase inhibitors: Mechanism and applications in skin health, cosmetics and agriculture industries. Phytother. Res. 2007, 21, 805–816. [Google Scholar] [CrossRef]
- Maeda, K.; Fukuda, M. Arbutin: Mechanism of its depigmenting action in human melanocyte culture. J. Pharmacol. Exp. Ther. 1996, 276, 765–769. [Google Scholar] [PubMed]
- Cabanes, J.; Chazarra, S.; García-Carmona, F. Kojic acid, a cosmetic skin whitening agent, is a slow-binding inhibitor of catecholase activity of tyrosinase. J. Pharm. Pharmacol. 1994, 46, 982–985. [Google Scholar] [CrossRef] [PubMed]
- McGregor, D. Hydroquinone: An evaluation of the human risks from its carcinogenic and mutagenic properties. Crit. Rev. Toxicol. 2007, 37, 887–914. [Google Scholar] [CrossRef] [PubMed]
- Lert-Amornpat, T.; Maketon, C.; Fungfuang, W. Effect of Kaempferia parviflora on sexual performance in streptozotocin-induced diabetic male rats. Andrologia 2017, 49, e127770. [Google Scholar] [CrossRef] [Green Version]
- Tewtrakul, S.; Subhadhirasakul, S.; Kummee, S. Anti-allergic activity of compounds from Kaempferia parviflora. J. Ethnopharmacol. 2008, 116, 191–193. [Google Scholar] [CrossRef]
- Kummee, S.; Tewtrakul, S.; Subhadhirasakul, S. Antimicrobial activity of the ethanol extract and compounds from the rhizomes of Kaempferia parviflora. J. Sci. Technol. 2008, 30, 463–466. [Google Scholar]
- Mekjaruskul, C.; Jay, M.; Sripanidkulchai, B. Pharmacokinetics, bioavailability, tissue distribution, excretion, and metabolite identification of methoxyflavones in Kaempferia parviflora extract in rats. Drug Metab. Dispos. 2012, 40, 2342–2353. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, S.; Kim, M.; Awa, R.; Kuwahara, H.; Kano, Y.; Kawada, T. Kaempferia parviflora extract increases energy consumption through activation of BAT in mice. Food Sci. Nutr. 2014, 2, 634–637. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Mohamed, T.A.; Essa, A.F.; Gawad, A.M.A.; Alqahtani, A.S.; Shahat, A.A.; Yoneyama, T.; Farrag, A.R.H.; Noji, M.; El-Seedi, H.R.; et al. Recent advances in Kaempferia phytochemistry and biological activity: A comprehensive review. Nutrients 2019, 11, 2396. [Google Scholar] [CrossRef] [Green Version]
- Saokaew, S.; Wilairat, P.; Raktanyakan, P.; Dilokthornsakul, P.; Dhippayom, T.; Kongkaew, C.; Sruamsiri, R.; Chuthaputti, A.; Chaiyakunapruk, N. Clinical effects of Krachaidum (Kaempferia parviflora): A systematic review. Evid. Based Complement. Altern. Med. 2017, 22, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Sutthanut, K.; Sripanidkulchai, B.; Yenjai, C.; Jay, M. Simultaneous identification, and quantitation of 11 flavonoid constituents in Kaempferia parviflora by gas chromatography. J. Chromatogr. 2007, 1143, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Sitthichai, P.; Chanpirom, S.; Maneerat, T.; Charoensup, R.; Tree-Udom, T.; Pintathong, P.; Laphookhieo, S.; Sripisut, T. Kaempferia parviflora rhizome extract as potential anti-acne ingredient. Molecules 2022, 27, 4401. [Google Scholar] [CrossRef]
- Chen, D.; Li, H.; Li, W.; Feng, S.; Deng, D. Kaempferia parviflora and its methoxyflavones: Chemistry and biological activities. Evid. based Complement. Altern. Med. 2018, 2018, 4057456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yenjai, C.; Wanich, S.; Pitchuanchom, S.; Sripanidkulchai, B. Structural modification of 5,7-dimethoxyflavone from Kaempferia parviflora and biological activities. Arch. Pharm. Res. 2009, 32, 1179–1184. [Google Scholar] [CrossRef]
- Jansakul, C.; Tachanaparuksa, K.; Mulvany, M.J.; Sukpondma, Y. Relaxant mechanisms of 3,5,7,3’,4’-pentamethoxyflavone on isolated human cavernosum. Eur. J. Clin. Pharmacol. 2012, 691, 235–244. [Google Scholar] [CrossRef]
- Tewtrakul, S.; Subhadhirasakul, S. Effects of compounds from Kaempferia parviflora on nitric oxide, prostaglandin E2 and tumor necrosis factor-alpha productions in RAW264.7 macrophage cells. J. Ethnopharmacol. 2008, 120, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.S.; So, H.M.; Kim, S.; Kim, J.K.; Kim, J.; Kang, D.; Ahn, M.; Ko, Y.; Kim, K.H. Comparative evaluation of bioactive phytochemicals in Spinacia oleracea cultivated under greenhouse and open field conditions. Arch. Pharm. Res. 2022, 45, 795–805. [Google Scholar] [CrossRef]
- Cho, H.; Kim, K.H.; Han, S.H.; Kim, H.; Cho, I.; Lee, S. Structure determination of heishuixiecaoline A from Valeriana fauriei and its content from different cultivated regions by HPLC/PDA Analysis. Nat. Prod. Sci. 2022, 28, 181–186. [Google Scholar] [CrossRef]
- Yu, J.S.; Jeong, S.Y.; Li, C.; Oh, T.; Kwon, M.; Ahn, J.S.; Ko, S.; Ko, Y.; Cao, S.; Kim, K.H. New phenalenone derivatives from the Hawaiian volcanic soil-associated fungus Penicillium herquei FT729 and their inhibitory effects on indoleamine 2,3-dioxygenase 1 (IDO1). Arch. Pharm. Res. 2022, 45, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Lee, B.S.; Yu, J.S.; Kang, H.; Yoo, M.J.; Yi, S.A.; Han, J.; Kim, S.; Kim, J.K.; Kim, J.; et al. Identification of anti-adipogenic withanolides from the roots of Indian ginseng (Withania somnifera). J. Ginseng. Res. 2022, 46, 357–366. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, J.K.; Yu, J.S.; Jeong, S.Y.; Choi, J.H.; Kim, J.; Ko, Y.; Kim, S.; Kim, K.H. Ginkwanghols A and B, osteogenic coumaric acid-aliphatic alcohol hybrids from the leaves of Ginkgo biloba. Arch. Pharm. Res. 2021, 44, 514–524. [Google Scholar] [CrossRef]
- Lee, S.; Jang, T.; Kim, K.H.; Kang, K.S. Improvement of damage in human dermal fibroblasts by 3,5,7-trimethoxyflavone from black ginger (Kaempferia parviflora). Antioxidants 2022, 11, 425. [Google Scholar] [CrossRef]
- Ghani, N.A.; Ahmat, N.; Ismail, N.H.; Zakaria, I. Flavonoid constituents from the stem bark of polyalthia cauliflora var. Cauliflora. Aust. J. Basic Appl. Sci. 2011, 5, 154–158. [Google Scholar]
- Rosandy, A.R.; Din, L.B.; Yaacob, W.A.; Yusoff, N.I.; Sahidin, I.; Latip, J.; Nataqain, S.; Noor, N.M. Isolation and characterization of compounds from the stem bark of Uvaria rufa (Annonaceae). Malays. J. Anal. Sci. 2013, 17, 50–58. [Google Scholar]
- Dao, T.T.; Chi, Y.S.; Kim, J.; Kim, H.P.; Kim, S.; Park, H. Synthesis and PGE 2 inhibitory activity of 5,7-dihydroxyflavones and their o-methylated flavone analogs. Arch. Pharm. Res. 2003, 26, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Buschi, C.A.; Pomilio, A.B.; Gros, E.G. New methylated flavones from Gomphrena martiana. Phytochemistry 1980, 19, 903–904. [Google Scholar] [CrossRef]
- Nguyen, T.K.P.; Nguyen, K.P.P.; Kamounah, F.S.; Zhang, W.; Hansen, P.E. NMR of a series of novel hydroxyflavothiones. Magn. Reason. Chem. 2009, 47, 1043–1054. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Byun, E.B.; Song, H.Y.; Mushtaq, S.; Kim, H.M.; Kang, J.A.; Yang, M.S.; Sung, N.Y.; Jang, B.S.; Byun, E.H. Gamma-irradiated luteolin inhibits 3-isobutyl-1-methylxanthine-induced melanogenesis through the regulation of CREB/MITF, PI3K/Akt, and ERK pathways in B16BL6 melanoma cells. J. Med. Food 2017, 20, 812–819. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative stress in the pathogenesis of skin disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [Green Version]
- Fistarol, S.K.; Itin, P.H. Disorders of pigmentation. J. Dtsch. Dermatol. Ges. 2010, 8, 187–202. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.; Swope, V.B.; Pallas, J.; Krug, K.; Nordlund, J.J. Mitogenic, melanogenic, and cAMP responses of cultured neonatal human melanocytes to commonly used mitogens. J. Cell. Physiol. 1992, 150, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Y.; Chen, X. Computational identification of bioactive natural products by structure activity relationship. J. Mol. Graph. Model. 2010, 29, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Perkins, R.; Fang, H.; Tong, W.; Welsh, W.J. Quantitative structure-activity relationship methods: Perspectives on drug discovery and toxicology. Environ. Toxicol. Chem. 2003, 22, 1666–1679. [Google Scholar] [CrossRef] [PubMed]
- Rahmana, M.M.; Sahaa, T.; Islama, K.J.; Sumana, R.H.; Biswasa, S.; Rahata, E.U.; Hossena, M.R.; Islama, R.; Hossaina, M.N.; Mamunb, A.A.; et al. Virtual screening, molecular dynamics and structure–activity relationship studies to identify potent approved drugs for Covid-19 treatment. J. Biomol. Struct. Dyn. 2021, 39, 6231–6241. [Google Scholar] [CrossRef]
- Shamsudin, N.F.; Ahmed, Q.U.; Mahmood, S.; Shah, S.A.A.; Khatib, A.; Mukhtar, S.; Alsharif, M.A.; Parveen, H.; Zakaria, Z.A. Antibacterial effects of flavonoids and their structure-activity relationship study: A comparative interpretation. Molecules 2022, 27, 1149. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure–activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; Kang, S.; Lee, I.; Lee, J.; Jung, H.; Choi, C. Antioxidant and chemosensitizing effects of flavonoids with hydroxy and/or methoxy groups and structure-activity relationship. J. Pharm. Pharmaceut. Sci. 2007, 10, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Eghbali-Feriz, S.; Taleghani, A.; Al-Najjar, H.; Emami, S.; Rahimi, H.; Asili, J.; Tayarani-Najaran, Z. Anti-melanogenesis and anti-tyrosinase properties of Pistacia atlantica subsp. mutica extracts on B16F10 murine melanoma cells. Res. Pharm. Sci. 2018, 13, 533–545. [Google Scholar]
- Hosoi, J.; Abe, E.; Suda, T.; Kuroki, T. Regulation of melanin synthesis of B16 mouse melanoma cells by 1α, 25-dihydroxyvitamin D3 and retinoic acid. Cancer Res. 1985, 45, 1474–1478. [Google Scholar] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, C.; Lee, S.; Yoo, M.J.; Lee, B.S.; Jang, Y.S.; Kim, H.K.; Lee, S.; Bae, H.Y.; Kim, K.H. Methoxyflavones from Black Ginger (Kaempferia parviflora Wall. ex Baker) and their Inhibitory Effect on Melanogenesis in B16F10 Mouse Melanoma Cells. Plants 2023, 12, 1183. https://doi.org/10.3390/plants12051183
Huo C, Lee S, Yoo MJ, Lee BS, Jang YS, Kim HK, Lee S, Bae HY, Kim KH. Methoxyflavones from Black Ginger (Kaempferia parviflora Wall. ex Baker) and their Inhibitory Effect on Melanogenesis in B16F10 Mouse Melanoma Cells. Plants. 2023; 12(5):1183. https://doi.org/10.3390/plants12051183
Chicago/Turabian StyleHuo, Chen, Sullim Lee, Min Jeong Yoo, Bum Soo Lee, Yoon Seo Jang, Ho Kyong Kim, Seulah Lee, Han Yong Bae, and Ki Hyun Kim. 2023. "Methoxyflavones from Black Ginger (Kaempferia parviflora Wall. ex Baker) and their Inhibitory Effect on Melanogenesis in B16F10 Mouse Melanoma Cells" Plants 12, no. 5: 1183. https://doi.org/10.3390/plants12051183