
Turfgrass Salinity Stress and Tolerance—A Review

Abstract
:1. Introduction
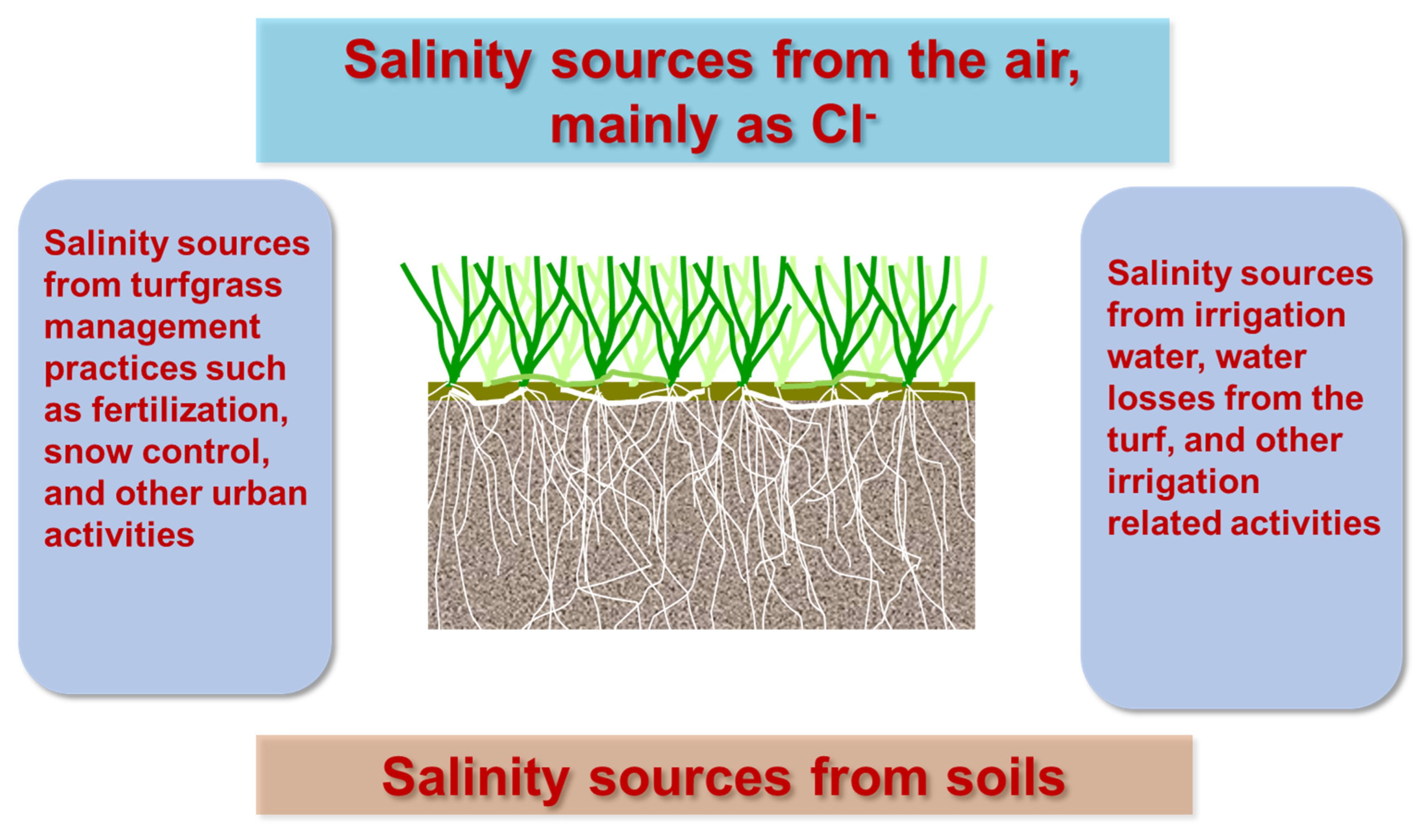
2. Causes and Factors Affecting Soil Salinity for Turf
3. Turfgrass Responses to Saline Stresses
3.1. Sodium (Na+) Toxicity
3.2. Chloride (Cl−) Toxicity
3.3. Potassium (K+) Deficiency
3.4. Calcium (Ca2+) Deficiency
3.5. Boron (B3+) Toxicity
4. Turfgrass Salinity Resistance Mechanisms
4.1. Turfgrass Morphological and Anatomical Resistance Mechanisms
4.2. Sodium and Potassium Uptake Mechanisms
4.3. Ionic Excretions
5. Management Practices to Improve Turfgrass Salinity Resistance
5.1. Turfgrass Species and Cultivar Salinity Tolerance Variations
5.2. Environmental Factors
5.3. Practical Management Approaches

{kind=link}
{kind=link}
{kind=link}
| Exogenous Applications | Turfgrasses Tested | The Rates Used | Salinity Stress Level Range Used | Findings and References |
|---|---|---|---|---|
| 24-epibrassinolide (EBL)-a plant hormone | Perennial ryegrass Tall fescue | 0.15 mg L−1 EBL in 100 mL of the solution per pot (18 cm diameter and 20 cm depth) | 0–6 dS m−1 | The EBL application under salt stress alleviated loss in clipping yield by 35% and 12%; and reduced leaf firing, through weeks 2–6 post-application, by75–40% and 50–20% for perennial ryegrass and tall fescue, respectively [114]. |
| Chitosan (CTS)-a natural polysaccharide | Creeping bentgrass | 0.1, 0.2, 0.5, 1, and 2 g L−1 | 0–200 mM NaCl | The application of CTS increased antioxidant enzyme activities, thereby reducing oxidative damage to roots and leaves [115]. |
| Glycinebetaine (GB)-an osmoprotectant | Creeping bentgrass Kentucky bluegrass Perennial ryegrass Tall fescue | 50, 100, 150, or 200 mM solution of GB of seed priming | 0.1 and 14.6 dS m−1 | Seeds primed with GB showed a higher germination rate (11.0% to 13.9% increase) and seedling growth (19.3% to 20.7% increase) in mannitol or NaCl solution than in distilled water [116]. |
| Melatonin (ME)-a natural hormone | Creeping bentgrass | 0 or 20 μM ME | Drought stress | ME-alleviation of drought-induced leaf senescence in creeping bentgrass was associated with the down-regulation of chlorophyll catabolism and the synergistically interaction with CK-synthesis gene and signaling pathways [117]. |
| Proline-a amino acid | Creeping bentgrass | Proline (10 mM) | Heat stress | Proline-enhanced heat tolerance of creeping bentgrass [118]. |
| Spermidine (Spd)-a polyamine compound | Zoysiagrass (Zoysia japonica Steud) cultivars, ‘Z081′ and ‘Z057′ | Spd 0, 0.15, 0.30, 0.45, 0.60 mM | 0–200 mM NaCl | H2O2 and malondialdehyde (MDA) levels significantly decreased in both cultivars treated with Spd [119]. |
| γ-Aminobutyric acid (GABA)-a neurotransmitter, a chemical messenger in human brain | Creeping bentgrass cv. Penncross | 0.5 mM GABA | 0–250 mM NaCl | GABA application is an efficient approach to enhance salt tolerance of creeping bentgrass during a prolonged period of salt stress and also provides valuable information to better understand key candidate genes and regulatory pathways of GABA-induced salt tolerance in plants [120]. |
5.4. Exogenous Applications of Plant Metabolites to Turfgrass Species
5.5. Symbiotic Relationship with Soil Microorganisms and Genetic Modifications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, G.L.; Kao-Kniffin, J. Applying biodiversity and ecosystem function theory to turfgrass management. Crop. Sci. 2017, 57, S238–S248. [Google Scholar] [CrossRef] [Green Version]
- Ignatieva, M.; Hedblom, M. An alternative urban green carpet. Science 2018, 362, 148–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Pressarakli, M.; Luo, H.; Menchyk, N.; Baldwin, C.W.; Taylor, D.H. Chapter 35: Growth and physiological responses of turfgrasses under stressful conditions. In Handbook of Plant and Crop Physiology Expanded, 4th ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 713–776. [Google Scholar]
- Fan, J.; Zhang, W.; Amombo, E.; Hu, L.; Kjorven, J.O.; Chen, L. Mechanisms of environmental stress tolerance in turfgrass. Agronomy 2020, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Chavarria, M.R.; Wherley, B.; Jessup, R.; Chandra, A. Leaf anatomical responses and chemical composition of warm-season turfgrasses to increasing salinity. Curr. Plant Biol. 2020, 22, 100147. [Google Scholar] [CrossRef]
- FAO-UN Map of Aridity (Global~19 km) 2021. Available online: https://data.apps.fao.org/map/catalog/static/search?keyword=aridity (accessed on 15 January 2023).
- Havlin, J.L.; Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers, 8th ed.; Pearson Education India: Delhi, India, 2016; pp. 51–72. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Zhu, J.-K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Singh, A. Soil salinity: A global threat to sustainable development. Soil Use Manag. 2022, 38, 39–67. [Google Scholar] [CrossRef]
- Carrow, R.N.; Duncan, R.R. Salt-Affected Turfgrass Sites: Assessment and Management; Ann Arbor Press: Chelsea, MI, USA, 1998; ISBN 978-1-57504-091-2. [Google Scholar]
- Marcum, K.B. Use of Saline and non-potable water in the turfgrass industry: Constraints and developments. Agric. Water Manag. 2006, 80, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Bondarenko, S.; Gan, J.; Ernst, F.; Green, R.; Baird, J.; McCullough, M. Leaching of pharmaceuticals and personal care products in turfgrass soils during recycled water irrigation. J. Environ. Qual. 2012, 41, 1268–1274. [Google Scholar] [CrossRef]
- Qian, Y.; Lin, Y. Comparison of soil chemical properties prior to and five to eleven years after recycled water irrigation. J. Environ. Qual. 2019, 48, 1758–1765. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.L.; Mecham, B. Long-term effects of recycled wastewater irrigation on soil chemical properties on golf course fairways. Agron. J. 2005, 97, 717–721. [Google Scholar] [CrossRef]
- Devitt, D.A.; Morris, R.L.; Kopec, D.; Henry, M. Golf course superintendents’ attitudes and perceptions toward using reuse water for irrigation in the southwestern United States. Horttechnology 2004, 14, 577–583. [Google Scholar] [CrossRef]
- Younis, A.; Riaz, A.; Mushtaq, N.; Tahir, Z.; Siddique, M.I. Evaluation of the suitability of sewage and recycled water for irrigation of ornamental plants. Commun. Soil Sci. Plant Anal. 2015, 46, 62–79. [Google Scholar] [CrossRef]
- Carrow, R.N.; Duncan, R.R. Best Management Practices for Saline and Sodic Turfgrass Soils: Assessment and Reclamation; CRC Press: Boca Raton, FL, USA, 2012; pp. 6–10. [Google Scholar]
- McCune, D.C. Effects of airborne saline particles on vegetation in relation to variables of exposure and other factors. Environ. Pollut. 1991, 74, 176–203. [Google Scholar] [CrossRef]
- Alshammary, S.F.; Qian, Y.L.; Wallner, S.J. Growth response of four turfgrass species to salinity. Agric. Water Manag. 2004, 66, 97–111. [Google Scholar] [CrossRef]
- Chang, B.; Wherley, B.G.; Aitkenhead-Peterson, J.A.; West, J.B. Irrigation salinity effects on Tifway bermudagrass growth and nitrogen uptake. Crop Sci. 2019, 59, 2820–2828. [Google Scholar] [CrossRef]
- Karimi, I.Y.M.; Kurup, S.S.; Salem, M.A.M.A.; Cheruth, A.J.; Purayil, F.T.; Subramaniam, S.; Pessarakli, M. Evaluation of bermuda and paspalum grass types for urban landscapes under saline water irrigation. J. Plant Nutr. 2018, 41, 888–902. [Google Scholar] [CrossRef]
- Keyikoglu, R.; Aksu, E.; Arslan, M. Effects of salinity stress on the growth characteristics of four turfgrass species. Fresenius Environ. Bull. 2019, 28, 2942–2948. [Google Scholar]
- Li, J.; Ma, J.; Guo, H.; Zong, J.; Chen, J.; Wang, Y.; Li, D.; Li, L.; Wang, J.; Liu, J. Growth and physiological responses of two phenotypically distinct accessions of centipedegrass (Eremochloa ophiuroides (Munro) Hack.) to salt stress. Plant Physiol. Biochem. 2018, 126, 1–10. [Google Scholar] [CrossRef]
- Tenikecier, H.; Ates, E. Impact of salinity on germination and seedling growth of four cool-season turfgrass species and cultivars. Pol. J. Environ. Stud. 2022, 31, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.L.; Fu, J.M. Response of creeping bentgrass to salinity and mowing management: Carbohydrate availability and ion accumulation. HortSci 2005, 40, 2170–2174. [Google Scholar] [CrossRef] [Green Version]
- Tada, Y.; Komatsubara, S.; Kurusu, T. Growth and physiological adaptation of whole plants and cultured cells from a halophyte turf grass under salt stress. AoB Plants 2014, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Song, C.-P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.-K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack: Ros signaling and stomatal movement. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.; Vishwakarma, K.; Hossen, M.S.; Kumar, V.; Shackira, A.M.; Puthur, J.T.; Abdi, G.; Sarraf, M.; Hasanuzzaman, M. Potassium in plants: Growth regulation, signaling, and environmental stress tolerance. Plant Physiol. Biochem. 2022, 172, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Taïbi, K.; Taïbi, F.; Ait Abderrahim, L.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. South Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant Physiol 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D. Assessing leaf senescence in tall fescue (Festuca arundinacea Schreb.) under salinity stress using leaf spectrum. Eur. J. Hortic. Sci. 2015, 80, 170–176. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef]
- Ozkur, O.; Ozdemir, F.; Bor, M.; Turkan, I. Physiochemical and antioxidant responses of the perennial xerophyte Capparis Ovata Desf. to drought. Environ. Exp. Bot. 2009, 66, 487–492. [Google Scholar] [CrossRef]
- Kurup, S.S.; Abdul Mohsen Ali Salem, M.; Cheruth, A.J.; Sreeramanan, S.; Purayil, F.T.; Al Amouri, A.W.; Pessarakli, M. Changes in antioxidant enzyme activity in turfgrass cultivars under various saline water irrigation levels to suit landscapes under arid regions. Commun. Soil Sci. Plant Anal. 2017, 48, 1989–2001. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; Li, D.; Chen, J.; Zong, J.; Wang, Z.; Chen, X.; Liu, J. Growth response and ion regulation of seashore paspalum accessions to increasing salinity. Environ. Exp. Bot. 2016, 131, 137–145. [Google Scholar] [CrossRef]
- Mastalerczuk, G.; Borawska-Jarmułowicz, B.; Kalaji, H.M. How Kentucky bluegrass tolerate stress caused by sodium chloride used for road de-icing? Environ. Sci. Pollut. Res. 2019, 26, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R. Chlorine Toxicity. Iowa State University Extension TURF Blog 2014. Available online: https://www.extension.iastate.edu/turfgrass/blog/chlorine-toxicity (accessed on 15 January 2023).
- Geilfus, C.-M. Review on the significance of chlorine for crop yield and quality. Plant Sci. 2018, 270, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Heber, U.; Heldt, H.W. The chloroplast envelope: Structure, function, and role in leaf metabolism. Annu. Rev. Plant. Physiol. 1981, 32, 139–168. [Google Scholar] [CrossRef]
- Chen, W.; He, Z.L.; Yang, X.E.; Mishra, S.; Stoffella, P.J. Chlorine nutrition of higher plants: Progress and perspectives. J. Plant Nutr. 2010, 33, 943–952. [Google Scholar] [CrossRef]
- Xu, R.; Fujiyama, H. Comparison of ionic concentration, organic solute accumulation and osmotic adaptation in Kentucky bluegrass and tall fescue under nacl stress. Soil Sci. Plant Nutr. 2013, 59, 168–179. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. IJMS 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Tighe-Neira, R.; Alberdi, M.; Arce-Johnson, P.; Romero, J.; Reyes-Díaz, M.; Rengel, Z.; Inostroza-Blancheteau, C. Role of potassium in governing photosynthetic processes and plant yield. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 191–203. ISBN 978-981-10-9043-1. [Google Scholar]
- Raza, M.A.S.; Saleem, M.F.; Shah, G.M.; Khan, I.H.; Raza, A. Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. J. Soil Sci. Plant Nutr. 2014, 14, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-T.; Lee, C.-L.; Yeh, D.-M. Effects of nitrogen, phosphorus, potassium, calcium, or magnesium deficiency on growth and photosynthesis of Eustoma. Hortscience 2018, 53, 795–798. [Google Scholar] [CrossRef]
- Ye, X.; Wang, H.; Cao, X.; Jin, X.; Cui, F.; Bu, Y.; Liu, H.; Wu, W.; Takano, T.; Liu, S. Transcriptome profiling of Puccinellia tenuiflora during seed germination under a long-term saline-alkali stress. BMC Genom. 2019, 20, 589. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, E.; Nazari, F.; Javadi, T.; Samadi, S.; Teixeira da Silva, J.A. Potassium-enriched clinoptilolite zeolite mitigates the adverse impacts of salinity stress in perennial ryegrass (Lolium perenne L.) by increasing silicon absorption and improving the K/Na ratio. J. Environ. Manag. 2021, 285, 112142. [Google Scholar] [CrossRef] [PubMed]
- Thor, K. Calcium nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Manishankar, P.; Wang, N.; Köster, P.; Alatar, A.A.; Kudla, J. Calcium signaling during salt stress and in the regulation of ion homeostasis. J. Exp. Bot. 2018, 69, 4215–4226. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Bi, A.; Amombo, E.; Li, H.; Zhang, L.; Cheng, C.; Hu, T.; Fu, J. Exogenous calcium enhances the photosystem II photochemistry response in salt stressed tall fescue. Front. Plant Sci. 2017, 8, 2032. [Google Scholar] [CrossRef] [Green Version]
- García-Sánchez, F.; Simón-Grao, S.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Liu, C.; Chatzissavvidis, C.; Pérez-Pérez, J.G.; Cámara-Zapata, J.M. Multiple stresses occurring with boron toxicity and deficiency in plants. J. Hazard. Mater. 2020, 397, 122713. [Google Scholar] [CrossRef]
- Nejad, S.A.G.; Etesami, H. The Importance of boron in plant nutrition. In Metalloids in Plants; Deshmukh, R., Tripathi, D.K., Guerriero, G., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 433–449. ISBN 978-1-119-48721-0. [Google Scholar]
- Bolaños, L.; Lukaszewski, K.; Bonilla, I.; Blevins, D. Why boron? Plant Physiol. Biochem. 2004, 42, 907–912. [Google Scholar] [CrossRef]
- Farooq, M.A.; Saqib, Z.A.; Akhtar, J.; Bakhat, H.F.; Pasala, R.-K.; Dietz, K.-J. Protective role of silicon (si) against combined stress of salinity and boron (b) toxicity by improving antioxidant enzymes activity in rice. Silicon 2019, 11, 2193–2197. [Google Scholar] [CrossRef]
- Carrow, R.N.; Waddington, D.V.; Rieke, P.E. Turfgrass Soil Fertility and Chemical Problems: Assessment and Management; Ann Arbor Press: Chelsea, MI, USA, 2001; ISBN 978-1-57504-153-7. [Google Scholar]
- Jackson, M.B.; Lee, C.W.; Schumacher, M.A.; Duysen, M.E.; Self, J.R.; Smith, R.C. Micronutrient toxicity in buffalograss. J. Plant Nutr. 1995, 18, 1337–1349. [Google Scholar] [CrossRef]
- Soliman, W.S.; Sugiyama, S.; Abbas, A.M. Contribution of avoidance and tolerance strategies towards salinity stress resistance in eight C3 turfgrass species. Hortic. Environ. Biotechnol. 2018, 59, 29–36. [Google Scholar] [CrossRef]
- Chavarria, M.; Wherley, B.; Thomas, J.; Chandra, A.; Raymer, P. Salinity tolerance and recovery at-tributes of warm-season turfgrass cultivars. Hortscience 2019, 54, 1625–1631. [Google Scholar] [CrossRef]
- Wang, J.; An, C.; Guo, H.; Yang, X.; Chen, J.; Zong, J.; Li, J.; Liu, J. Physiological and transcriptomic analyses reveal the mechanisms underlying the salt tolerance of Zoysia japonica Steud. BMC Plant Biol. 2020, 20, 114. [Google Scholar] [CrossRef] [PubMed]
- Pessarakli, M. (Ed.) Handbook of Turfgrass Management and Physiology; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-0-367-38850-8. [Google Scholar]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2020, 225, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcum, K.B. Salinity tolerance mechanisms of grasses in the subfamily Chloridoideae. Crop Sci. 1999, 39, 1153–1160. [Google Scholar] [CrossRef]
- Qian, Y.L.; Engelke, M.C.; Foster, M.J.V. Salinity effects on zoysiagrass cultivars and experimental lines. Crop Sci. 2000, 40, 488–492. [Google Scholar] [CrossRef]
- Naeem, M.; Iqbal, M.; Shakeel, A.; Ul-Allah, S.; Hussain, M.; Rehman, A.; Zafar, Z.U.; Athar, H.-R.; Ashraf, M. Genetic basis of ion exclusion in salinity stressed wheat: Implications in improving crop yield. Plant Growth Regul. 2020, 92, 479–496. [Google Scholar] [CrossRef]
- Marcum, K.B.; Pessarakli, M. Salinity Tolerance and salt gland excretion efficiency of bermudagrass turf cultivars. Crop Sci. 2006, 46, 2571–2574. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Farhangi-Abriz, S. Foliar sprays of salicylic acid and jasmonic acid stimulate H+-ATPase activity of tonoplast, nutrient uptake and salt tolerance of soybean. Ecotoxicol. Environ. Saf. 2018, 166, 18–25. [Google Scholar] [CrossRef]
- Keisham, M.; Mukherjee, S.; Bhatla, S. Mechanisms of sodium transport in plants—Progresses and challenges. IJMS 2018, 19, 647. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Wan, S.; Jiang, Y.; Xia, Y.; Chen, X.; Gao, M.; Cao, Y.; Luo, Y.; Zhou, Y.; Jiang, X. Over-expression of a plasma membrane H+-ATPase SpAHA1 conferred salt tolerance to transgenic Arabidopsis. Protoplasma 2018, 255, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Cuin, T.A.; Bazihizina, N.; Shabala, S. Xylem ion loading and its implications for plant abiotic stress tolerance. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 87, pp. 267–301. ISBN 978-0-12-809390-0. [Google Scholar]
- Zarei, M.; Shabala, S.; Zeng, F.; Chen, X.; Zhang, S.; Azizi, M.; Rahemi, M.; Davarpanah, S.; Yu, M.; Shabala, L. Comparing kinetics of xylem ion loading and its regulation in halophytes and glycophytes. Plant Cell Physiol. 2020, 61, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Cai, X.; Wu, X.; Karahara, I.; Schreiber, L.; Lin, J. Casparian strip development and its potential function in salt tolerance. Plant Signal. Behav. 2011, 6, 1499–1502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yuan, S.; Zhou, M.; Yuan, N.; Li, Z.; Hu, Q.; Bethea, F.G.; Liu, H.; Li, S.; Luo, H. Transgenic creeping bentgrass overexpressing Osa-miR393a exhibits altered plant development and improved multiple stress tolerance. Plant Biotechnol. J. 2019, 17, 233–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium Signaling and Salt Tolerance Are Diversely Entwined in Plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef]
- Tian, F.; Wang, W.; Liang, C.; Wang, X.; Wang, G.; Wang, W. Overaccumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop J. 2017, 5, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Chu, Y.; Chen, H.; Li, X.; Wu, Q.; Jin, L.; Wang, G.; Huang, J. Rice transcription factor Os-MADS25 modulates root Grgwth and confers salinity tolerance via the ABA-mediated regulatory pathway and ROS scavenging. PLoS Genet. 2018, 14, e1007662. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Hu, T.; Zhang, X.; Pang, H.; Fu, J. Exogenous glycine betaine ameliorates the adverse effect of salt stress on perennial ryegrass. J. Amer. Soc. Hortic. Sci. 2012, 137, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Marcum, K.B.; Anderson, S.J.; Engelke, M.C. Salt gland ion secretion: A salinity tolerance mechanism among five zoysiagrass species. Crop Sci. 1998, 38, 806–810. [Google Scholar] [CrossRef]
- Céccoli, G.; Ramos, J.; Pilatti, V.; Dellaferrera, I.; Tivano, J.C.; Taleisnik, E.; Vegetti, A.C. Salt glands in the Poaceae family and their relationship to salinity tolerance. Bot. Rev. 2015, 81, 162–178. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, Y.; Naidoo, G. Cytochemical localization of adenosine triphosphatase activity in salt glands of Sporobolus virginicus (L.) Kunth. South Afr. J. Bot. 1999, 65, 370–373. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, S. Salt gland characteristics and activity of Zoysia matrella Merr. in environments with different levels of salinity. J. Jpn. Soc. Turfgrass Sci. 2020, 48, 142–148. [Google Scholar]
- Imada, S.; Acharya, K.; Yamanaka, N. Short-term and diurnal patterns of salt secretion by Tamarix ramosissima and their relationships with climatic factors. J. Arid. Environ. 2012, 83, 62–68. [Google Scholar] [CrossRef]
- Spiekerman, J.J.; Devos, K.M. The halophyte seashore paspalum uses adaxial leaf papillae for sodium sequestration. Plant Physiol. 2020, 184, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, M.; Bandurska, H.; Breś, W. Response of lawn grasses to salinity stress and protective potassium effect. Agronomy 2021, 11, 843. [Google Scholar] [CrossRef]
- An, P.; Inanaga, S.; Li, X.J.; Eneji, A.E.; Zhu, N.W. Interactive effects of salinity and air humidity on two tomato cultivars differing in salt tolerance. J. Plant Nutr. 2005, 28, 459–473. [Google Scholar] [CrossRef]
- Jiang, K.; Yang, Z.; Sun, J.; Liu, H.; Chen, S.; Zhao, Y.; Xiong, W.; Lu, W.; Wang, Z.-Y.; Wu, X. Evaluation of the Tolerance and Forage Quality of Different Ecotypes of Seashore Paspalum. Front. Plant Sci. 2022, 13, 944894. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.L.; Wilhelm, S.J.; Marcum, K.B. Comparative responses of two Kentucky bluegrass cultivars to salinity stress. Crop Sci. 2001, 41, 1895–1900. [Google Scholar] [CrossRef]
- Challa, A.; Kitila, K.; Workina, M. Evaluation of gypsum and leaching application on salinity reclamation and crop yield at Dugada District, East Shoa Zone of Oromia. Int. J. Environ. Chem. 2022, 6, 1–6. [Google Scholar] [CrossRef]
- Bello, S.K.; Alayafi, A.H.; AL-Solaimani, S.G.; Abo-Elyousr, K.A.M. Mitigating soil salinity stress with gypsum and bio-organic amendments: A review. Agronomy 2021, 11, 1735. [Google Scholar] [CrossRef]
- Rahayu, R.; Mo, Y.G.; Soo, C.J. Amendments on salinity and water retention of sand base rootzone and turfgrass yield. Sains Tanah J. Soil Sci. Agroclimatol. 2019, 16, 103. [Google Scholar] [CrossRef] [Green Version]
- Sekar, S. Efficacy of Salinity Mitigation on Warm Season Turfgrasses. Master’s Thesis, Cornell University, Ithaca, NY, USA, 2016. [Google Scholar]
- Rahman, A.; Mostofa, M.G.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous calcium alleviates cadmium-induced oxidative stress in rice (Oryza sativa L.) seedlings by regulating the antioxidant defense and glyoxalase systems: Calcium-induced cadmium stress tolerance in rice. Braz. J. Bot. 2016, 39, 393–407. [Google Scholar] [CrossRef]
- Zhu, H.; Li, D. Using humus on golf course fairways to alleviate soil salinity problems. Horttechnology 2018, 28, 284–288. [Google Scholar] [CrossRef]
- Fatima, T.; Arora, N.K. Plant Growth-promoting rhizospheric microbes for remediation of saline soils. In Phyto and Rhizo Remediation; Arora, N.K., Kumar, N., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2019; Volume 9, pp. 121–146. ISBN 978-981-329-663-3. [Google Scholar]
- Jha, Y.; Subramanian, R.B. PGPR regulate caspase-like activity, programmed cell death, and antioxidant enzyme activity in paddy under salinity. Physiol. Mol. Biol. Plants 2014, 20, 201–207. [Google Scholar] [CrossRef]
- Kumar, K.; Amaresan, N.; Madhuri, K. Alleviation of the adverse effect of salinity stress by inoculation of plant growth promoting rhizobacteria isolated from hot humid tropical climate. Ecol. Eng. 2017, 102, 361–366. [Google Scholar] [CrossRef]
- Manzoor, M.Z.; Sarwar, G.; Aftab, M.; Tahir, M.A.; Zafar, A. Role of leaching fraction to mitigate adverse effects of saline water on soil properties. J. Agric. Res. 2019, 7, 275–280. [Google Scholar]
- Fu, J.M.; Koski, A.J.; Qian, Y.L. Responses of creeping bentgrass to salinity and mowing management: Growth and turf quality. Hortscience 2005, 40, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Shahba, M.A. Interaction effects of salinity and mowing on performance and physiology of bermudagrass cultivars. Crop Sci. 2010, 50, 2620–2631. [Google Scholar] [CrossRef]
- Shahba, M.A.; Alshammary, S.F.; Abbas, M.S. Effects of salinity on seashore paspalum cultivars at different mowing heights. Crop Sci. 2012, 52, 1358–1370. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Bala, M.; Pareek, A.; Singla-Pareek, S.L.; Singh, A.K. Silicon-mediated abiotic and biotic stress mitigation in plants: Underlying mechanisms and potential for stress resilient agriculture. Plant Physiol. Biochem. 2021, 163, 15–25. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated application of selenium and silicon enhances growth and anatomical structure, antioxidant defense system and yield of wheat grown in salt-stressed soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Xu, X.-B.; Hu, Y.-H.; Han, W.-H.; Yin, J.-L.; Li, H.-L.; Gong, H.-J. Silicon improves salt tolerance by increasing root water uptake in Cucumis sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef]
- Esmaeili, S.; Salehi, H.; Eshghi, S. Silicon ameliorates the adverse effects of salinity on turfgrass growth and development. J. Plant Nutr. 2015, 38, 1885–1901. [Google Scholar] [CrossRef]
- Pakdel, E.; Daoud, W.A.; Seyedin, S.; Wang, J.; Razal, J.M.; Sun, L.; Wang, X. Tunable photocatalytic selectivity of TiO2 /SiO2 nanocomposites: Effect of silica and isolation approach. Colloids Surf. A Physicochem. Eng. Asp. 2018, 552, 130–141. [Google Scholar] [CrossRef]
- Ali, M.; Afzal, S.; Parveen, A.; Kamran, M.; Javed, M.R.; Abbasi, G.H.; Malik, Z.; Riaz, M.; Ahmad, S.; Chattha, M.S.; et al. Silicon mediated improvement in the growth and ion homeostasis by decreasing Na+ uptake in maize (Zea mays L.) cultivars exposed to salinity stress. Plant Physiol. Biochem. 2021, 158, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, W.; Chen, Q.; Ding, R. Effects of silicon on H+-ATPase and H+-PPase activity, fatty acid composition and fluidity of tonoplast vesicles from roots of salt-stressed barley (Hordeum vulgare L.). Environ. Exp. Bot. 2005, 53, 29–37. [Google Scholar] [CrossRef]
- Silva, P.; Gerós, H. Regulation by Salt of Vacuolar H+-ATPase and H+-Pyrophosphatase Activities and Na+/H+ Exchange. Plant Signal. Behav. 2009, 4, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Sever Mutlu, S.; Atesoglu, H.; Selim, C.; Tokgöz, S. Effects of 24-epibrassinolide application on cool-season turfgrass growth and quality under salt stress. Grassl. Sci. 2017, 63, 61–65. [Google Scholar] [CrossRef]
- Geng, W.; Li, Z.; Hassan, M.J.; Peng, Y. Chitosan regulates metabolic balance, polyamine accumulation, and Na+ transport contributing to salt tolerance in creeping bentgrass. BMC Plant Biol. 2020, 20, 506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Rue, K. Glycinebetaine seed priming improved osmotic and salinity tolerance in turfgrasses. Hortscience 2012, 47, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought-induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Rossi, S.; Chapman, C.; Yuan, B.; Huang, B. Improved heat tolerance in creeping bentgrass by γ-aminobutyric acid, proline, and inorganic nitrogen associated with differential regulation of amino acid metabolism. Plant Growth Regul. 2021, 93, 231–242. [Google Scholar] [CrossRef]
- Li, S.; Jin, H.; Zhang, Q. The effect of exogenous spermidine concentration on poly-amine metabolism and salt tolerance in zoysiagrass (Zoysia japonica Steud) subjected to short-term salinity stress. Front. Plant Sci. 2016, 7, 1221. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Cheng, B.Z.; Peng, Y.; Zhang, Y. γ-Aminobutyric acid induces transcriptional changes contributing to salt tolerance in creeping bentgrass. Biol. Plant 2020, 64, 744–752. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Manivannan, P.; Gomathinayagam, M.; Murali, P.V.; Panneerselvam, R. Soil applied propiconazole alleviates the impact of salinity on Catharanthus roseus by improving antioxidant status. Pestic. Biochem. Physiol. 2008, 90, 135–139. [Google Scholar] [CrossRef]
- Nabati, D.A.; Schmidt, R.E.; Parrish, D.J. Alleviation of salinity stress in Kentucky bluegrass by plant growth regulators and iron. Crop Sci. 1994, 34, 198–202. [Google Scholar] [CrossRef]
- Yan, J. Influence of Plant Growth Regulators on Turfgrass Polar Lipid Composition, Tolerance to Drought and Salinity Stresses, and Nutrient Efficiency. Ph.D. Thesis, Virginia Polytechnic and State University, Blacksburg, VA, USA, 1993. [Google Scholar]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed extracts enhance salam turfgrass performance during prolonged irrigation intervals and saline shock. Front. Plant Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [Green Version]
- Drake, A. Effect of Plant Growth Regulators on Creeping Bentgrass Growth and Health during Heat, Salt, and Combined Heat and Salt Stress. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2019. [Google Scholar]
- Khan, A.S.; Ali, S. Preharvest sprays affecting shelf life and storage potential of fruits. In Preharvest Modulation of Postharvest Fruit and Vegetable Quality; Elsevier: Amsterdam, The Netherlands, 2018; pp. 209–255. ISBN 978-0-12-809807-3. [Google Scholar]
- Wei, H.; He, W.; Li, Z.; Ge, L.; Zhang, J.; Liu, T. Salt-tolerant endophytic bacterium Enterobacter ludwigii B30 enhance bermudagrass growth under salt stress by modulating plant physiology and changing rhizosphere and root bacterial community. Front. Plant Sci. 2022, 13, 959427. [Google Scholar] [CrossRef]
- Dashtebani, F.; Hajiboland, R.; Aliasgharzad, N. Characterization of salt-tolerance mechanisms in mycorrhizal (Claroideoglomus etunicatum) halophytic grass, Puccinellia distans. Acta Physiol. Plant 2014, 36, 1713–1726. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant salinity tolerance conferred by arbuscular mycorrhizal fungi and associated mechanisms: A meta-analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef]
- Huang, B.; DaCosta, M.; Jiang, Y. Research advances in mechanisms of turfgrass tolerance to abiotic stresses: From physiology to molecular biology. Crit. Rev. Plant Sci. 2014, 33, 141–189. [Google Scholar] [CrossRef]
- Hu, X.; Hao, J.; Pan, L.; Xu, T.; Ren, L.; Chen, Y.; Tang, M.; Liao, L.; Wang, Z. Genome-Wide Analysis of tandem duplicated genes and their expression under salt stress in seashore paspalum. Front. Plant Sci. 2022, 13, 971999. [Google Scholar] [CrossRef]
- Wang, R.; Wang, X.; Liu, K.; Zhang, X.-J.; Zhang, L.-Y.; Fan, S.-J. Comparative transcriptome analysis of halophyte Zoysia macrostachya in response to salinity stress. Plants 2020, 9, 458. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Hu, Z.; Bi, A.; Fan, J.; Gitau, M.M.; Amombo, E.; Chen, L.; Fu, J. Photosynthesis, antioxidant system and gene expression of bermudagrass in response to low temperature and salt stress. Ecotoxicology 2016, 25, 1445–1457. [Google Scholar] [CrossRef] [PubMed]
- Noor, M.; Fan, J.-B.; Zhang, J.-X.; Zhang, C.-J.; Sun, S.-N.; Gan, L.; Yan, X.-B. Bermudagrass responses and tolerance to salt stress by the physiological, molecular mechanisms and proteomic perspectives of salinity adaptation. Agronomy 2023, 13, 174. [Google Scholar] [CrossRef]
- Huang, X.; Amee, M.; Chen, L. Bermudagrass CdWRKY50 gene negatively regulates plants’ response to salt stress. Environ. Exp. Bot. 2021, 188, 104513. [Google Scholar] [CrossRef]



| Class | EC | ESP | SAR | Soil pH |
|---|---|---|---|---|
| dSm−1 | % | |||
| Saline | >4.0 | <15 | <12 | <8.5 |
| Sodic | <4.0 | >15 | >12 | >8.5 |
| Saline-Sodic | >4.0 | >15 | >12 | <8.5 |
| Intolerant 0–2 dS m−1 0–22 mM NaCl | Slightly Tolerant 2–4 dS m−1 22–44 mM NaCl | Moderately Tolerant 4–8 dS m−1 44–110 mM NaCl | Tolerant 8–30 dS m−1 110–410 mM NaCl | Halophytes >30 dS m−1 >410 mM NaCl |
|---|---|---|---|---|
| Average ocean water salinity level = 35,000 ppm; 44 dS m−1; or equivalent to 600 mM NaCl | ||||
| Distichlis spicata; C4 Inland saltgrass | ||||
| Paspalum vaginatum; C4 Seashore paspalum | ||||
| Puccinellia spp.; C3 Alkaligrasses | ||||
| Sporobolus virginicus; C4 Seashore dropseed | ||||
| Zoysia macrostachya; C4 Macrospike zoysiagrass | ||||
| Zoysia japonica; C4 Japanese lawn grass | ||||
| Zoysia matrella; C4 Manila zoysiagrass | ||||
| Zoysia pauciflora | ||||
| Zoysia sinica; C4 Chinese zoysiagrass | ||||
| Zoysia spp.; C4 Hybrid zoysia grasses | ||||
| Buchloe dactyloides; C4 Buffalograss | ||||
| Cynodon dactylon; C4 Common bermudagrass | ||||
| Cynodon transvaalensis; C4 African bermudagrass | ||||
| Zoysia tenuifolia; C4 Mascarene grass | ||||
| Zoysia spp.; C4 Hybrid zoysiagrasses | ||||
| Agrostis alba; C3 Redtop bentgrass Agrostis canina; C3 Velvet bentgrass Agrostis capillaris; C3 Colonial bentgrass Agrostis spp.; C3 Hybrid bentgrasses Agrostis stolonifera; C3 Creeping bentgrass Axonopus spp.; C4 Carpetgrasses Bouteloua gracilis; C4 Blue grama grass Eremochloa ophiuroides; C4 Centipedegrass Fescue ovina var. ovina; C3 Sheeps fescue Festuca arundinacea; C3 Tall fescue Festuca longifolia; C3 Hard fescue Festuca rubra subsp. commutata; C3 Chewings fescue Festuca rubra var. rubra; C3 Creeping red fescue Koeleria macrantha; C3 prairie Junegrass Lolium perenne; C3 Perennial ryegrass Lolium spp.; C3 Hybrid ryegrasses Pennisetum clandestinum; C4 Kikuyugrass Poa pratensis; C3 Kentucky bluegrass Poa spp.; C3–4 Hybrid bluegrasses Poa supina; C3 Supina bluegrass Poa trivialis; C3 Rough-stock bluegrass Stenotaphrum secundatum; C4 St. Augustinegrass | ||||
| Paspalum notatum; C4 Bahiagrass | ||||
| Lolium multiflorum; C3 Annual ryegrass | ||||
| Poa spp.; C3 Hybrid bluegrasses | ||||
| Poa annua; C3 Annual bluegrass | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Todd, J.L.; Luo, H. Turfgrass Salinity Stress and Tolerance—A Review. Plants 2023, 12, 925. https://doi.org/10.3390/plants12040925
Liu H, Todd JL, Luo H. Turfgrass Salinity Stress and Tolerance—A Review. Plants. 2023; 12(4):925. https://doi.org/10.3390/plants12040925
Chicago/Turabian StyleLiu, Haibo, Jason L. Todd, and Hong Luo. 2023. "Turfgrass Salinity Stress and Tolerance—A Review" Plants 12, no. 4: 925. https://doi.org/10.3390/plants12040925