
Comparison of Soil Bacterial Communities under Canopies of Pinus tabulaeformis and Populus euramericana in a Reclaimed Waste Dump
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
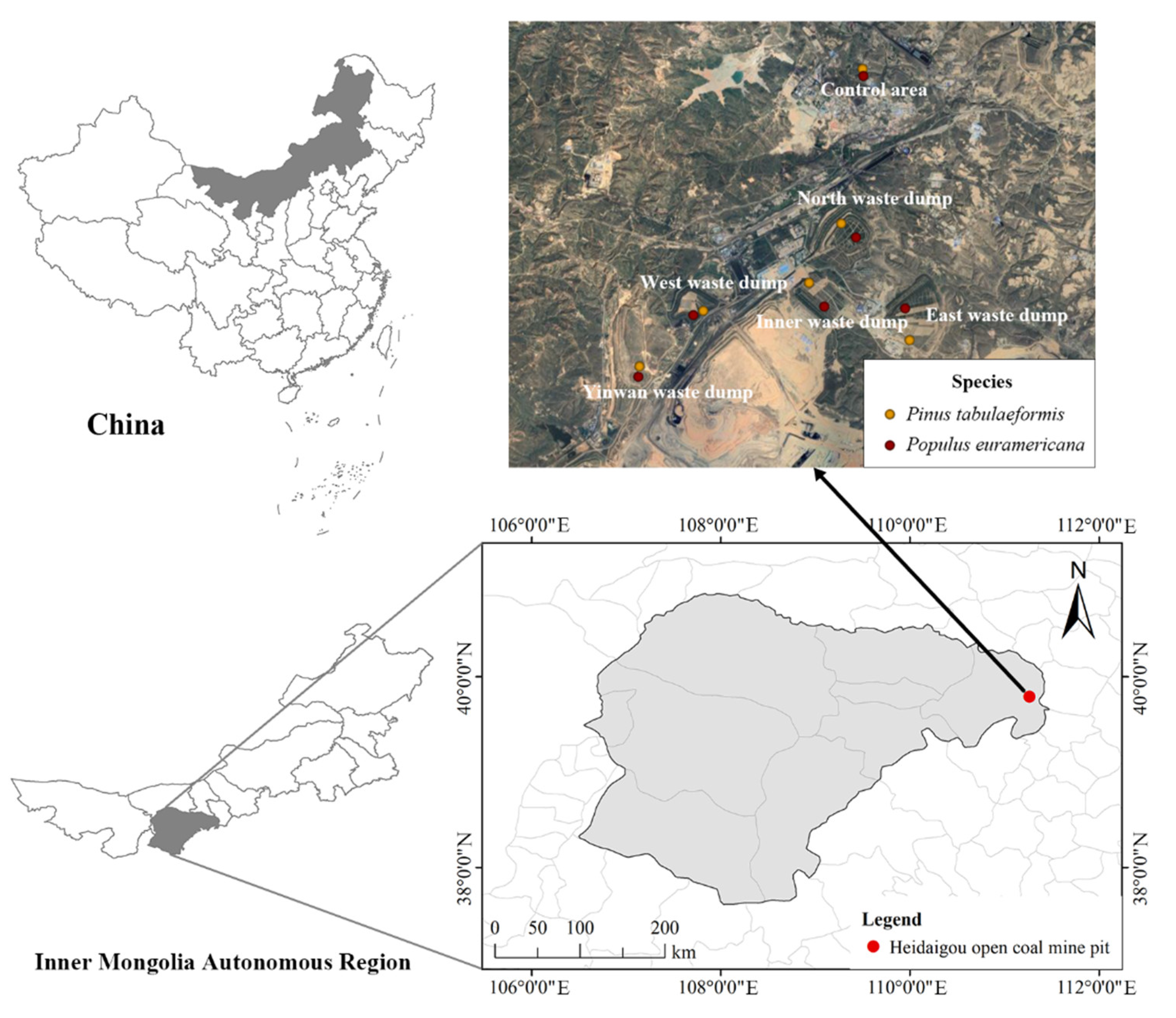
2.1. Overview of the Study Area
2.2. Sample Collection and Analysis
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Molecular Ecological Network Analysis
2.5. Data Analysis and Processing
3. Results
3.1. Physical and Chemical Properties and Enzyme Activity Characteristics of Soil under Different Reclamation Tree Species and Different Years
3.2. Characteristics of Bacterial Community Structure and Diversity in the Reclaimed Soil of P. tabulaeformis and P. euramericana
3.2.1. Characteristics of the Bacterial Community Structure in the Reclaimed Soil of P. tabulaeformis and P. euramericana
3.2.2. Diversity of the Bacterial Community in the Reclaimed Soil of P. tabulaeformis and P. euramericana
3.2.3. Beta Diversity of the Bacterial Community in the Reclaimed Soil of P. tabulaeformis and P. euramericana
3.3. Analysis of Bacterial Molecular Ecological Network in the Reclaimed Soil of P. tabulaeformis and P. euramericana
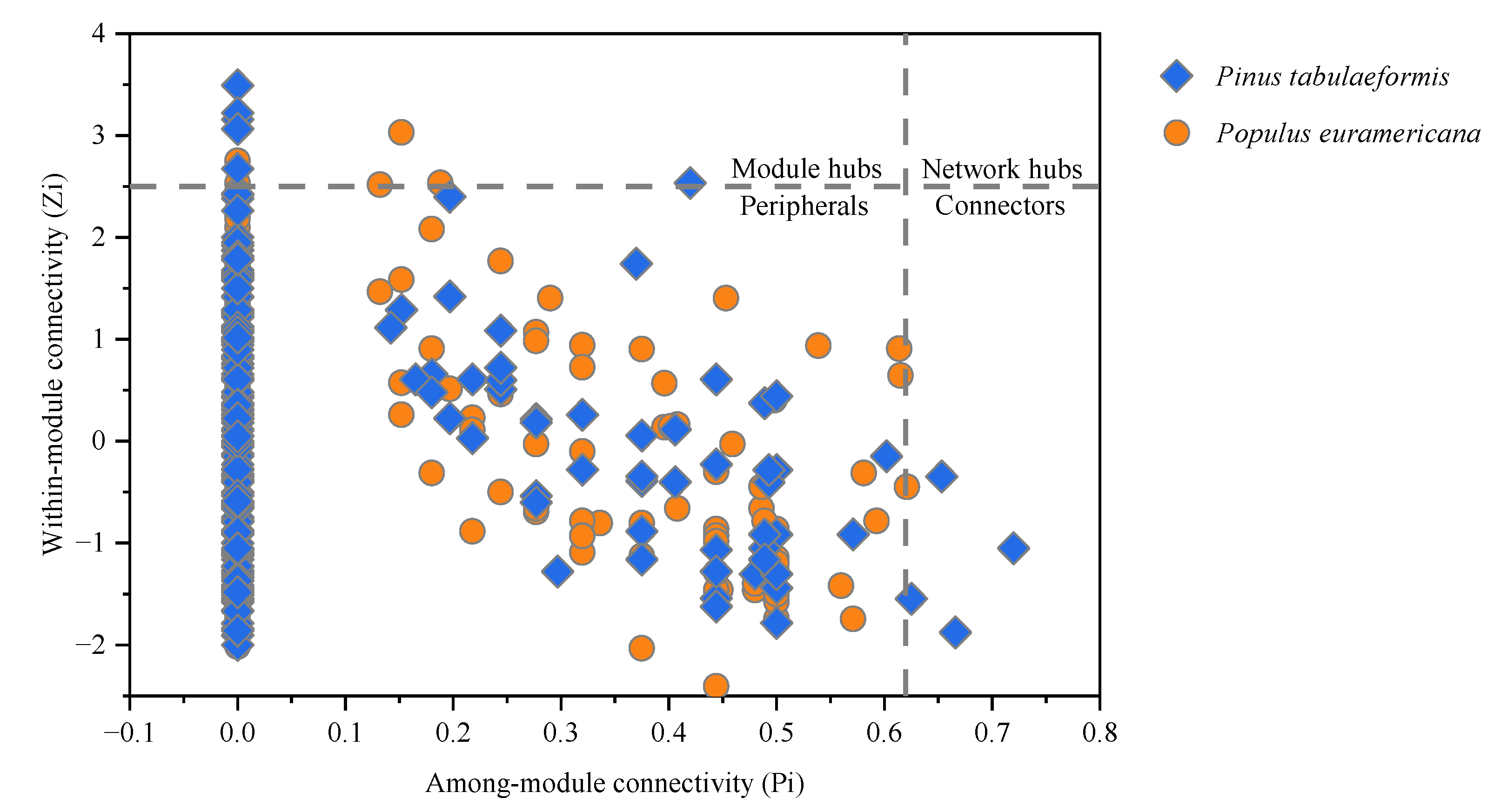
3.3.1. Topological Structural Analysis of the Molecular Ecological Network
3.3.2. Modularization Analysis of the Molecular Ecological Network
3.3.3. Results of the Molecular Ecological Network Construction
4. Discussion
4.1. Differences in Response of Soil Bacterial Communities to Different Vegetation Types
4.2. Differences in Soil Bacterial Community Response to Different Reclamation Years
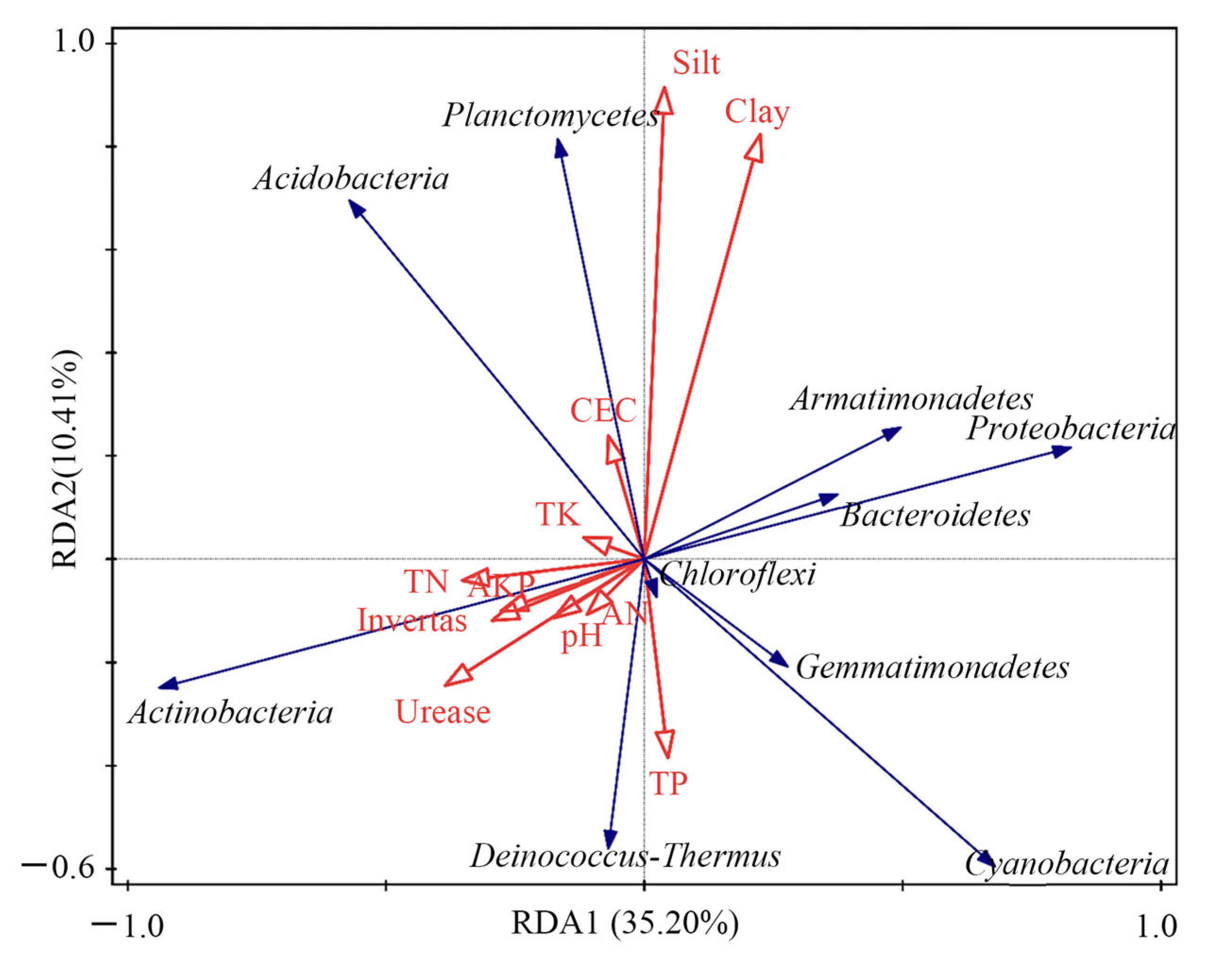
4.3. Driving Mechanisms of Bacterial Community Structure Change in the Reclaimed Soil
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, Z.Q. The 30 years’ land reclamation and ecological restoration in China: Review, rethinking and prospect. Coal Sci. Technol. 2019, 47, 25–35. [Google Scholar] [CrossRef]
- Ministry of Natural Resource of the People’s Republic of China. China Mineral Resources (2020) [EB/OL]. Available online: http://www.mnr.gov.cn/sj/sjfw/kc_19263/zgkczybg/202010/t20201022_2572964.html (accessed on 22 October 2020).
- Du, J.P.; Shao, J.G.; Tan, S.J.; Cao, F. The research of land reclamation in coal mining area: Prospects and progress. J. Chongqing Norm. Univ. (Nat. Sci.) 2018, 35, 131–140. [Google Scholar] [CrossRef]
- Wang, G.; Ren, Y.; Bai, X.; Su, Y.; Han, J. Contributions of beneficial microorganisms in soil remediation and quality improvement of medicinal plants. Plants 2022, 11, 3200. [Google Scholar] [CrossRef]
- Veen, G.F.; Wubs, E.J.; Bardgett, R.D.; Barrios, E.; Bradford, M.A.; Carvalho, S.; De Deyn, G.B.; De Vries, F.T.; Giller, K.E.; Kleijn, D.; et al. Applying the aboveground-belowground interaction concept in agriculture: Spatio-temporal scales matter. Front. Ecol. Evol. 2019, 7, 300. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Sun, H.; Zhang, D.; Zhang, J.; Cai, Z.; Qin, G.; Song, Y. Response of soil microbes to vegetation restoration in coal mining subsidence areas at huaibei coal mine, China. Int. J. Environ. Res. Public Health 2019, 16, 1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y. Microbial Diversity and Enzyme Activity of Soil Reclaimed by Hydraulic Dredge Pump in Coal-Mining Subsidence Areas; China University of Mining and Technology: Xuzhou, China, 2015. [Google Scholar]
- Luo, Z.; Ma, J.; Chen, F.; Li, X.; Zhang, Q.; Yang, Y. Adaptive development of soil bacterial communities to ecological processes caused by mining activities in the Loess Plateau, China. Microorganisms 2020, 8, 477. [Google Scholar] [CrossRef] [Green Version]
- Dimitriu, P.A.; Prescott, C.E.; Quideau, S.A.; Grayston, S.J. Impact of reclamation of surface-mined boreal forest soils on microbial community composition and function. Soil Biol. Biochem. 2010, 42, 2289–2297. [Google Scholar] [CrossRef]
- Pascual, J.A.; Garcia, C.; Hernandez, T.; Moreno, J.L.; Ros, M. Soil microbial activity as a biomarker of degradation and remediation processes. Soil Biol. Biochem. 2000, 32, 1877–1883. [Google Scholar] [CrossRef]
- Harris, J.A. Measurements of the soil microbial community for estimating the success of restoration. Eur. J. Soil Sci. 2003, 54, 801–808. [Google Scholar] [CrossRef]
- Eaton, W.D.; Shokralla, S.; McGee, K.M.; Hajibabaei, M. Sing metagenomics to show the efficacy of forest restoration in the New Jersey Pine Barrens. Genome 2017, 60, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Liu, X.; Tian, Z.; Shao, X. Analysis on characteristics of vegetation and soil bacterial community under 20 Years’ restoration of different tree species: A case study of the qinling mountains. Forests 2021, 12, 562. [Google Scholar] [CrossRef]
- Hou, H.; Wang, C.; Ding, Z.; Zhang, S.; Yang, Y.; Ma, J.; Chen, F.; Li, J. Variation in the soil microbial community of reclaimed land over different reclamation periods. Sustainability 2018, 10, 22867. [Google Scholar] [CrossRef] [Green Version]
- Mummey, D.L.; Stahl, P.D.; Buyer, J.S. Soil microbiological properties 20 years after surface mine reclamation: Spatial analysis of reclaimed and undisturbed sites. Soil Biol. Biochem. 2002, 34, 1717–1725. [Google Scholar] [CrossRef]
- Dangi, S.R.; Stahl, P.D.; Wick, A.F.; Ingram, L.J.; Buyer, J.S. Soil Microbial Community Recovery in Reclaimed Soils on a Surface Coal Mine Site. Soil Sci. Soc. Am. J. 2012, 76, 915–924. [Google Scholar] [CrossRef]
- Strickland, M.S.; Lauber, C.; Fierer, N.; Bradford, M.A. Testing the functional significance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldrop, M.P.; Firestone, M.K. Seasonal dynamics of microbial community composition and function in oak canopy and open grassland soils. Microb. Ecol. 2006, 52, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, W.; Zhao, C.; Shi, R.; Xue, R.; Li, X. Revegetation by sowing reduces soil bacterial and fungal diversity. Ecol. Evol. 2020, 10, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jia, Z.; Sun, Q.; Zhan, J.; Yang, Y.; Wang, D. Ecological restoration alters microbial communities in mine tailings profiles. Sci. Rep. 2016, 6, 25193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Mills, J.G.; Gellie, N.J.; Bissett, A.; Lowe, A.J.; Breed, M.F. High-throughput eDNA monitoring of fungi to track functional recovery in ecological restoration. Biol. Conserv. 2018, 217, 113–120. [Google Scholar] [CrossRef]
- Zhang, L. Effect of Different Combination of Phytoremediation on Soil Ecological Restoration at the Exploited Rare Earth Mine Site; Zhongkai University of Agriculture and Engineering: Guangzhou, China, 2017. [Google Scholar]
- Xu, R.; Zhao, H.; Liu, G.; You, Y.; Ma, L.; Liu, N.; Zhang, Y. Effects of nitrogen and maize plant density on forage yield and nitrogen uptake in an alfalfa-silage maize relay intercropping system in the North China Plain. Field Crops Res. 2021, 263, 108068. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Zhang, F. Abundant and rare bacteria possess different diversity and function in crop monoculture and rotation systems across regional farmland. Soil Biol. Biochem. 2022, 171, 108742. [Google Scholar] [CrossRef]
- Dong, W.H.; Zhang, S.; Rao, X.; Liu, C.A. Newly-reclaimed alfalfa forage land improved soil properties comparison to farmland in wheat-maize cropping systems at the margins of oases. Ecol. Eng. 2016, 94, 57–64. [Google Scholar] [CrossRef]
- Xu, R.; Zhao, H.; Liu, G.; Li, Y.; Li, S.; Zhang, Y.; Liu, N.; Ma, L. Alfalfa and silage maize intercropping provides comparable productivity and profitability with lower environmental impacts than wheat-maize system in the North China plain. Agric. Syst. 2022, 195, 103305. [Google Scholar] [CrossRef]
- Xu, H.; Chen, C.; Pang, Z.; Zhang, G.; Wu, J.; Kan, H. Short-term vegetation restoration enhances the complexity of soil fungal network and decreased the complexity of bacterial network. J. Fungi 2022, 8, 1122. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional molecular ecological networks. MBio 2010, 1, e00169-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Jiang, Y.H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinformaics 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, F.; Yang, L.; Wang, D.; Wang, W. Response of soil bacterial community structure to different reclamation years of abandoned salinized farmland in arid China. Arch. Microbiol. 2019, 201, 1219–1232. [Google Scholar] [CrossRef]
- Du, J.P.; Shao, J.A.; Zhou, C.R.; Sun, J.; Jiang, J.J. Study on reconstruction technology of concave convex landform of temporary construction land in coal mine-taking chongqing mulang coal mine as an example. J. Southwest Univ. (Nat. Sci. Ed.) 2018, 40, 140–148. [Google Scholar] [CrossRef]
- Li, P.F. Study on Soil Quality Restoration and Changes in Microbial Characteristics of Reconstructed Soil in Mining Area of Shanxi-Shaanxi-Inner Mongolia Adjacent Region; Chinese Academy of Science and Ministry of Education (Research Center of Soil and Water Conservation and Ecological Environment): Shangxi, China, 2019. [Google Scholar]
- Wang, J. Impacts of Coal Mining Disturbance on Rhizosphere Microecological and Microbial Reclamation Effect in Western Area of China; China University of Mining and Technology (Beijing): Beijing, China, 2015. [Google Scholar]
- He, L.; Li, Y.Q.; Li, B.C.; Li, J.J. Effects of different vegetation types and reclamation years on soil bacterial community structure in reclaimed mine areas. Environ. Sci. 2017, 38, 752–759. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Bezuidenhout, C.C.; Maboeta, M.S.; Khasa, D.P.; Adeleke, R.A. Structural and functional differentiation of bacterial communities in post-coal mining reclamation soils of South Africa: Bioindicators of soil ecosystem restoration. Sci. Rep. 2020, 10, 17591. [Google Scholar] [CrossRef] [Green Version]
- Buta, M.; Blaga, G.; Paulette, L.; Păcurar, I.; Roșca, S.; Borsai, O.; Grecu, F.; Sînziana, P.E.; Negrușier, C. Soil reclamation of abandoned mine lands by revegetation in northwestern part of transylvania: A 40-year retrospective study. Sustainability 2019, 11, 3393. [Google Scholar] [CrossRef] [Green Version]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.L.; Bouffaud, M.L.; Buée, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngugi, M.R.; Dennis, P.G.; Neldner, V.J.; Doley, D.; Fechner, N.; McElnea, A. Open-cut mining impacts on soil abiotic and bacterial community properties as shown by restoration chrono sequence. Restor. Ecol. 2018, 26, 839–850. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Species | Sample Number | Reclamation Year (a) | Site Name | Species | Sample Number | Reclamation Year (a) |
|---|---|---|---|---|---|---|---|
| North waste dump | Pinus tabulaeformis | N1 | 15 | Inner waste dump | Pinus tabulaeformis | I1 | 9 |
| Populus euramericana | N2 | 15 | Populus euramericana | I2 | 9 | ||
| East waste dump | Pinus tabulaeformis | E1 | 12 | Yinwang waste dump | Pinus tabulaeformis | Y1 | 6 |
| Populus euramericana | E2 | 12 | Populus euramericana | Y2 | 6 | ||
| West waste dump | Pinus tabulaeformis | W1 | 12 | Control area | Pinus tabulaeformis | C1 | |
| Populus euramericana | W2 | 12 | Populus euramericana | C2 |
| Network Index | P. tabulaeformis (0.960) | P. euramericana (0.960) |
|---|---|---|
| Total number of nodes | 642 | 746 |
| Total number of connections | 1518 | 2003 |
| Power law R2 value | 0.701 | 0.712 |
| Average connectivity | 4.729 | 5.370 |
| Average clustering coefficient | 0.541 | 0.579 |
| Average path distance | 9.425 | 11.582 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, H.; Liu, H.; Xiong, J.; Wang, C.; Zhang, S.; Ding, Z. Comparison of Soil Bacterial Communities under Canopies of Pinus tabulaeformis and Populus euramericana in a Reclaimed Waste Dump. Plants 2023, 12, 974. https://doi.org/10.3390/plants12040974
Hou H, Liu H, Xiong J, Wang C, Zhang S, Ding Z. Comparison of Soil Bacterial Communities under Canopies of Pinus tabulaeformis and Populus euramericana in a Reclaimed Waste Dump. Plants. 2023; 12(4):974. https://doi.org/10.3390/plants12040974
Chicago/Turabian StyleHou, Huping, Haiya Liu, Jinting Xiong, Chen Wang, Shaoliang Zhang, and Zhongyi Ding. 2023. "Comparison of Soil Bacterial Communities under Canopies of Pinus tabulaeformis and Populus euramericana in a Reclaimed Waste Dump" Plants 12, no. 4: 974. https://doi.org/10.3390/plants12040974