


Silver Inhibits Lemna minor Growth at High Initial Frond Densities

Abstract
:1. Introduction
2. Results
2.1. Experiment 1: Optimal Nutrient Concentration
2.2. Experiment 2: Effects of Silver Nanoparticle Concentrations
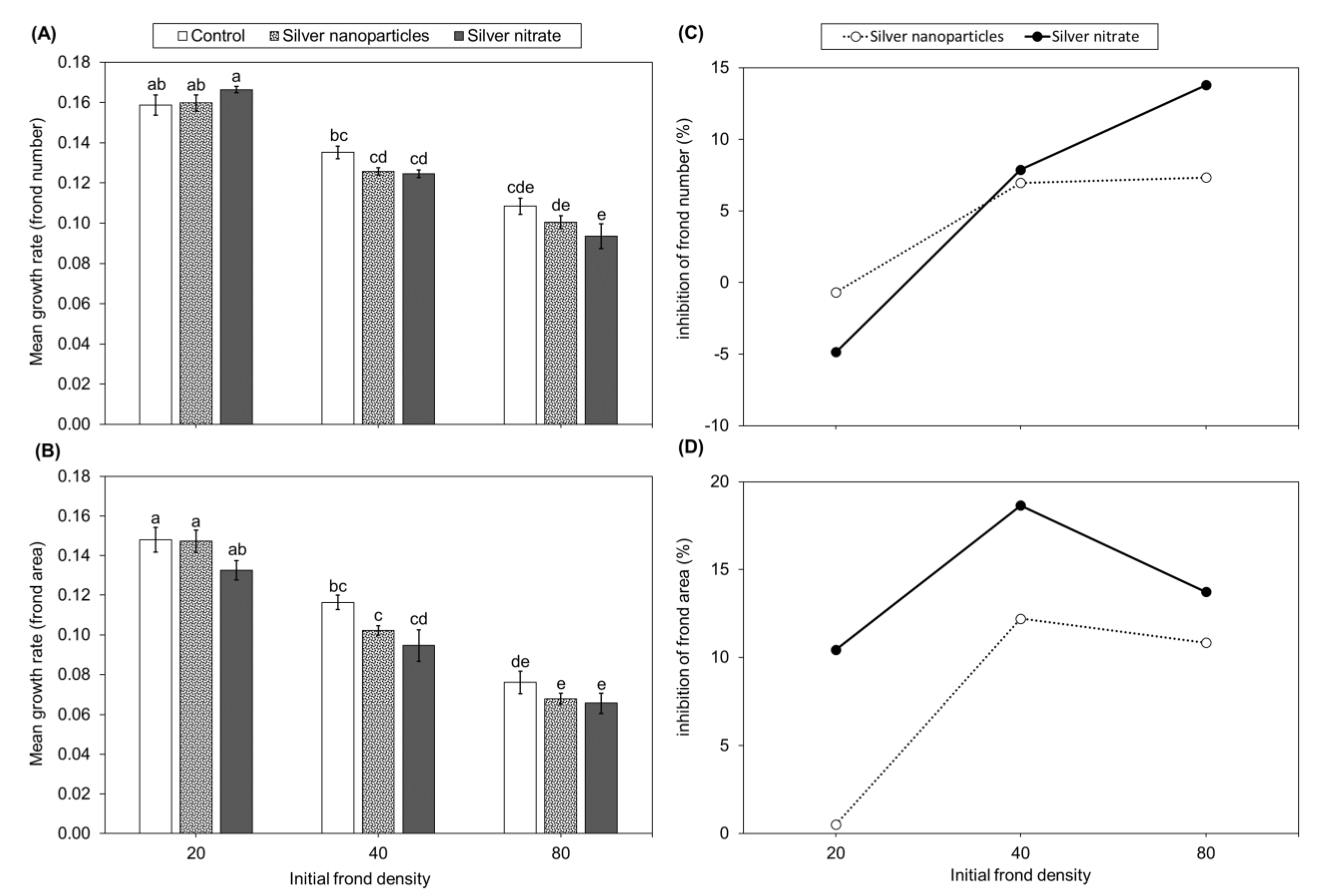
2.3. Experiment 3: Plant Growth Response of Lemna minor to Different Forms of Silver
3. Discussion
4. Materials and Methods
4.1. Experiment 1: Optimal Nutrient Concentration
4.2. Experiment 2: Effects of Silver Nanoparticle Concentrations
4.3. Experiment 3: Plant Growth Response of Lemna minor to Different Forms of Silver
4.4. Calculations and Statistical Analysis
- μi–j = average specific growth rate from time i to j
- Ni = measurement in treatment or control at time i
- Nj = measurement in treatment or control at time j
- t = time from i to j
- μC = mean growth rate of control
- μT = mean growth rate of treatment
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jain, D.; Daima, H.K.; Kachhwaha, S.; Kothari, S.L. Synthesis of plant-mediated silver nanoparticles using papaya fruit extract and evaluation of their anti microbial activities. Dig. J. Nanomater. Biostruct. 2009, 4, 557–563. [Google Scholar]
- Zhang, X.F.; Liu, Z.G.; Shen, W.; Gurunathan, S. Silver nanoparticles: Synthesis, characterization, properties, applications, and therapeutic approaches. Int. J. Mol. Sci. 2016, 4, 1534. [Google Scholar] [CrossRef]
- Choi, O.; Deng, K.K.; Kim, N.-J.; Ross, L.; Surampalli, R.Y.; Hu, Z. The inhibitory effects of silver nanoparticles, silver ions, and silver chloride colloids on microbial growth. Water Res. 2008, 4, 3066–3074. [Google Scholar] [CrossRef]
- Choi, O.K.; Hu, Z.Q. Nitrification inhibition by silver nanoparticles. Water Sci. Technol. 2009, 4, 1699–1702. [Google Scholar] [CrossRef]
- Benn, T.M.; Westerhoff, P. Nanoparticle silver released into water from commercially available sock fabrics. Environ. Sci. Technol. 2008, 4, 4133–4139. [Google Scholar] [CrossRef]
- Geranio, L.; Heuberger, M.; Nowack, B. The behavior of silver nanotextiles during washing. Environ. Sci. Technol. 2009, 4, 8113–8118. [Google Scholar] [CrossRef] [Green Version]
- Mueller, N.C.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 4, 4447–4453. [Google Scholar] [CrossRef]
- Blaser, S.A.; Scheringer, M.; MacLeod, M.; Hungerbühler, K. Estimation of cumulative aquatic exposure and risk due to silver: Contribution of nano-functionalized plastics and textiles. Sci. Total Environ. 2008, 4, 396–409. [Google Scholar] [CrossRef]
- Kaegi, R.; Sinnet, B.; Zuleeg, S.; Hagendorfer, H.; Mueller, E.; Vonbank, R.; Boller, M.; Burkhardt, M. Release of silver nanoparticles from outdoor facades. Environ. Pollut. 2010, 4, 2900–2905. [Google Scholar] [CrossRef]
- Kaegi, R.; Voegelin, A.; Ort, C.; Sinnet, B.; Thalmann, B.; Krismer, J.; Hagendorfer, H.; Elumelu, M.; Mueller, E. Fate and transformation of silver nanoparticles in urban wastewater systems. Water Res. 2013, 4, 3866–3877. [Google Scholar] [CrossRef]
- Lee, W.-M.; Kwak, J.I.; An, Y.-J. Effect of silver nanoparticles in crop plants Phaseolus radiatus and Sorghum bicolor: Media effect on phytotoxicity. Chemosphere 2012, 4, 491–499. [Google Scholar] [CrossRef]
- Oukarroum, A.; Barhoumi, L.; Pirastru, L.; Dewez, D. Silver nanoparticle toxicity effect on growth and cellular viability of the aquatic plant Lemna gibba. Environ. Toxicol. Chem. 2013, 4, 902–907. [Google Scholar] [CrossRef]
- Thwala, M.; Klaine, S.; Musee, N. Exposure Media and Nanoparticle Size Influence on the Fate, Bioaccumulation, and Toxicity of Silver Nanoparticles to Higher Plant Salvinia minima. Molecules 2021, 4, 2305. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Colman, B.P.; McGill, B.M.; Wright, J.P.; Bernhardt, E.S. Effects of silver nanoparticle exposure on germination and early growth of eleven wetland plants. PLoS ONE 2012, 4, e47674. [Google Scholar] [CrossRef] [Green Version]
- Naumann, B.; Eberius, M.; Appenroth, K.-J. Growth rate based dose–response relationships and EC-values of ten heavy metals using the duckweed growth inhibition test (ISO 20079) with Lemna minor L. clone St. J. Plant Physiol. 2007, 4, 1656–1664. [Google Scholar] [CrossRef]
- Daud, M.K.; Shafaqaat, A.; Abbas, Z.; Zaheer, I.E.; Riaz, M.A.; Malik, A.; Hussain, A.; Rizwan, M.; Zia-ur-Rehman, M.; Zhu, S.J. Potential of Duckweed (Lemna minor) for the Phytoremediation of Landfill Leachate. J. Chem. 2018, 2018, 3951540. [Google Scholar] [CrossRef] [Green Version]
- Ekperusi, A.O.; Sikoki, F.D.; Nwachukwu, E.O. Application of common duckweed (Lemna minor) in phytoremediation of chemicals in the environment: State and future perspective. Chemosphere 2019, 4, 285–309. [Google Scholar] [CrossRef]
- Chaudhary, E.; Sharma, P. Chromium and cadmium removal from wastewater using duckweed—Lemna gibba L. and ultrastructural deformation due to metal toxicity. Int. J. Phytoremediat. 2019, 4, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Landesman, L.; Fedler, C.; Duan, R. Plant Nutrient Phytoremediation Using Duckweed. In Eutrophication: Causes, Consequences and Control; Ansart, A.A., Gill, S.S., Lanza, G.R., Rast, W., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 341–354. [Google Scholar]
- Lalau, C.M.; Simioni, C.; Vicentini, D.S.; Ouriques, L.C.; Mohedano, R.A.; Puerari, R.C.; Matias, W.G. Toxicological effects of AgNPs on duckweed (Landoltia punctata). Sci. Total Environ. 2020, 4, 136318. [Google Scholar] [CrossRef] [PubMed]
- Thwala, M.; Musee, N.; Sikhwivhilu, L.; Wepener, V. The oxidative toxicity of Ag and ZnO nanoparticles towards the aquatic plant Spirodela punctata and the role of testing media parameters. Environ. Sci. Process Impacts 2013, 4, 1830–1843. [Google Scholar] [CrossRef]
- Minogiannis, P.; Valenti, M.; Kati, V.; Kalantzi, O.-I.; Biskos, G. Toxicity of pure silver nanoparticles produced by spark ablation on the aquatic plant Lemna minor. J. Aerosol. Sci. 2019, 4, 17–21. [Google Scholar] [CrossRef]
- Üçüncü, E.; Özkan, A.D.; Kurşungöz, C.; Ülger, Z.E.; Ölmez, T.T.; Tekinay, T.; Ortaç, B.; Tunca, E. Effects of laser ablated silver nanoparticles on Lemna minor. Chemosphere 2014, 4, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Gubbins, E.J.; Batty, L.C.; Lead, J.R. Phytotoxicity of silver nanoparticles to Lemna minor L. Environ. Pollut. 2011, 4, 1551–1559. [Google Scholar] [CrossRef]
- Pereira, S.P.P.; Jesus, F.; Aguiar, S.; de Oliveira, R.; Fernandes, M.; Ranville, J.; Nogueira, A.J.A. Phytotoxicity of silver nanoparticles to Lemna minor: Surface coating and exposure period-related effects. Sci. Total Environ. 2018, 4, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.R.; Corrêa, T.Z.; Bruni, A.T.; da Veiga, M.A. The effects of solubility of silver nanoparticles, accumulation, and toxicity to the aquatic plant Lemna minor. Environ. Sci. Pollut. Res. 2021, 4, 16720–16733. [Google Scholar] [CrossRef]
- Jiang, H.-S.; Li, M.; Chang, F.-Y.; Li, W.; Yin, L.-Y. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 2012, 4, 1880–1886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-S.; Qiu, X.-N.; Li, G.-B.; Li, W.; Yin, L.-Y. Silver nanoparticles induced accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 2014, 4, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Li, P.; Huang, Q.; Zhang, H. The different response mechanisms of Wolffia globosa: Light-induced silver nanoparticle toxicity. Aquat. Toxicol. 2016, 4, 97–105. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development. OECD Guidelines for the Testing of Chemicals, Section 2. Effects on Biotic Systems, Test No. 221: Lemna sp. Growth Inhibition Test; OECD Publishing: Paris, France, 2006. [Google Scholar]
- Ding, Y.; Bai, X.; Ye, Z.; Gong, D.; Cao, J.; Hua, Z. Humic acid regulation of the environmental behavior and phytotoxicity of silver nanoparticles to Lemna minor. Environ. Sci. Nano 2019, 4, 3712–3722. [Google Scholar] [CrossRef]
- Kim, E.; Kim, S.-H.; Kim, H.-C.; Lee, S.G.; Lee, S.J.; Jeong, S.W. Growth inhibition of aquatic plant caused by silver and titanium oxide nanoparticles. Toxicol. Environ Health Sci. 2011, 4, 1–6. [Google Scholar] [CrossRef]
- Frédéric, M.; Samir, L.; Louise, M.; Abdelkrim, A. Comprehensive modeling of mat density effect on duckweed (Lemna minor) growth under controlled eutrophication. Water Res. 2006, 4, 2901–2910. [Google Scholar] [CrossRef]
- Walsh, É.; Coughlan, N.E.; O’Brien, S.; Jansen, M.A.K.; Kuehnhold, H. Density Dependence Influences the Efficacy of Wastewater Remediation by Lemna minor. Plants 2021, 4, 1366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.M.; Jin, Y.; Yao, S.M.; Lei, N.F.; Chen, J.S.; Zhang, Q.; Yu, F.H. Growth and morphological responses of duckweed to clonal fragmentation, nutrient availability, and population density. Front. Plant. Sci. 2020, 4, 618. [Google Scholar] [CrossRef]
- Iqbal, J.; Javed, A.; Javed, H. Effect of Initial Plant Density on Growth and Nutrients Removal Efficiency of Duckweed (Lemna minor) from Leachate. Asian J. Environ. Ecol. 2021, 45–51. [Google Scholar] [CrossRef]
- Demirezen, D.; Aksoy, A.; Uruç, K. Effect of population density on growth, biomass and nickel accumulation capacity of Lemna gibba (Lemnaceae). Chemosphere 2007, 4, 553–557. [Google Scholar] [CrossRef]
- Dosnon-Olette, R.; Couderchet, M.; El Arfaoui, A.; Stéphanie, S.; Philippe, E. Influence of initial pesticide concentrations and plant population density on dimethomorph toxicity and removal by two duckweed species. Sci. Total Environ. 2010, 4, 2254–2259. [Google Scholar] [CrossRef] [PubMed]
- Färber, E.; Kandeler, R. Significance of Calcium Ions in the Overcrowding Effect in Spirodela polyrrhiza P 143. J. Plant Physiol. 1989, 4, 94–98. [Google Scholar] [CrossRef]
- Färber, E.; Kandeler, R. Phytochrome effect on the ethylene production after overcrowding in Spirodela (Lemnaceae). Phyton 1990, 30, 89–95. [Google Scholar]
- Stegemeier, J.P.; Colman, B.P.; Schwab, F.; Wiesner, M.R.; Lowry, G.V. Uptake and distribution of silver in the aquatic plant Landoltia punctata (duckweed) exposed to silver and silver sulfide nanoparticles. Environ. Sci. Technol. 2017, 4, 4936–4943. [Google Scholar] [CrossRef] [PubMed]
- Kufel, L.; Strzałek, M.; Przetakiewicz, A. Plant response to overcrowding—Lemna minor example. Acta Oecologica 2018, 4, 73–80. [Google Scholar] [CrossRef]
- Iannelli, M.A.; Bellini, A.; Venditti, I.; Casentini, B.; Battocchio, C.; Scalici, M.; Ceschin, S. Differential phytotoxic effect of silver nitrate (AgNO3) and bifunctionalized silver nanoparticles (AgNPs-Cit-L-Cys) on Lemna plants (duckweeds). Aquat. Toxicol. 2022, 4, 106260. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant response to silver nanoparticles: A critical review. Crit. Rev. Biotechnol. 2022, 4, 973–990. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.H.; Evert, R.F.; Eichorn, S.E. Biology of Plants, 5th ed.; Worth Publishers: New York, NY, USA, 1992. [Google Scholar]
- Reimer, D.N. Introduction to Freshwater Vegetation; AVI Publishing Company, Inc.: Mokelumne Hill, CA, USA, 1984. [Google Scholar]
- Yatskievych, G. Steyermark’s Flora of Missouri, Revised ed.; Missouri Department of Conservation: Jefferson City, MO, USA, 1999; Volume 1.
- Landolt, E. Lemnaceae. In Flora of North America; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2000; Volume 22. [Google Scholar]
- Zayed, A.; Gowthaman, S.; Terry, N. Phytoaccumulation of trace elements by wetland plants: I. Duckweed J. Environ. Qual. 1998, 4, 715–721. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 4, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | AgNP (µg/L) | Size (nm) | Effect | References |
|---|---|---|---|---|
| Landoltia punctata | 10 | 19–27 | Decreased growth rate, frond area, and root size | [20] |
| 10 | 40–60 | Increased reactive oxygen species (ROS) | [21] | |
| Lemna gibba | 10 | 50 | Decreased frond number | [12] |
| 100 | Decreased cell viability | |||
| Lemna minor | 5 | 10–80 | Decreased frond number | [22] |
| 8 | 5–20 | Decreased frond number and dry weight | [23] | |
| 10 | 20 | Decreased dry weight | [24] | |
| 20 | Decreased frond number | |||
| 50 | 79 | Decreased growth rate | [25] | |
| 800 | Decreased fronds per colony | |||
| 2000 | Chlorosis | |||
| 10,000 | 30 | Decreased growth rate | [26] | |
| Spirodela polyrrhiza | 500 | 6 | Chlorosis | [27] |
| 1000 | Root abscission | |||
| 5000 | Decreased dry weight | |||
| 10,000 | 20 | Increased ROS | [28] | |
| Wolffia globosa | 1000 | 10 | Decreased photosynthetic activity | [29] |
| 10,000 | Decreased photosynthetic pigments |
| Plant Species | Initial Frond Number | Growth Measurements | References | ||
|---|---|---|---|---|---|
| Frond Number | Dry Weight | Frond Area | |||
| Landoltia punctata | not given | x | x | [20] | |
| Lemna gibba | 15 | x | [12] | ||
| Lemna minor | 10 | x | [25] | ||
| 15–20 | x | x | [24] | ||
| 21 | x | x | [23] | ||
| not given | x | [31] | |||
| not given | x | [22] | |||
| Lemna aequinoctialis (formerly Lemna paucicostata) | 5 | x | [32] | ||
| Spirodela polyrrhiza | not given | x | x | [27] | |
| Hoagland Nutrient Solution | Frond Number | Dry Weight (mg) | Frond Area (cm2) | |||
|---|---|---|---|---|---|---|
| 25% | 471 ± 12 | a | 121 ± 3 | a | 27 ± 2 | a |
| 2.5% | 307 ± 11 | b | 83 ± 3 | ab | 16 ± 0.7 | ab |
| 1% | 193 ± 2 | c | 55 ± 5 | bc | 8 ± 0.3 | b |
| 0.5% | 148 ± 2 | d | 43 ± 3 | cd | 6 ± 0.3 | c |
| 0.3% | 132 ± 4 | d | 37 ± 2 | cd | 5 ± 0.2 | c |
| 0.25% | 115 ± 2 | e | 32 ± 4 | d | 5 ± 0.4 | c |
| AgNP (μg/L) | Dry Weight (mg) | Frond Number | Frond Area (cm2) | % Inhibition of Frond Number | % Inhibition of Frond Area | |||
|---|---|---|---|---|---|---|---|---|
| 0 | 78 ± 2 | a | 208 ± 13 | ab | 7 ± 0.4 | a | 0 | 0 |
| 60 | 72 ± 0.6 | b | 220 ± 10 | ab | 6 ± 0.2 | ab | −4 | 6 |
| 125 | 69 ± 2 | b | 236 ± 14 | a | 6 ± 0.3 | a | −8 | 9 |
| 250 | 67 ± 0.2 | bc | 200 ± 7 | ab | 6 ± 0.4 | ab | 2 | 12 |
| 500 | 61 ± 1 | c | 169 ± 9 | bc | 4 ± 0.4 | b | 12 | 30 |
| 1000 | 53 ± 1 | d | 111 ± 10 | c | 2 ± 0.3 | c | 38 | 75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, I.T.; Heiman, J.A.; Lydy, V.R.; Kissoon, L.T. Silver Inhibits Lemna minor Growth at High Initial Frond Densities. Plants 2023, 12, 1104. https://doi.org/10.3390/plants12051104
Tran IT, Heiman JA, Lydy VR, Kissoon LT. Silver Inhibits Lemna minor Growth at High Initial Frond Densities. Plants. 2023; 12(5):1104. https://doi.org/10.3390/plants12051104
Chicago/Turabian StyleTran, Indigo T., Jordan A. Heiman, Victoria R. Lydy, and La Toya Kissoon. 2023. "Silver Inhibits Lemna minor Growth at High Initial Frond Densities" Plants 12, no. 5: 1104. https://doi.org/10.3390/plants12051104