

Exploring Thrips Preference and Resistance in Flowers, Leaves, and Whole Plants of Ten Capsicum Accessions



Abstract
:1. Introduction
2. Results
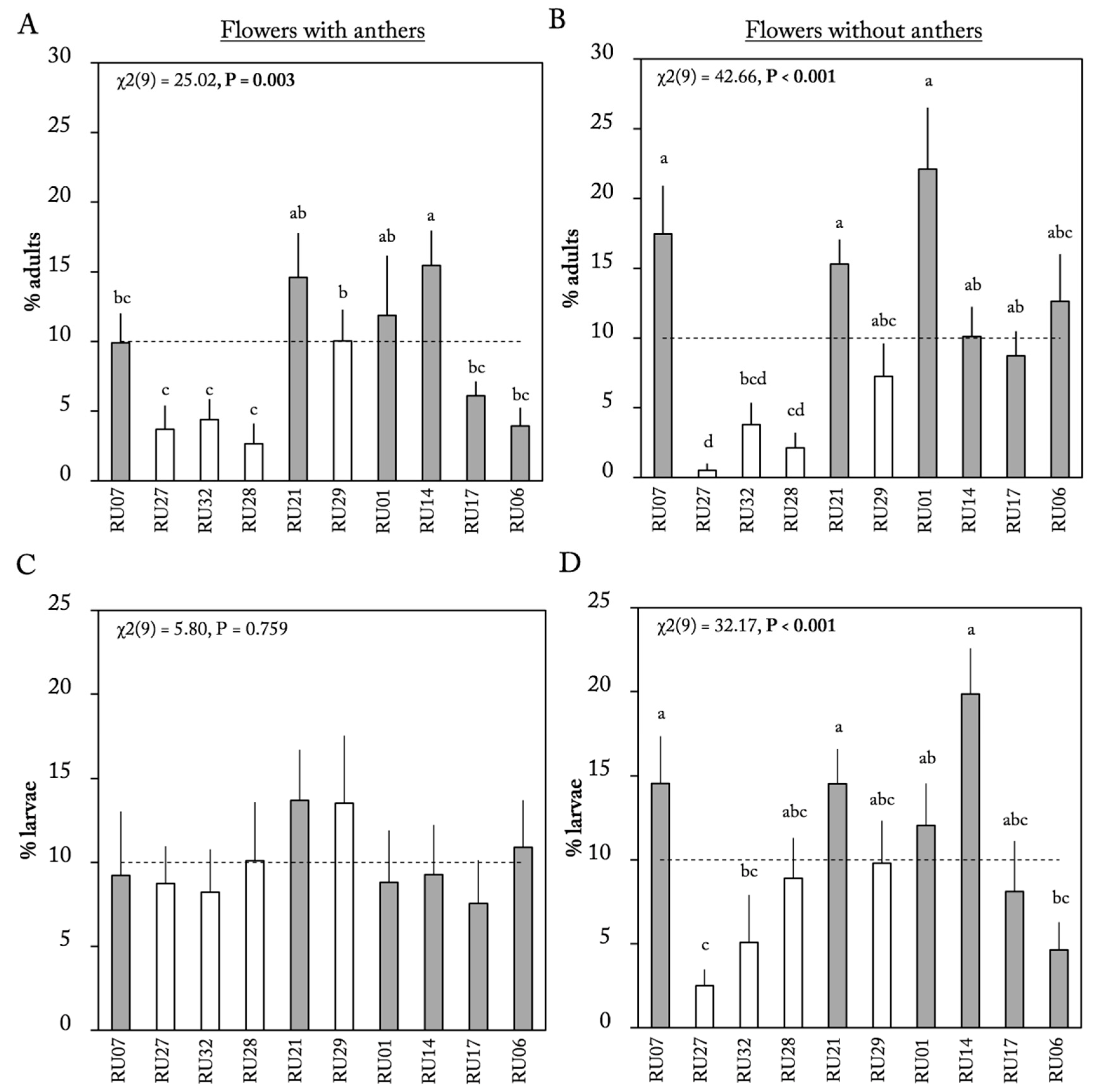
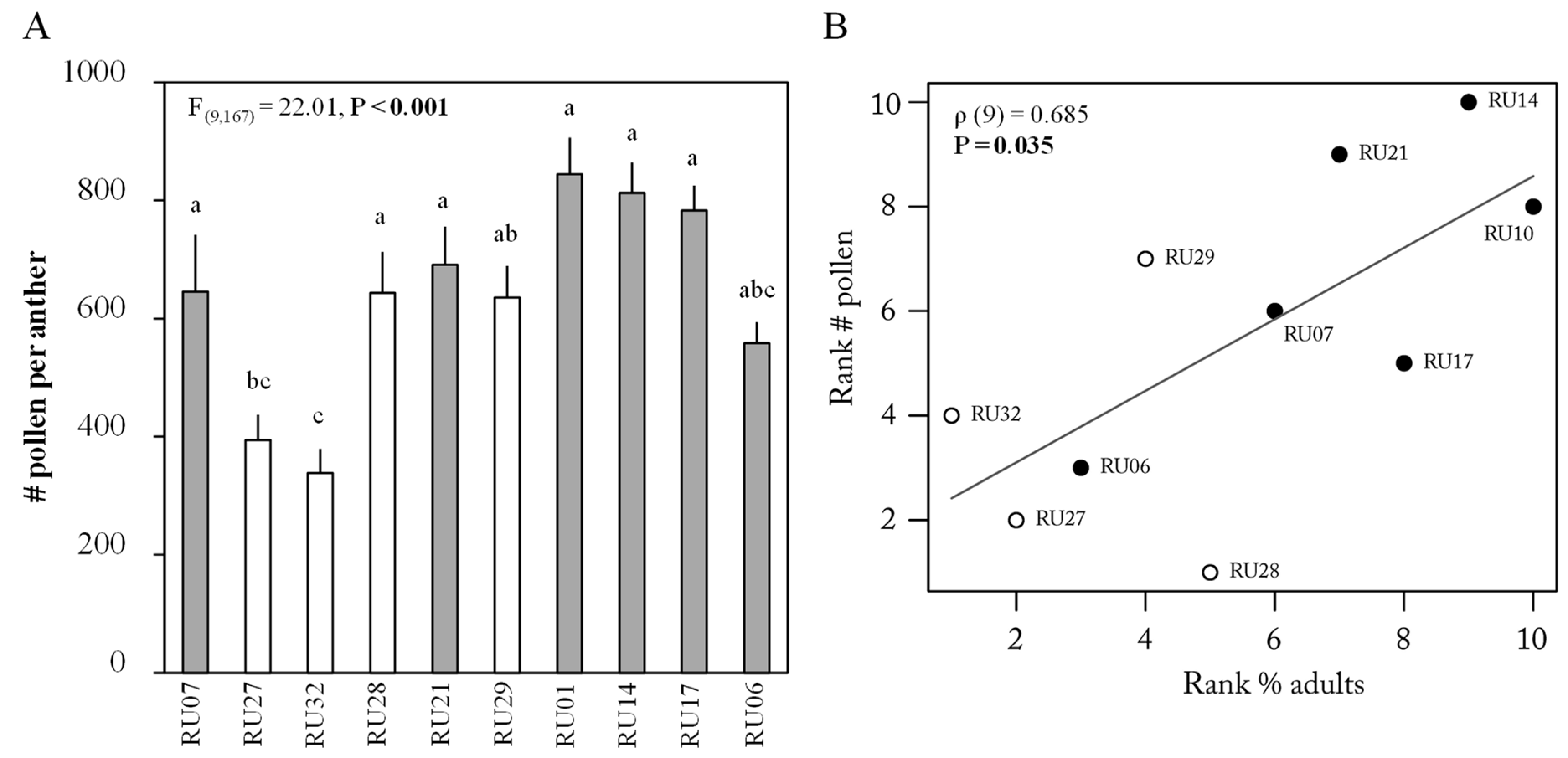
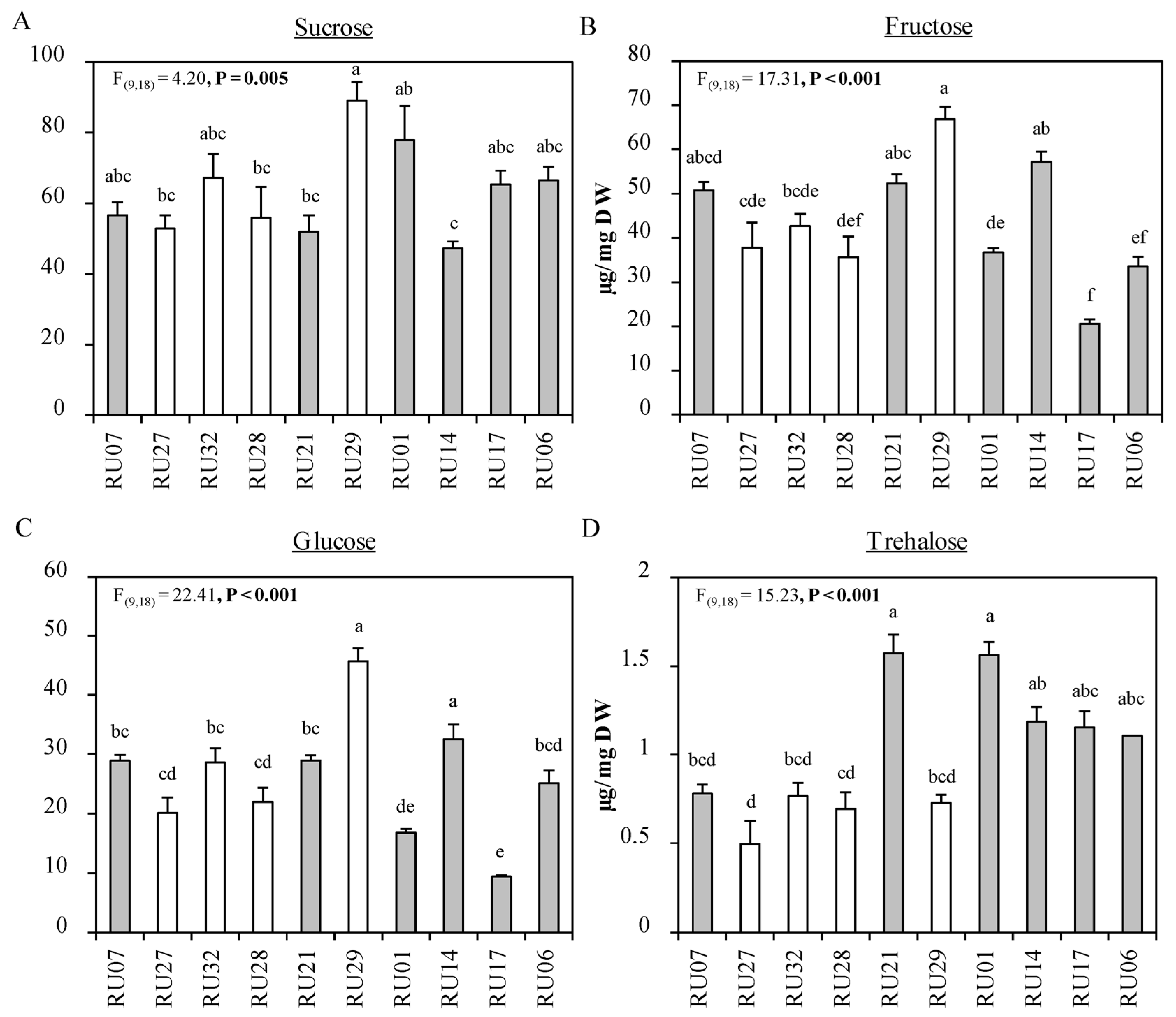
2.1. Flower Choice Assay
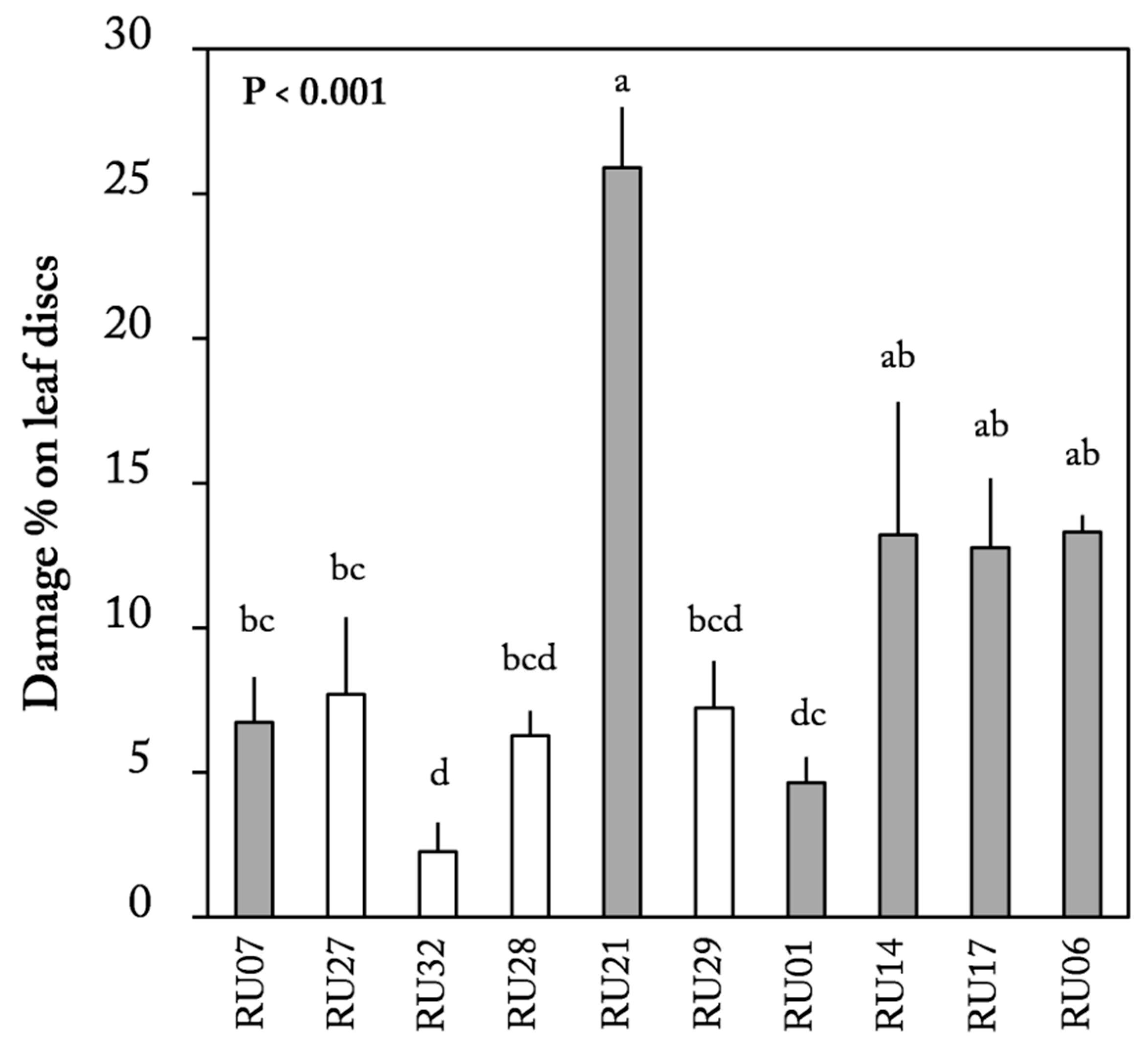
2.2. Leaf Disc Choice Assay
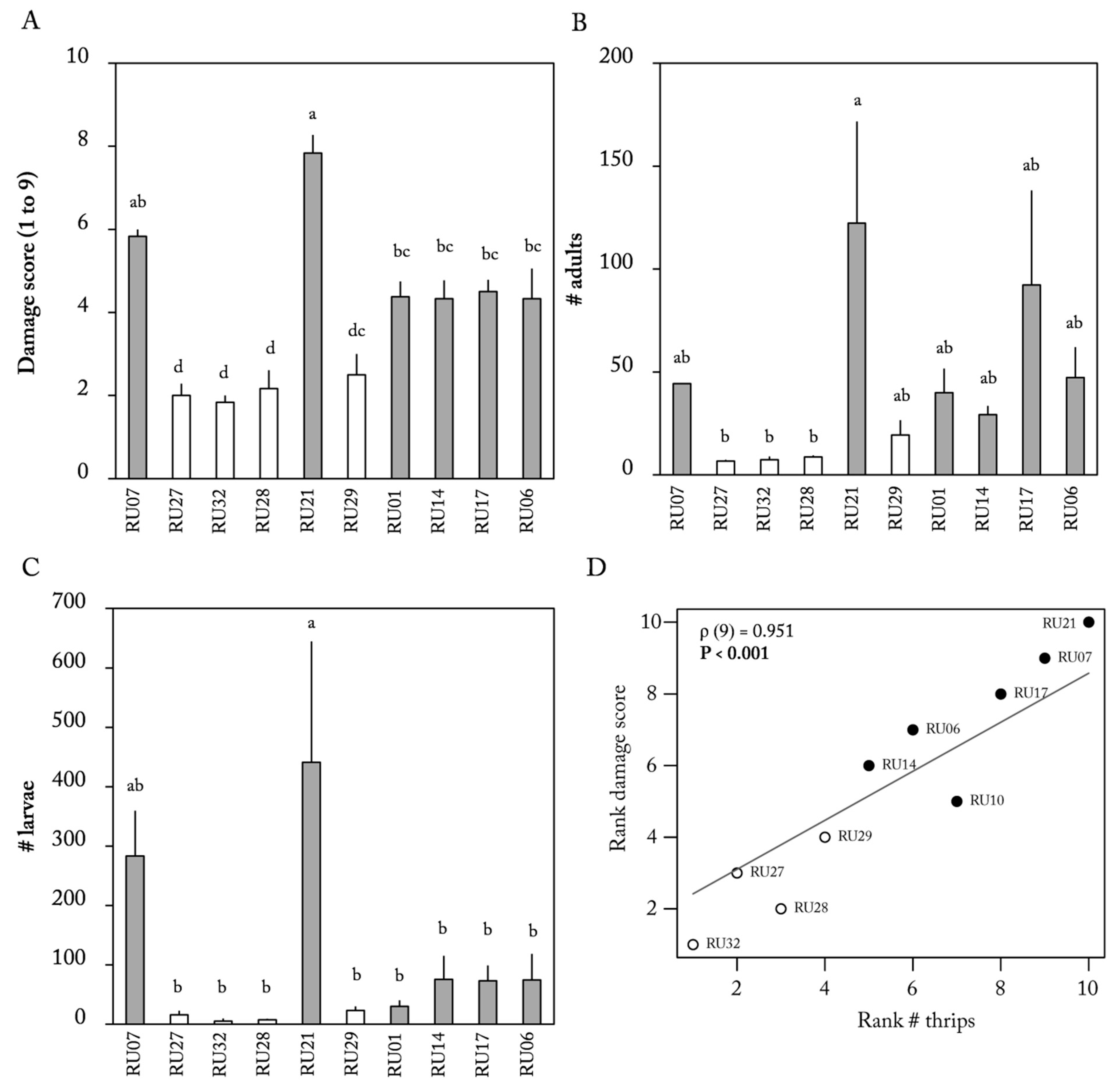
2.3. Thrips Population Development
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Plant Rearing
4.3. Insect Culture
4.4. Full Choice Settling Behavior Assay on Flowers
4.5. Pollen Counting
4.6. Soluble Carbohydrates in Anthers
4.7. Leaf Disc Assay
4.8. Population Development on Whole Plants
5. Statistical Analysis
5.1. Choice Assay Flowers
5.2. Choice Assay Leaf Discs
5.3. Population Development
5.4. Pollen and Anther Data
5.5. Correlation Analyses between Parameters
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenheim, J.A.; Welter, S.C.; Johnson, M.W.; Mau, R.F.L.; Gusukuma-Minuto, L.R. Direct Feeding Damage on Cucumber by Mixed-Species Infestations of Thrips palmi and Frankliniella occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 1990, 83, 1519–1525. [Google Scholar] [CrossRef]
- Tommasini, M.; Maini, S. Frankliniella occidentalis and other thrips harmful to vegetable and ornamental crops in Europe. Wagening. Agric. Univ. Pap. 1995, 95, 1–24. [Google Scholar]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whitfield, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Van Dijken, F.R.; Dik, M.T.A.; Gebala, B.; De Jong, J.; Mollema, C. Western Flower Thrips (Thysanoptera: Thripidae) Effects on Chrysanthemum Cultivars: Plant Growth and Leaf Scarring in Nonflowering Plants. J. Econ. Entomol. 1994, 87, 1312–1317. [Google Scholar] [CrossRef]
- Kogel, W.; Hoek, M.; Mollema, C. Oviposition preference of western flower thrips for cucumber leaves from different positions along the plant stem. Entomol. Exp. Appl. 1997, 82, 283–288. [Google Scholar] [CrossRef]
- Childers, C.C.; Achor, D.S. Feeding and Oviposition Injury to Flowers and Developing Floral Buds of ‘Navel’ Orange by Frankliniella bispinosa (Thysanoptera: Thripidae) in Florida. Ann. Entomol. Soc. Am. 1991, 84, 272–282. [Google Scholar] [CrossRef]
- Cockfield, S.D.; Beers, E.H.; Horton, D.R.; Miliczky, E. Timing of oviposition by western flower thrips (Thysanoptera: Thripidae) in apple fruit. J. Entomol. Soc. Br. Columbia 2007, 104, 10. [Google Scholar]
- Chitturi, A.; Riley, D.G.; Joost, P.H. Effect of pine pollen on settling behavior of Frankliniella occidentalis and Frankliniella fusca (Thysanoptera:Thripidae) on tomato and peanut. Environ. Entomol. 2006, 35, 1396–1403. [Google Scholar] [CrossRef]
- Riley, D.G.; Chitturi, A.; Sparks, A.N. Does natural deposition of pine pollen affect the ovipositional behavior of Frankliniella occidentalis and Frankliniella fusca? Entomol. Exp. Appl. 2007, 124, 133–141. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Pollen-Feeding in Thrips (Insecta, Thysanoptera). J. Zool. 1984, 204, 107–117. [Google Scholar] [CrossRef]
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agric. For. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Wang, Z.H.; Gong, Y.J.; Jin, G.H.; Li, B.Y.; Chen, J.C.; Kang, Z.J.; Zhu, L.; Gao, Y.L.; Reitz, S.; Wei, S.J. Field-evolved resistance to insecticides in the invasive western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) in China. Pest Manag. Sci. 2016, 72, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.G.; Hoddle, M.S. Invasion biology of thrips. Annu. Rev. Entomol. 2006, 51, 67–89. [Google Scholar] [CrossRef]
- Weintraub, P.G. Integrated control of pests in tropical and subtropical sweet pepper production. Pest Manag. Sci. 2007, 63, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Mouden, S.; Sarmiento, K.F.; Klinkhamer, P.G.L.; Leiss, K.A. Integrated pest management in western flower thrips: Past, present and future. Pest Manag. Sci. 2017, 73, 813–822. [Google Scholar] [CrossRef]
- Ssemwogerere, C.; Nyaburu Ochwo-Ssemakula Mildred, K.; Kovach, J.; Kyamanywa, S.; Karungi, J. Species Composition and Occurrence of Thrips on Tomato and Pepper as Influenced by Farmers’ Management Practices in Uganda. J. Plant Prot. Res. 2013, 53, 158–164. [Google Scholar] [CrossRef]
- Fery, R.L.; Schalk, J.M. Resistance in Pepper (Capsicum annuum L.) to Western Flower Thrips [Frankliniella occidentalis (Pergande)]. HortScience 1991, 26, 1073–1074. [Google Scholar] [CrossRef]
- Maris, P.C.; Joosten, N.N.; Peters, D.; Goldbach, R.W. Thrips Resistance in Pepper and Its Consequences for the Acquisition and Inoculation of Tomato spotted wilt virus by the Western Flower Thrips. Phytopathology 2003, 93, 96–101. [Google Scholar] [CrossRef]
- Maharijaya, A.; Vosman, B.; Steenhuis-Broers, G.; Harpenas, A.; Purwito, A.; Visser, R.F.; Voorrips, R.E. Screening of pepper accessions for resistance against two thrips species (Frankliniella occidentalis and Thrips parvispinus). Euphytica 2011, 177, 401–410. [Google Scholar] [CrossRef]
- Visschers, I.G.S.; Peters, J.L.; van de Vondevoort, J.A.H.; Hoogveld, R.H.M.; van Dam, N.M. Thrips resistance is comping of age: Leaf postition and ontogeny are important determinants of damage scores. Front. Plant Sci. 2019, 10, 510. [Google Scholar] [CrossRef]
- Leiss, K.A.; Choi, Y.H.; Abdel-Farid, I.B.; Verpoorte, R.; Klinkhamer, P.G. NMR metabolomics of thrips (Frankliniella occidentalis) resistance in Senecio hybrids. J. Chem. Ecol. 2009, 35, 219–229. [Google Scholar] [CrossRef]
- Koschier, E.H.; Sedy, K.A.; Novak, J. Influence of plant volatiles on feeding damage caused by the onion thrips Thrips tabaci. Crop Prot. 2002, 21, 419–425. [Google Scholar] [CrossRef]
- Escobar-Bravo, R.; Cheng, G.; Kyong Kim, H.; Grosser, K.; van Dam, N.M.; Leiss, K.A.; Klinkhamer, P.G.L. Ultraviolet radiation exposure time and intensity modulate tomato resistance against herbivory through activation of the jasmonic acid signaling. J. Exp. Bot. 2018, 70, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, S.; Funderburk, J.; Stavisky, J.; Olson, S. Population abundance and movement of Frankliniella species and Orius insidiosus in field pepper. Agric. For. Entomol. 2001, 3, 129–137. [Google Scholar] [CrossRef]
- Funderburk, J.; Stavisky, J.; Olson, S. Predation of Frankliniella occidentalis (Thysanoptera: Thripidae) in Field Peppers by Orius insidiosus (Hemiptera: Anthocoridae). Environ. Entomol. 2000, 29, 376–382. [Google Scholar] [CrossRef]
- Hansen, E.A.; Funderburk, J.E.; Reitz, S.R.; Ramachandran, S.; Eger, J.E.; McAuslane, H. Within-Plant Distribution of Frankliniella species (Thysanoptera: Thripidae) and Orius insidiosus (Heteroptera: Anthocoridae) in Field Pepper. Environ. Entomol. 2003, 32, 1035–1044. [Google Scholar] [CrossRef]
- Cao, Y.; Zhi, J.; Zhang, R.; Li, C.; Liu, Y.; Lv, Z.; Gao, Y. Different population performances of Frankliniella occidentalis and Thrips hawaiiensis on flowers of two horticultural plants. J. Pest Sci. 2017, 91, 79–91. [Google Scholar] [CrossRef]
- Ben-Mahmoud, S.; Smeda, J.R.; Chappell, T.M.; Stafford-Banks, C.; Kaplinsky, C.H.; Anderson, T.; Mutschler, M.A.; Kennedy, G.G.; Ullman, D.E. Acylsugar amount and fatty acid profile differentially suppress oviposition by western flower thrips, Frankliniella occidentalis, on tomato and interspecific hybrid flowers. PLoS ONE 2018, 13, e0201583. [Google Scholar] [CrossRef]
- Macel, M.; Visschers, I.G.S.; Peters, J.L.; Kappers, I.F.; De Vos, R.C.; van Dam, N.M. Natural variation in insect resistance of pepper (Capsicum spp.) reveals monomer and dimer acyclic diterpene glycosides as potential chemical defenses. J. Chem. Ecol. 2019, 45, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Visschers, I.G.S.; Peters, J.L.; Timmermans, L.L.H.; Edwards, E.; Ferrater, J.B.; Balatero, C.H.; Stratongjun, M.; Bleeker, P.M.; Herwijnen, Z.; Glawe, G.A.; et al. Resistance to three thrips species in Capsicum spp. depends on site conditions and geographic regions. J. Appl. Entomol. 2019, 143, 929–941. [Google Scholar] [CrossRef]
- Cheng, G.; Huang, H.; Zhou, L.; He, S.; Zhang, Y.; Cheng, X.A. Chemical composition and water permeability of the cuticular wax barrier in rose leaf and petal: A comparative investigation. Plant Physiol. Biochem. 2019, 135, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Wang, L.; Wu, H.; Yu, X.; Zhang, N.; Wan, X.; He, L.; Huang, H. Variation in petal and leaf wax deposition affects cuticular transpiration in cut lily flowers. Front. Plant Sci. 2021, 12, 781987. [Google Scholar] [CrossRef] [PubMed]
- Narayanankutty, A.; Kunnath, K.; Alfarhan, A.; Rajagopal, R.; Ramesh, V. Chemical composition of Cinnamomum verum leaf and flower essential oils and analysis of their antibacterial, insecticidal, and larvicidal properties. Molecules 2021, 26, 6303. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.C.; Nicolson, S.W.; Wright, G.A. Plant secondary metabolites in nectar: Impacts on pollinators and ecological functions. Funct. Ecol. 2017, 31, 65–75. [Google Scholar] [CrossRef]
- Tadmor-Melamed, H.; Markman, S.; Arieli, A.; Distl, M.; Wink, M.; Izhaki, I. Limited ability of Palestine sunbirds Nectarinia osea to cope with pyridine alkaloids in nectar of tree tobacco Nicotiana glauca. Funct. Ecol. 2004, 18, 844–850. [Google Scholar] [CrossRef]
- Kessler, D.; Baldwin, I.T. Making sense of nectar scents: The effects of nectar secondary metabolites on floral visitors of Nicotiana attenuata. Plant J. 2007, 49, 840–854. [Google Scholar] [CrossRef]
- Honda, K.; Ômura, H.; Hayashi, N. Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly, Pieris rapae. J. Chem. Ecol. 1998, 24, 2167–2180. [Google Scholar] [CrossRef]
- Ômura, H.; Honda, K.; Hayashi, N. Floral Scent of Osmanthus fragrans Discourages Foraging Behavior of Cabbage Butterfly, Pieris rapae. J. Chem. Ecol. 2000, 26, 655–666. [Google Scholar] [CrossRef]
- Cao, Y.; Reitz, S.R.; Germinara, G.S.; Wang, C.; Wang, L.; Yang, S.; Gao, Y.; Zhang, W.; Li, C. Host preference of Thrips hawaiiensis for different ornamental plants. J. Pest Sci. 2022, 95, 761–770. [Google Scholar] [CrossRef]
- Mbaabu, P. Evaluation Of Sugar-based Feeding Stimulants’ Effect On Imidacloprid (Confidor® 70wg), Commonly Used To Control Thrips In Chillies (Capsicum annum). Master’s Thesis, University of Nairobi, Nairobi, Kenya, 2014. [Google Scholar]
- Megharaj, K.; Ajjappalavara, P.; Revanappa, R.S.; Tatagar, M.; Satish, D. Study on morphological and biochemical bases for thrips (Scirtothrips dorsalis Hood) resistance in Chilli (Capsicum annum L.). Res. Environ. Life Sci. 2016, 9, 1200–1202. [Google Scholar]
- Brown, S. Interactions of Thrips and Their Control Agents on Host Plants within a Glasshouse Containing a Diverse Collection of Plant Species. Ph.D. Thesis, University of London, London, UK, 2002. [Google Scholar]
- Njau, G.M.; Nyomora, A.M.S.; Dinssa, F.F.; Chang, J.-C.; Malini, P.; Subramanian, S.; Srinivasan, R. Evaluation of onion (Allium cepa) germplasm entries for resistance to onion thrips, Thrips tabaci (Lindeman) in Tanzania. Int. J. Trop. Insect Sci. 2017, 37, 98–113. [Google Scholar] [CrossRef]
- Akhtari, M.; Dashti, F.; Madadi, H.; Rondon, S. Evaluation of resistance to onion thrips (Thrips tabaci Lind.) in several Tareh Irani (Persian leek: Allium ampeloprasum Tareh group) landraces. Arch. Phytopathol. Plant Prot. 2014, 47, 29–41. [Google Scholar] [CrossRef]
- Elbein, A.D.; Carroll, D.; Pastuszak, I.; Pan, Y.T. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Yamada, T.; Yoshida, M.; Nishimura, T. Flies without trehalose. J. Biol. Chem. 2015, 290, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A Key Organic Osmolyte Effectively Involved in Plant Abiotic Stress Tolerance. J. Plant Growth Regul. 2018, 38, 606–618. [Google Scholar] [CrossRef]
- Singh, V.; Louis, J.; Ayre, B.G.; Reese, J.C.; Shah, J. Trehalose phosphate synthase11-dependent trehalose metabolism promotes Arabidopsis thaliana defense against the phloem-feeding insect Myzus persicae. Plant J. 2011, 67, 94–104. [Google Scholar] [CrossRef]
- Singh, V.; Shah, J. Tomato responds to green peach aphid infestation with the activation of trehalose metabolism and starch accumulation. Plant Signal. Behav. 2012, 7, 605–607. [Google Scholar] [CrossRef] [PubMed]
- Mittler, T.E.; Dadd, R.H.; Daniels, S.C. Utilization of different sugars by the aphid Myzus persicae. J. Insect Physiol. 1970, 16, 1873–1890. [Google Scholar] [CrossRef]
- Brødsgaard, H. Coloured sticky traps for Frankliniella occidentalis (Pergande) (Thysanoptera, Thripidae) in glasshouses. J. Appl. Entomol. 1989, 107, 136–140. [Google Scholar] [CrossRef]
- Van Tol, R.W.; Tom, J.; Roher, M.; Schreurs, A.; Van Dooremalen, C. Haze of glue determines preference of western flower thrips (Frankliniella occidentalis) for yellow or blue traps. Sci. Rep. 2021, 11, 6557. [Google Scholar] [CrossRef]
- Elimem, M.; Chermiti, B. Color preference of Frankliniella occidentalis (Pergande)(Thysanoptera: Thripidae) and Orius sp. (Hemiptera: Anthocoridae) populations on two rose varieties. Plant Biotechnol. 2014, 7, 94–98. [Google Scholar]
- Macel, M.; Visschers, I.G.; Peters, J.L.; van Dam, N.M.; de Graaf, R.M. High concentrations of very long chain leaf wax alkanes of thrips susceptible pepper accessions (Capsicum spp). J. Chem. Ecol. 2020, 46, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Maharijaya, A.; Vosman, B.; Pelgrom, K.; Wahyuni, Y.; de Vos, R.C.H.; Voorrips, R.E. Genetic variation in phytochemicals in leaves of pepper (Capsicum) in relation to thrips resistance. Arthropod-Plant Interact. 2019, 13, 1–9. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kiyota, N.; Ikeda, T.; Nohara, T. Acyclic diterpene glycosides, capsianosides C, D, E, F and III, from the fruits of hot red pepper Capsicum annuum L. used in Kimchi and their revised structures. Chem. Pharm. Bull. 2007, 55, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidmann, I.; Di Berardino, M. Impedance flow cytometry as a tool to analyze microspore and pollen quality. In Plant Germline Development: Methods and Protocols; Schmidt, A., Ed.; Springer: New York, NY, USA, 2017; pp. 339–354. [Google Scholar]
- Ribeiro, P.R.; Fernandez, L.G.; de Castro, R.D.; Ligterink, W.; Hilhorst, H.W. Physiological and biochemical responses of Ricinus communis seedlings to different temperatures: A metabolomics approach. BMC Plant Biol. 2014, 14, 223. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Sommer, C.; Straehle, C.; Köthe, U.; Hamprecht, F.A. Ilastik: Interactive learning and segmentation toolkit. In Proceedings of the IEEE International Symposium on Biomedical Imaging: From Nano to Macro, Chicago, IL, USA, 30 March–2 April 2011; pp. 230–233. [Google Scholar]
- Visschers, I.G.S.; van Dam, N.M.; Peters, J.L. Quantification of Thrips Damage Using Ilasik and ImageJ Fjij. Bio-Protoc. 2018, 8, e2806. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Values | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Flower Assays | ||||||||||
| Adults | Offspring | Population Development | ||||||||
| Leaf Disc Assay | Anthers + | Anthers - | Anthers + | Anthers - | Damage Score | # Thrips | ||||
| ρ | Leaf disc assay | 0.427 | 0.973 | 0.178 | 0.811 | 0.166 | 0.039 | |||
| Flower assay | Adults | Anthers + | 0.284 | 0.143 | 0.448 | 0.008 | 0.113 | 0.133 | ||
| Anthers - | −0.018 | 0.503 | 0.657 | 0.178 | 0.104 | 0.178 | ||||
| Offspring | Anthers + | 0.467 | 0.273 | 0.163 | 0.33 | 0.47 | 0.427 | |||
| Anthers - | 0.091 | 0.806 | 0.467 | 0.345 | 0.104 | 0.166 | ||||
| Population development | Score | 0.479 | 0.539 | 0.551 | 0.261 | 0.551 | <0.001 | |||
| # Thrips | 0.673 | 0.515 | 0.467 | 0.284 | 0.479 | 0.951 | ||||
| Species | Accession Code | Ru Code | Accession Name | Geographic Origin |
|---|---|---|---|---|
| C. annuum | CGN16913 | RU07 | Liebesapfel | Germany |
| C. chinense | CGN16994 | RU27 | RU 72–194 | Brazil |
| C. chinense | CGN16995 | RU32 | RU 72–241 | Brazil |
| C. chinense | CGN17004 | RU28 | No.1736; PI 281428 | Suriname |
| C. annuum | CGN17227 | RU21 | Bodroghalmi | Hungary |
| C. chinense | CGN21557 | RU29 | No.4661; PI 159236 | United States of America |
| C. annuum | CGN22151 | RU10 | Ta Pien Chiao; PI 162607 | China |
| C. annuum | CGN23222 | RU14 | Keystone Resistant Giant | United States of America |
| C. annuum | CGN23289 | RU17 | Long Sweet | Zambia |
| C. annuum | CGN23765 | RU06 | CM 331; Criollos de Morelos | Mexico |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visschers, I.G.S.; Macel, M.; Peters, J.L.; Sergeeva, L.; Bruin, J.; van Dam, N.M. Exploring Thrips Preference and Resistance in Flowers, Leaves, and Whole Plants of Ten Capsicum Accessions. Plants 2023, 12, 825. https://doi.org/10.3390/plants12040825
Visschers IGS, Macel M, Peters JL, Sergeeva L, Bruin J, van Dam NM. Exploring Thrips Preference and Resistance in Flowers, Leaves, and Whole Plants of Ten Capsicum Accessions. Plants. 2023; 12(4):825. https://doi.org/10.3390/plants12040825
Chicago/Turabian StyleVisschers, Isabella G. S., Mirka Macel, Janny L. Peters, Lidiya Sergeeva, Jan Bruin, and Nicole M. van Dam. 2023. "Exploring Thrips Preference and Resistance in Flowers, Leaves, and Whole Plants of Ten Capsicum Accessions" Plants 12, no. 4: 825. https://doi.org/10.3390/plants12040825